
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceHealthy and pro-inflammatory gut ecology plays a crucial role in the digestion and tolerance of a novel Gluten Friendly™ bread in celiac subjects: a randomized, double blind, placebo control in vivo study†
Angelo
Andriulli‡
a,
Antonio
Bevilacqua‡
b,
Orazio
Palmieri‡
a,
Anna
Latiano
a,
Rosanna
Fontana
a,
Domenica
Gioffreda
a,
Stefano
Castellana
c,
Tommaso
Mazza
 c,
Anna
Panza
a,
Claudia
Menzaghi
d,
Elvira
Grandone
e,
Lazzaro
di Mauro
f,
Ivana
Decina
g,
Michele
Tricarico
g,
Daniela
Musaico
b,
Markku
Mäki
h,
Jorma
Isola
i,
Alina
Popp
j,
Juha
Taavela
k,
Leonardo
Petruzzi
b,
Milena
Sinigaglia
b,
Maria
Rosaria Corbo
*b and
Carmela
Lamacchia
*bg
c,
Anna
Panza
a,
Claudia
Menzaghi
d,
Elvira
Grandone
e,
Lazzaro
di Mauro
f,
Ivana
Decina
g,
Michele
Tricarico
g,
Daniela
Musaico
b,
Markku
Mäki
h,
Jorma
Isola
i,
Alina
Popp
j,
Juha
Taavela
k,
Leonardo
Petruzzi
b,
Milena
Sinigaglia
b,
Maria
Rosaria Corbo
*b and
Carmela
Lamacchia
*bg
aDivision of Gastroenterology, Fondazione IRCCS-Casa Sollievo della Sofferenza, San Giovanni Rotondo, Foggia, Italy
bUniversità degli Studi di Foggia, Dipartimento di Scienze dell'Agricoltura, degli Alimenti e dell'Ambiente, Foggia, Italy. E-mail: carmela.lamacchia@unifg.it; mariarosaria.corbo@unifg.it
cBioinformatics Unit, Fondazione IRCCS-Casa Sollievo della Sofferenza, 71013 San Giovanni Rotondo (Foggia), Italy
dResearch Unit of Diabetes and Endocrine Diseases, Fondazione IRCCS-Casa Sollievo della Sofferenza, San Giovanni Rotondo, Foggia, Italy
eAtherosclerosis and Thrombosis Unit, Fondazione IRCCS-Casa Sollievo della Sofferenza, San Giovanni Rotondo, Foggia, Italy
fImmunohaematology and Transfusion Medicine Service, Fondazione IRCCS-Casa Sollievo della Sofferenza, San Giovanni Rotondo, Foggia, Italy
gNew Gluten World s.r.l., Foggia, Italy
hFaculty of Medicine and Health Technology, Tampere University and Tampere University Hospital Tampere, Finland
iFaculty of Medicine and Health Technology, Tampere University and Jilab Inc., Tampere, Finland
jUniversity of Medicine and Pharmacy “Carol Davila” and National Institute for Mother and Child Health “Alessandrescu-Rusescu”, Bucharest, Romania
kDepartment of Internal Medicine, Central Finland Central Hospital, Jyväskylä, Finland
First published on 14th January 2022
Abstract
Gluten Friendly™ (GF) is a new gluten achieved through a physicochemical process applied to wheat kernels. The goal of this research was to assess the in vivo effects of Gluten Friendly™ bread on celiac gut mucosa and microbiota. In a double-blind placebo-controlled intervention study, 48 celiac disease (CD) patients were randomized into 3 groups to eat 100 g of bread daily, containing different doses (0; 3 g; 6 g) of GF for 12 weeks. The small-bowel morphology (VH/CrD), intraepithelial densities of CD3+, celiac serology, MUC2, CB1, gut permeability, proinflammatory cytokines, gluten in stools, symptoms, and gut microbial composition were assessed. All 48 CD subjects experienced no symptoms. K-means analysis evidenced celiac subjects clustering around unknown parameters independent of GF dosage: K1 35%; K2 30%; K3 35%. VH/CrD significantly decreased in K1 and K2. VH/CrD did not correlate with IEL increase in K2. 33-mer was not detected in 47% and 73% of patients in both K1 and K2, respectively. VH/CrD and IEL did not change significantly and strongly correlated with the absence of 33-mer in K3. Inflammation and VH/CrD decrease are strongly related with the presence of proinflammatory species at the baseline. A boost in probiotic, butyrate-producing genera, is strongly related with GF tolerance at the end of the trial. Our research suggests that a healthy and proinflammatory ecology could play a crucial role in the digestion and tolerance of the new gluten molecule in celiac subjects. However, GF can be completely digested by gut microbiota of CD subjects and shapes it toward gut homeostasis by boosting healthy butyrate-producing populations. The clinical trial registry number is NCT03137862 (https://clinicaltrials.gov).
Introduction
Celiac disease (CD) is a chronic multiorgan autoimmune disorder that is triggered by dietary gluten proteins in genetically predisposed individuals. Environmental factors such as gut microbiota variations may be implicated in the pathogenesis of CD.1 In fact, gastrointestinal dysbiosis is linked with the inflammatory milieu in celiac patients.2In CD patients, the mucosal layer fails to stabilize the gut microbiota, exposing the host to harmful antigens and pathogens.2 Such dysbiosis is characterized by a reduction of intra- and inter-genera biodiversity,3 showing an imbalance between beneficial bacteria and potentially pathogenic or proinflammatory species, as compared to healthy subjects.4
Gluten Friendly™ (GF) is a new type of gluten5 achieved through a patented, physicochemical process that is applied to wheat kernels before milling. GF has shown, in vitro, some unprecedented and very positive characteristics such as (i) reduced immunoreactivity on gut-derived T-cell lines from celiac patients,6 (ii) reduced the immunogenicity in treated flours by R5 Elisa test and cross-reactivity toward antibodies recognizing the antigenic epitope of gluten proteins in treated wheat kernels,5,7 (iii) positive modulation of celiac gut microbiota with an increase in Lactobacillus spp. and Bifidobacterium spp.8,9 and (iv) increased Transepithelial Electrical Resistance (TEER) in human intestinal goblet cells.10
GF safety and efficacy was tested in vitro on T lines generated by the intestinal mucosa of celiac patients. The cells of each line, highly reactive to gluten, were analyzed for their reactivity to ethanolic extracts obtained from various flours pre- and post-“Gluten Friendly™” treatment. The ethanolic extracts were digested with pepsin and trypsin and subjected to deamidation with tTG. No immunological reactivity, measured as the production of interferon-gamma, to gluten extracted from flour treated according to the GF method to the concentrations of 50 and 100 μg mL−1, was observed.6
GF cytotoxicity assays performed both on mononuclear cells from peripheral blood (PBMCs) from 2 healthy donors and on HT29-Human Colon Epithelial Cells10 showed always high cell viability and no variations in gamma interferon production in the presence of various preparations.
In a double-blind placebo-controlled clinical study, we challenged celiac patients with GF for 12 weeks. The goal of this research was to assess the in vivo effects of Gluten Friendly™ bread on celiac gut mucosa and microbiota. Histological, serological, and metagenomic data were recorded. Symptom charts were collected before, during, and after trial. GF clinical results were compared to previous gluten challenge literature. Finally, we carried out the statistical analysis of celiac gut microbiota composition and variations.
Our research explored the following open questions: (a) What are the effects of GF bread on the serology and histology of celiac patients? (b) Could these effects be mediated by the microbiota? (c) Could there be markers and/or predictors of the GF effect? (d) How does the microbiota ecology at the baseline affect the response of celiac patients to GF? (e) Can GF modulate the microbiota in celiac subjects as suggested by the in vitro studies?
Subjects and methods
Therapeutic intervention study
A prospective, double-blind, placebo-controlled, randomized trial on young and adult subjects with biopsy-proven Celiac Disease (CD) was carried out at the Division of Gastroenterology, Fondazione IRCCS Casa Sollievo della Sofferenza (CSS) Hospital, in San Giovanni Rotondo, Italy. The study protocol was approved by the Ethics Committee of the hospital, and was in compliance with the Good Clinical Practice guidelines of the Helsinki Declaration.Ethical approval was granted by the local Ethics Committee (46/CE). The study was registered on Clinicaltrials.gov (Identifier: NCT03137862).
At the screening period and week 12, participants were asked to consume a multi-sugar drink for a gut permeability test: 5 g lactulose (Duphalac, Solvay Pharmaceuticals Ltd), 1 g Dmannitol ≥98% (Sigma-Aldrich, UK), and 20 g of Sucrose (Sigma-Aldrich, UK) in 200 mL of water.11 The study subjects were then instructed to collect spot urine samples for 5 hours. The subjects also provided a fecal sample for microbiome analysis and gluten quantification in stools.
All the subjects were followed closely and instructed to maintain their usual GFD. From week 0 to week 12, they were called weekly via telephone to verify the study on bread consumption, adherence to GFD, and compliance to the study protocol. Any protocol violation (including ingestion of prescription drugs) was registered. The subjects were also asked to compile a daily food diary as well as provide information on the symptoms (CSI-Celiac Symptom Index Questionnaire; GSRS-Gastrointestinal Symptom Rating Scale), and stool frequency and consistency (Bristol Stool Chart).
The subject safety was monitored through physical examinations and clinical laboratory tests. At the end of the 2nd and 4th weeks, the subjects were tested for anti-TG2. EMA, instead, was tested at the end of the 4th week. Those with positive results underwent repeat endoscopy to check the integrity of the duodenal mucosa. In the event of any histological deterioration, the subjects were considered as therapeutic failures. The study continued for the scheduled 12 weeks for all the subjects with negative serology but also for those with positive serology who decided on a voluntary basis.
At week 12, the subjects returned for a clinical examination, repeat blood drawing for anti-tTG2, anti-EMA, anti-gliadin (AGA both IgG and IgA) antibodies checking, IgG, IgA routine chemistry, complete hematology work up including serum iron, phosphorus, folate, and vitamin D3 measurements. In addition, a follow-up endoscopy was scheduled for all the enrolled subjects to acquire information on the histologic state of the duodenal mucosa.
![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :2:2 using automated randomization system (computer generated) to receive treatment with either a placebo gluten-free bread prepared from corn flour (Group A), or the experimental “Gluten Friendly™” (GF) bread containing either 3 g of GF (Group B) or 6 g of GF (Group C). The overall design of the study is shown in Fig. 1. A positive control with celiac patients fed with 3 and 6 g of gluten was not performed because it was not considered to be ethical. All the study participants, care providers, data managers, and personnel remained blinded of the group assignment throughout the study.
:2:2 using automated randomization system (computer generated) to receive treatment with either a placebo gluten-free bread prepared from corn flour (Group A), or the experimental “Gluten Friendly™” (GF) bread containing either 3 g of GF (Group B) or 6 g of GF (Group C). The overall design of the study is shown in Fig. 1. A positive control with celiac patients fed with 3 and 6 g of gluten was not performed because it was not considered to be ethical. All the study participants, care providers, data managers, and personnel remained blinded of the group assignment throughout the study.
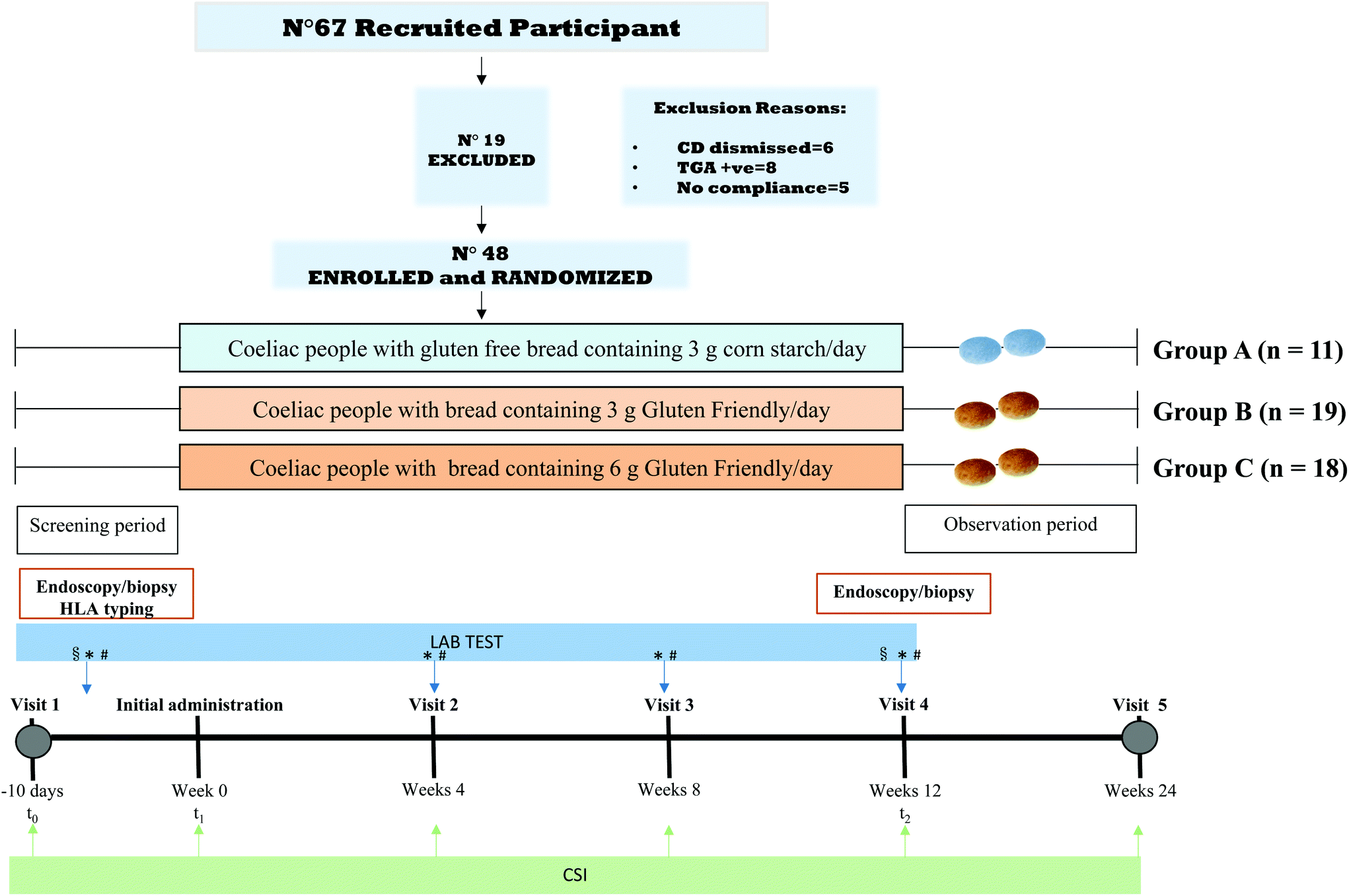 | ||
| Fig. 1 Participant flow-chart and therapeutic intervention study design. Celiac subjects were recruited and randomized into three groups receiving bread buns containing either 3g of GF, 6 g of GF, or a placebo gluten free bread containing 3 g of cornstarch, daily for 12 weeks. Upper gastrointestinal endoscopy with duodenal mucosal biopsies was performed on day −10 (baseline) and post treatment on 12 weeks. The biopsy specimens were explored for the measures of the mucosal morphological change as villous height crypt depth ratio (VH/CrD) and for the intraepithelial lymphocytes (IELs). In addition, the expression of Mucin 2 (MUC2) and Cyclin B1 (CB1) and the tissue EMA were also evaluated. Human leukocyte antigen (HLA) genotyping was performed in the screening period. Lab tests, namely anti-endomysial antibodies (EMA), anti-transglutaminase antibodies (TGA), anti-gliadin (AGA both IgG and IgA) and clinical/hematology exams were performed at the screening period and at week 4, 8, and 12 (*). Permeability test, cytokines panel, microbiome analysis and immunotoxic gluten peptides (GIP) were performed at screening period and week 12 (§). Patients were tested weekly through a Biocard Celiac Test (#). The symptoms were registered before the trial, at week 4, 8, and 12 of GF administration, as well as at the end of the trial (observation period) by Celiac Symptoms Index Questionnaire (CSI). | ||
Study treatment
The recipe was as follows.
1. Group B: 50 g of GF flour, 50 g of corn/rice starch, 2.5% yeast, 2% salt, 65% water. The actual content of GF was 3 g per day (1.5 g in each bun; see the protocol reported in the following lines).
2. Group C: 100 g of GF flour, 2.5% yeast, 2% salt, 65% water; the actual content of GF was 6 g per day (3.0 g in each bun; see the protocol reported in the following lines).
3. The control bread (group A) was prepared with the following ingredients: 100 g of corn/rice starch, 2.5% yeast, 2% salt, 65% water. The actual content of GF was 0 g (gluten free bread).
Doughs were prepared by mixing all the ingredients in a dough mixer, then leavening for approximately 20 min (at 28/30 °C and 70% humidity) and finally shaped into buns of 50 g each. The leavened bread buns were placed on Teflon perforated trays and baked in an oven at 220 °C for 20 min. The bread buns (2 buns of 50 g per day) were baked daily, packed in a controlled atmosphere (N2/CO2, 30/70%), and home delivered once a week to all the study participants.
Analyses
Histology. Routinely formalin-fixed samples were processed for paraffin blocks using a standard paraffin-infiltration protocol. Each biopsy was embedded in a separate paraffin block under a dissection microscope, aiming at a perpendicular cutting plane to the mucosal lumen surface.13 A crucial step in the procedure involves training laboratory technicians to obtain correctly oriented cuttings of biopsy specimens for morphometric evaluation. Six tissue sections (thickness 3–4 μm) were cut on the SuperFrost Plus microscope objective slides from each block by tilting the block 5–10 degrees between each cut. For VH/CrD measurements, the slides were stained with hematoxylin and eosin.
Immunohistochemistry. Immunohistochemistry of IELs was done with anti-CD3 antibody (Clone SP7, REF: RM-9107-S1, Thermo Fisher Scientific, Waltham, MA. Diluted 1
:300). Crypt cell proliferation index was determined with anti-Cyclin B1 Ab-4 antibody (Clone GNS11, REF: MS-869-P1, NeoMarkers Inc., Fremont, CA. Diluted 1:100). The expression of MUC2 in the villus epithelium goblet cells was visualized with anti-MUC2 antibody (Clone BSB-45, REF: BSB 6160, Bio SB Inc., Santa Barbara, CA. Diluted 1:250). A standard IHC protocol using high-pH antigen retrieval (Tris-EDTA buffer, pH 9) and a peroxidase-polymer based detection kit (Histofine High Stain HRP (MULTI), REF: 414483F, Nichirei Biosciences Inc.) was employed. Diaminobenzidine (DAB) was used as the chromogen and hematoxylin as the counterstain. Stainings were carried out using an automated stainer platform (LabVision Autostainer).
The slides were scanned as high-resolution whole-slide images at a resolution of 0.16 μm per pixel (Jilab Inc., Tampere, Finland). Areas containing the strongest labeling at the lowest magnification were chosen for digital image analysis (hotspot sampling). Three to six villi covering at least 300 enterocytes were counted for IELs. Counting was done automatically using the Celiac Slide Analyzer (Jilab Inc., Tampere, Finland), which is a modification of a multi-purpose IHC cell counting software.14 The results were reported as the number of IELs and MUC2+ cells per 100 enterocytes, or as the percentage of proliferating Cyclin B1+ crypt cells. The measurements were independent of the clinical information.
Digital histomorphometry. All slides were scanned as whole slide images using a SlideStrider scanner at a resolution of 0.28 μm per pixel (Jilab Inc., Tampere, Finland). Images were stored as JPEG2000 files in the image server and viewed via the internet with a web-based client software developed for this study (Celiac Slide Viewer). Villus height and crypt depth were measured digitally by drawing polylines. Two academic observers (JT, AP) analyzed all the slides independently and an average of their measurements was used as the final result for the VH/CrD ratio. During the GF challenge, a decrease in the VH/CrD of 0.5 or more was considered to be significant and equivalent to the clinical gluten sensitivity.13,15 CD3 positive intraepithelial T-lymphocytes were enumerated with the AutoIEL software, and the results were expressed per 100 enterocytes. After the GF challenge, an increase in the IEL counts of >30% was considered to be significant and equivalent to the clinical gluten sensitivity.15 Proliferating Cyclin B1 staining-positive crypt epithelial cells were enumerated with ImmunoRatio 2.5 software.16 For both IELs and CB1, at least 300 cells were counted.
Cytokines. Plasma samples drawn at the baseline, during, and at the end of the study period were immediately stored at −80 °C at the CSS Hospital. The premixed multiplex beads of the Bio-Plex human cytokine Human 27-Plex Panel (Bio-Rad Laboratories, Milan, Italy), which included twenty-seven cytokines and chemokines [IL-1b; IL-1ra, IL-2, IL-4, Il-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-10, IL-12 (P70), IL-13, IL-15, IL-17, Basic FGF, Eotaxin, G-CSF, GM-CSF, IFN-γ, IP-10, MCP-1 (MCAF), MIP-1a, MIP-1b, PDGF-BB, RANTES, TNF-α, VEGF] were used in the presence of 30 μL of plasma. All the plasma samples and standards were run in duplicate, and fluorescent signals were read using Biorad 200 system (Bio-Rad).
Sample collection. Subjects enrolled in the study were provided with guidelines for dietary restrictions. On the morning of the test, subjects drank a solution containing 5 g of lactulose, 1 g of mannitol, and 20 g of sucrose in 200 mL of deionized water. The study subjects were then instructed to collect spot urine samples for 5 hours in a collecting bottle with 1 mL chlorhexidine (1 mg mL−1) as the antimicrobial agent.
Total urine volume was measured, and 10 mL aliquots were stored at −20 °C until the analysis. The urine samples were allowed to thaw at room temperature, then stirred for 1 minute using a vortex mixer, and then centrifuged at 5000g for 4 min to remove the sediment.
For each study participant, a 50 μL urine sample was mixed with an internal standard solution of 450 μL. In parallel, 50 μL of the control and 50 μL of the standard were each mixed with 450 μL of the internal standard solution. After mixing, a 200 μL aliquot from each of the three samples was transferred into a glass vial and injected into HPLC-MS/MS.11
Instrumentation. The measurements were performed on a API 3000 Tandem Mass Spectrometer (AB Sciex, Toronto, Canada) equipped with a turbo ion spray source. Quantification was achieved using multiple reaction monitoring (MRM).
Chromatographic conditions. High-Performance Liquid Chromatography (HPLC) separation was performed using a 150 × 2 mm, Luna 5 μm NH2 100 Å column (Phenomenex, USA) operating at a flow rate of 300 μL min−1, and eluted with a 4 min linear gradient from 70% to 30% acetonitrile in water. The oven temperature was set at 40 °C. The injection volume was 5 μL, and the total analysis time was 9 min.
Mass spectrometer conditions. The electrospray ionization (ESI) source was operated in the negative mode. The capillary voltage was set to 3500 V at a temperature of 300 °C. The gas source was set as follows: nebulizer gas 8 (arbitrary units), curtain gas 7 (arbitrary units), and collision gas 4 (arbitrary units).
Each multiple reaction monitoring (MRM) transition was collected at a resolution of 0.7 amu full width half maximum (FWHM) in the first quadrupole, with a scan time of 0.1 s. The tube lens and collision settings were established individually for each compound. The conditions for the detection of lactulose, mannitol, and raffinose were obtained by the direct infusion of a standard solution (10 μg mL−1), in line with the HPLC at the initial mobile phase conditions.
Method validation. In order to validate the method, the linearity, LOQ, imprecision, accuracy, recovery, and matrix effect were assessed.17 The within-run precision and accuracy ranged from 0.9% to 3.2% and from 98.0% to 102.0%, respectively. The between-run precision and accuracy ranged from 2.2% to 4.7% and from 96.2% to 101.3%, respectively.
Subsequently, the amplicons were purified using AMPure XP beads (Beckman Coulter, Milan, Italy). Afterward, the ligation of the dual indexing adapters was performed in the presence of Nextera XT Index Primer 1 and Primer 2 (Illumina), Taq Phusion High-Fidelity (Thermo Fisher Scientific), and 5 μL purified DNA, according to the manufacturer's instructions. The products were purified using AMPure XP beads to create the final cDNA library.
Libraries concentration and fragment size were measured using a fluorometric based system (Qubit dsDNA BR Assay System; Thermo Fisher Scientific) and an Agilent 2200 TapeStation Bioanalyzer (HS D1000, Agilent Technologies, Santa Clara, CA, USA), respectively. Equal amounts of cDNA libraries were pooled, denatured with NaOH, diluted with a hybridization buffer to 7 pM following the Illumina protocol, and spiked with 20% PhiX (Illumina). The libraries were loaded into a flow cell V2 (500 cycles) by paired-end sequencing (2 × 250) (Illumina), and sequenced with MiSeq (Illumina) according to the manufacturer's recommendations.
Statistical analysis
The subjects’ sample size was pragmatic, and was calculated to assess the safety and tolerability of the GF bread while minimizing unnecessary participant exposure.All the serum laboratory tests, namely, anti-tTG2, anti-EMA, anti-AGA antibodies, serum iron, phosphorus, folate, vitamin D3, the cytokine panel, the concentration of GIP (33-mer) in stools, the EMA in the biopsies, and the lactulose/mannitol ratio in urine were evaluated by comparing the studied time points (Fig. 1). All the statistical tests were performed through Statistica for Windows, ver. 12.0 software (Statsoft, Tulsa, OK, USA).
The values of VH/CrD, cytokines (2, 4, 6, 8, IFN, TNF), IELs, MUC2, CB1, 33-mer detection, and antibodies were analyzed by means of one-way analysis of variance (ANOVA) and Tukey's test. If data distribution did not show a normal trend, the values were analyzed through the non-parametric Friedman test. The critical level of P was set to 0.05. Here, we shown only statistically significant results or those with a clear involvement in the CD pathogenesis (to note, although all the panel was analyzed, only the cytokines named 2, 4, 6, 8, IFN, and TNF were shown).
where E is the value at the end of the trial, and B is the value at the beginning; 33-mer detection was reported as the percentage of subjects with a negative G12 (i.e., subjects where 33-mer was not detected).

For K-means clustering, the parameter K (number of cluster) was set to 3, while the interactions were at least 10. The initial inter-cluster centers were evaluated by sorting the distance and taking observations at constant intervals.
Preparation was done using DADA225 and included denoising, chimera checking, pair-joining, and clustering through dereplication. Taxonomic classification, from the domain to the genus, was obtained through the embedded Naïve Bayes fitted classifier, trained on the Silva release.132 rRNA database.26 All the samples were then rarefied based on the outcomes of the feature table produced during the preparation phase. Any possible loss of information was ruled out by the examination of the rarefaction plots.
Alpha and Beta diversity analyses were performed in order to assess both within-sample and between-sample diversity, using several metrics. In particular, Faith's Phylogenetic Diversity index was investigated for Alpha diversity, and was tested using the Kruskal–Wallis test. Unweighted UniFrac distance was used to build a PCoA plot for Beta diversity, and was tested using the PERMANOVA test.27 Finally, the ANCOM abundance test28 was applied at the genus level in order to identify the features that were differentially abundant across the sample groups.
The readings of each genus, family, or species were modelled as the recovery detection (%) on the total number of reading for each subject; then, the changes in the recovery detection were evaluated as the difference between the recovery after trial and the recovery before trial. These values were analyzed through the Friedman test (P < 0.05) to highlight the significant differences (P = <0.05).
Results and discussion
Between April and September 2017, 67 celiac subjects were recruited and forty-eight were enrolled in a 12-week therapeutic intervention study (Fig. 1) after the informed consent form was signed and approved from the local Ethics Committee (N.46/CE). 19 subjects were excluded for these reasons: CD dismissed (n = 6), presence of TGA (n = 8), and no compliance (n = 5). The subjects ranged from 15 to 75 years of age, with a mean age of 38 years (Table 1). Patients randomized into 3 groups ate 100 g of bread daily, containing different doses of GF. Group A (11 subjects) received 0 g; Group B (19 subjects) received 3 g; and group C (18 subjects) received 6 g (Fig. 1). All the patients completed the study. The strict monitoring of a gluten-free diet (GFD) was imposed throughout the trial.| Gluten friendly | |||
|---|---|---|---|
| Placebo | 3.0 g | 6.0 g | |
| N of patients | 11 | 19 | 18 |
| Female | 7 | 11 | 10 |
| Age in years, mean (ds) | 38.6 ± 21.1 | 35.0 ± 15.3 | 39.1 ± 15.1 |
| <20 | 3 | 3 | 3 |
| 20–49 | 5 | 11 | 10 |
| ≥50 | 3 | 5 | 5 |
| Measurement | |||
| Height, means (SD) in cm | 168.5 ± 9.4 | 168.9 ± 9.6 | 168.4 ± 10.1 |
| BMI mean (SD) | 21.7 ± 2.4 | 22.8 ± 3.8 | 23.5 ± 3.1 |
| <20 | 2 | 4 | 2 |
| 20–24.9 | 9 | 11 | 9 |
| 25–29.9 | 3 | 6 | |
| ≥30 | 1 | 1 | |
| Duration of GFD in years | |||
| Mean (SD) | 8.6 ± 6.2 | 12.3 ± 6.6 | 11.6 ± 9.1 |
| 1–1.9 | 0 | 0 | |
| 2–3.9 | 2 | 1 | 3 |
| 4–5.9 | 2 | 4 | 4 |
| 6–7.9 | 2 | 2 | 1 |
| >8 | 5 | 12 | 10 |
| HLA | |||
| DQ2 | 9 (82%) | 16 (84%) | 17 (94%) |
| DQ8 | 1 (9%) | 0 (0%) | 1 (6%) |
| DQ2–DQ8 | 1 (9%) | 3 (16%) | 0 (0%) |
Gluten Friendly™ does not induce symptoms in celiac subjects
Gastrointestinal symptoms from mild to severe can occur frequently in patients with untreated celiac disease. A life-long GFD can significantly improve gastrointestinal conditions but symptoms may not completely disappear.Gluten challenge studies have shown that celiac patients experience a rapid, dose-dependent surge of symptoms when gluten is reintroduced in their diet.15,29 In this research, the symptoms were evaluated through two different methods, i.e., Celiac Symptoms Index Questionnaire (CSI) and the Gastrointestinal Symptom Rating Scale (GSRS). The CSI results are shown in Fig. 2 as a box-plot graphs due to the non-parametric distribution of data. CSI scores (Fig. 2) did not differ significantly at the baseline, thus confirming the homogeneity among the groups (A, B, and C). However, the symptoms were reported in all the groups in spite of the CD subjects following a GFD for a minimum of two years, as suggested by the median and quartile ranges in the three groups: 31 with an interquartile range from 22 to 40. After 4 weeks, all the subjects experienced a significant decrease in the CSI scores (P < 0.05, Friedmann test) and remained at their lowest value throughout the study, with a median value of 16–19 for 12 weeks (Fig. 2). This significant decrease, as evidenced by the Friedman test, could be due to different reasons: in control group A, it could be a result of the study protocol. Strict monitoring of the GFD, in fact, was imposed on all the groups (A, B, C) throughout the 12 weeks of the trial because patients were tested weekly through a Biocard Celiac Test for the rapid screening of the levels of anti-tissue transglutaminase IgA antibodies. On the other hand, in groups B and C, there was clearly an effect of GF eating, thus suggesting the possible effect of GF on symptoms’ recovery and the possible action on symptoms’ relief.
 | ||
| Fig. 2 Gluten friendly induces symptoms relief the box-whisker plot indicates CSI scores for the three trial groups (a, control group/gluten free group; b, 3 g of GF daily; c, 6 g of GF daily). Lower case letters (a; b; a,b) indicate significant differences (Friedmann test, p < 0.05). | ||
After the trial, the CSI scores increased, although the levels were lower than on week 0 of the study (Fig. 2). Such relapse in the symptoms could be a result of gluten ingestion, either deliberate or inadvertent, while on a GFD30 since this diet is socially troublesome, expensive, and compliance is problematic.
As reported above, the symptoms were also assessed through GSRS, which is based on a different prerequisite. With this second assessment, the symptoms were not recorded in line with the Bristol Stool Chart data of normal stool consistency (ESI Table 1†).
After 12 weeks of Gluten Friendly, the results show an uncoupling between the mucosal injury and inflammation in celiac subjects
Gluten-induced small intestinal mucosal injury is the gold standard for diagnosing celiac disease.31 During a gluten challenge, an inflammatory process with a dose-dependent accumulation of intraepithelial lymphocytes (IELs) is followed by villous atrophy with crypt hyperplasia.32Crypt hyperplasia is associated with an increased turnover of epithelial cells, marked by a higher expression of Cyclin B1 (CB1).33 Gut barrier permeability is altered with the infiltration of cytotoxic T-cells in the lamina propria.34 The total number of goblet cells producing Mucin 2 (MUC2) in the duodenal mucosa is reduced.35
To investigate the effects of GF on mucosal histology and inflammation, we analyzed small-bowel morphology (VH/CrD), intraepithelial densities of CD3+, celiac serology, i.e., EMA (anti-endomysium antibodies) and TGA (anti-transglutaminase antibodies), MUC2, CB1, gut barrier permeability, and a panel of proinflammatory cytokines. The absence/presence of the 33-mer gluten peptide in stools was assessed through the G12-ELISA immunoassay.
Duodenal mucosal deterioration was significant in groups B and C, compared to the placebo group A (Fig. 3a). However, the boxes and plots suggest a strong variability within each group. The median values of IELs at the end of the trial (Fig. 3b) were 26%, 38%, and 57% for groups A, B, and C, respectively; however, the differences among the groups were not significant and the box-plots suggest a strong variability within all the groups.
 | ||
| Fig. 3 VH/CrD and IEL at the end of trial in groups A, B and C. A, 0 g of GF daily; B, 3 g of GF daily; C, 6 g of GF daily. Morphometric analysis (A) measuring VH/CrD was made in well-oriented biopsy samples. A decrease in VH/CrD of 0.5 or more was considered significant and equivalent to clinical gluten sensitivity. Immunohistochemistry (B) of IELs was carried out through anti-CD3 antibody. An IEL increase of more than 30% was considered significant and equivalent to clinical gluten sensitivity. Measurements were independent of clinical information. | ||
MUC2 and CB1 were analyzed by one-way ANOVA (ESI Fig. 1 and 2†). The differences among the groups were not significant and each group showed high variability. We recorded similar findings for our cytokine panel (IL-2, IL-4, IL-6, and IL-8), TNF-α, and IFN-γ (ESI Fig. 3 and 4†).
EMA and TGA antibodies were never detected in the subjects from the placebo group A. In the GF groups, seroconversion occurred in 15% (B) and 47% (C) of the subjects after 4 weeks, which further increased to 42% (B) and 68% (C) after 12 weeks.
We utilized G12/33-mer assay to detect gluten in stools because this assay is generally used to assess if celiac patients consume gluten. The results are in Fig. 4 and are reported on a qualitative basis (presence/absence of 33-mer in the stool). As expected, due to the consumption of a gluten free-diet, 33-mer was always lacking on the stools of the patients of group A, while the 33-mer peptide in the stool samples was below the detection limit (absence) in 8 patients from group B, corresponding to 47% of the total. In the group C, 33-mer was below the detection limit in 10 subjects (55% of the total).
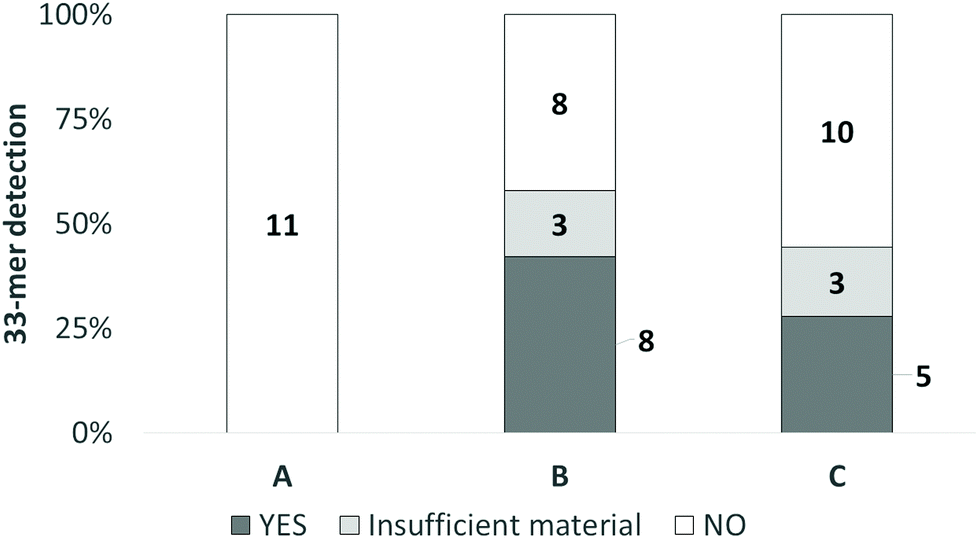 | ||
| Fig. 4 Detection of 33-mer peptide at the end of trial in groups A, B and C. A, 0 g of GF daily (control group); B, 3 g of GF daily; C, 6 g of GF daily. White indicates no 33-mer detection (NO); dark grey indicates 33-mer detection (YES); light grey indicates insufficient material during sampling (insufficient material). The number on the bars indicate the subjects per each class (NO; YES; insufficient material). | ||
In some patients, 33-mer was not detectable due to a technical issue (3 from B and 3 from C groups).
As a consequence, we could not establish the expected correlation between 33-mer detection and serology, nor between 33-mer detection and mucosal injury. Rather, we found partial uncoupling between the 33-mer peptide and both VH/CrD and antibodies.
We expected VH/CrD to correlate strongly with an increase in the IELs, an increase in the CB1, and a decrease in MUC2. Though there was a decrease in MUC2, VH/CrD was unrelated and uncoupled with both IELs and CB1. The partial correlation of VH/CrD with TG2 and EMA in our results was expected as it has been found that serum TG2 and EMA often underestimate the degree of VH/CrD.36
Finally, we expected VH/CrD to strongly correlate with an increase in the gut permeability and proinflammatory cytokines,37 which can disrupt the proteic components of tight junctions (TJs).38 However, gut permeability did not support this hypothesis; in fact, the differences between the gut permeability amongst the groups A, B, and C were not significant both before and at the end of the trial; in addition, the differences between the data before and after the trial were not significant (ESI Fig. 5,†), suggesting that GF did not cause the opening of TJs, in contrast to the CD literature. These results are in line with previous in vitro studies, where GF flour showed the ability to increase TEER in human intestinal goblet cells.10
Overall, the data suggested an uncoupling between VH/CrD and IEL, CB1, cytokines, gut permeability, and 33-mer detection in stools. While histology was consistent with well-established gluten challenge literature, all the subjects showed a high variability of data in some key parameters, eluding the strong two-class correlations that are usually established within the framework of untreated celiac disease.
Clustering through K-means shows an uncoupling of mucosal injury, inflammation markers, and 33-mer peptide detection
The new hypothesis that the subjects clustered around the “unknown variables” emerged. Therefore, we approached the data as a whole, regardless of the GF dosage. We searched for major factors at play through K-means analysis (Fig. 5) (ESI Tables 2 and 3†). | ||
| Fig. 5 Clustering through K-means shows an uncoupling of mucosal injury, inflammation markers and 33-mer peptide detection parameters above show mean values for VH/CrD decrease (A), MUC2 decrease (B), CB1 increase (C), IELs increase (D), and 33-mer peptide detection in K-means clusters. In (A), (B) and (D), bars represent 95% confidence intervals, whereas letters denote significant differences (one-way anova or Friedmann test). In (C) and (E), asterisks denote clusters belonging to homogeneous groups (one-way anova). K1, K2 and K3: clusters 1, 2 or 3 of K-means. | ||
K was set to 3 to reflect the number of groups in the study (A, B, C) and to avoid statistical artifacts. The input variables were VH/CrD, IEL, MUC2, CB1, and 33-mer. The permeability and cytokines were excluded as they did not present significant variations in the study groups. IEL, MUC2, CB1, and 33-mer underwent preliminary standardization; CB1, IELs, and MUC2 were converted to “relative increase/decrease”; 33-mer was converted to “relative detection”. Using relative increases was appropriate as all the subjects showed similar markers at the baseline. Preliminary standardization and conversion allowed for statistics based on homogeneous factors.
K-means calculations resulted in a new clustering of celiac subjects from groups A, B, and C. The details are shown in the ESI Tables 2 and 3.† Cluster 1 (K1) included 15 individuals (7 group C, 6 group B, and 2 group A); Cluster 2 (K2) included 11 individuals (5 from C and 6 from B), and Cluster 3 (K3) included 22 individuals (6 group C, 7 group B, and 9 group A). ESI Table 3† shows mean values of the categorical predictors of K-means, while Fig. 5 shows the statistical distribution of the celiac subjects for different parameters.
VH/CrD (Fig. 5a) experienced a 2-class trend. K1 and K2 were similar (VH/CrD at −1.07 and −1.28, respectively), whereas K3 differed significantly (VH/CrD at −0.10). A similar trend was found with MUC2 (Fig. 5b). K1 and K2 showed a decrease in MUC2 (−13.15% and −23.80%, respectively), while K3 showed an increase in MUC2 (+15.23%). K1 and K2 also showed an increase in CB1 (+16.88% and +20.10%, respectively), while K3 showed a decrease in CB1 (−3.12%) (Fig. 5c). These results enhanced the idea of a coupling/correlation among VH/CrD, MUC2, and CB1. However, these parameters did not correlate with IEL levels, which instead showed a 3-class trend. K1 showed the highest mean value of IELs (+97.80%), K2 showed an intermediate mean value (+46.22%), and K3 showed the smallest mean value (+13.52%) (Fig. 5d). All the subjects in K1 had IELs >30%. K2 showed a varied distribution: 4 subjects had IELs <30% and 7 subjects had IELs ranging from 40% to 80%. The vast majority of subjects in K3—specifically 19 out of 22—showed a value of IELs <30%. The remaining 3 subjects from K3 experienced a slight increase (32–42%). Among them were one subject on GFD (Group A) and 2 subjects eating GF. The number of celiac subjects with IELs <30% is particularly significant as any value of IELs >30% is clinically considered as a sign of inflammation.
In terms of 33-mer detection, the data showed a 3-class distribution (Fig. 5e). In K1, 47% (5 subjects) were negative to G12. In K2, 73% (8 subjects) were negative to G12 or G12 was not detectable. In K3, 91% (20 subjects) were negative to G12. Of those in K3, 11 were on a GF and 9 were eating GFD. In conclusion, the 33-mer gluten peptide could not be found in the stool samples of 65% (24 subjects) of celiac subjects eating GF.
Overall, K-means analysis outlined two trends—a two-class distribution for VH/CrD, MUC2, and CB1; a three-class distribution for IELs and 33-mer detection. From a statistical standpoint, different distributions indicate a lack of correlation among the groups of parameters analyzed for K1, K2, and K3. Such uncoupling is in contrast with the gold standard of celiac disease diagnosis, where gut mucosal damage and inflammation markers are strongly correlated.
K-means analysis evidenced that the GF subjects in K3 clustered with a vast majority of GFD subjects. In order to express percentiles of GF subjects only, we then excluded all the GFD subjects across the k-means clusters. The remaining 37 celiac subjects eating GF were distributed as follows: K1 35% (13 subjects); K2 30% (11 subjects); K3 35% (13 subjects).
K-means analysis indicated an uncoupling among the same key parameters as in the data analysis based on the GF dosage (Groups A, B, C). VH/CrD significantly decreased in K1 and K2. VH/CrD did not correlate with IEL increase in K2. This could suggest the possibility of a positive modulation of IELs in the celiac subjects in K2 (30% of all subjects eating GF). VH/CrD did not correlate with the detection of 33-mer in both K1 and K2. Conversely, low IEL increase and 33-mer absence strongly correlated in K3, suggesting a positive effect in celiac subjects in K3 (35% of all the subjects eating GF). Nevertheless, statistical analysis was not able to explain (i) why, despite the absence of 33-mer in K1 (47%) and in K2 (73%), the majority experienced intestinal damage; (ii) why intestinal damage in K2 is uncoupled with IELs after 12 weeks of eating GF; (iii) why celiac subjects in K1 and K2 experienced symptom relief despite showing histological damage.
Although our analysis was based on parameters that are widely shared in CD research,39 our data set did not show strong correlations typical of gluten-induced mucosal damage, which are well documented in previous literature. We, therefore, decided to deepen our study of the effects of GF bread on celiac subjects by exploring its interactions with the intestinal microbiota.
Gluten Friendly is digested by celiac subjects on the basis of their microbiota traits
Several studies have stressed the role of the microbiota in CD, hypothesizing that CD might be triggered by the additive effects of immunotoxic gluten peptides and intestinal dysbiosis in people with or without genetic susceptibilities.40Celiac subjects generally show typical microbial imprinting with a prevalence of some proinflammatory genera/family.2
In our study, we analyzed the stool samples to determine the microbiota variability of the study subjects at the baseline. Through genus/family detection, we set out to investigate whether the initial differences in the gut microbiota could be related to how GF affected the symptoms, histology, and serology. As a preliminary step, Operational Taxonomic Unit (OTU) from bioinformatics was converted into the numeric code as follows: 0 when the genus/family was lacking and 1 when the genus/family was recovered.
The data was analyzed using both the study groups (A, B, and C) and the k-means clusters (K1, K2, and K3) as the categorical factors. Only K3 was divided into two sub-clusters: “Friendly” (13 subjects eating GF bread) and “Free” (9 subjects on a GFD).
Recovery detection (%) of each family in each cluster is shown in Fig. 6a. When using the K-means clusters, the differences in the genus/family detection at the baseline were significant only for two genera: Slackia and Sutterella. The genus Sutterella belongs to Proteobacteria. Previous studies have suggested its role in atopic dermatitis41 and diabetes.42 An increase in Proteobacteria might contribute to nonspecific mucosal inflammation due to lipopolysaccharide (LPS) stimulation, and possibly predispose the host to a chronic inflammatory disease.43Slackia has been associated with a variety of imbalances, in particular, microbial dysbiosis connected to gastric carcinogenesis.44
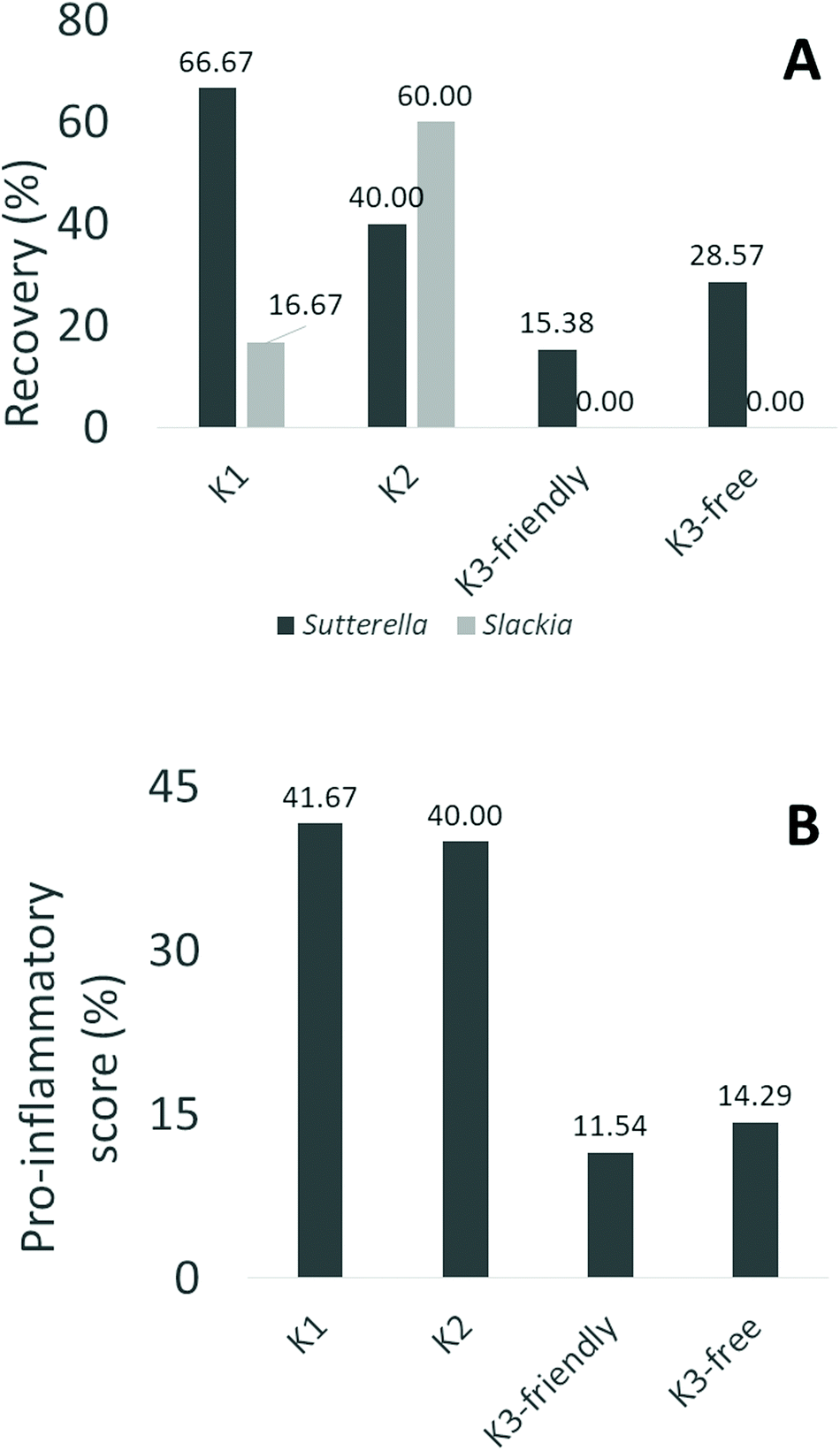 | ||
| Fig. 6 Presence of pathobionts in cluster 1 and 2. Figures above show the recovery of Slackia and Sutterella in the faecal microbiota of celiac subjects at the beginning of trial, distributed in K-means clusters and sub-clusters (A), and the standardized score of pro-inflammatory genera evaluated as the sum of Slackia and Sutterella (B). K1, K2 and K3: clusters 1, 2 or 3 of K-means. The word friendly and free indicate the sub-cluster of cluster 3 and the subjects receiving GF bread and gluten free bread. | ||
In our analysis, Sutterella was found in 66.67% of subjects in K1 and 40% of subjects in K2. In K3, recovery detection was significantly lower in both the “Friendly” (15.38%) and “Free” (28.57%) sub-clusters (Chi-square test, P < 0.05). Slackia was not found in any of the subjects in K3. The recovery detection was 16.67% in K1 and 60% in K2.
Recent studies have highlighted the role of gut microbes in determining the gluten immunogenicity. It has been shown that pathobionts such as P. aeruginosa isolated from CD patients cleaved the 33-mer peptide in such a manner that it activated gluten-specific T-cells. At the same time, the peptides could be further metabolized by duodenal Lactobacillus spp. to lose their immunogenicity.45 Different inflammatory pathways in the intestinal mucosal surface are activated by exposure to infectious agents that compete with beneficial microbes to adhere to the intestinal mucosa and disturb intestinal barrier function.40 A disturbed intestinal barrier leads to the exposure of intestinal immune cells to dietary antigens (i.e., gluten). Conversely, a healthy microbial ecology—defined as a balance between beneficial bacteria, harmful bacteria, and beneficial microbes—prevents the activation of inflammatory pathways and favors immune cell tolerance.46
Based on this literature, our hypothesis is that Slackia and Sutterella could jointly contribute to a proinflammatory response and interfere with the digestion of GF in celiac subjects. In order to assess their synergistic additive effect, we designed an arbitrary index. A ternary code was assigned to each subject: 0, when both Slackia and Sutterella were not detected; 50%, when only one genus was found; 100%, when both Slackia and Sutterella were found (ESI Table 4†).
The results of this index, called “standardized score of proinflammatory genera”, are shown in Fig. 6b. The score was 41.67% in K1 and 40% in K2. Instead, in K3, the score was significantly lower in both the “Friendly” (11.54%) and in the “Free” (14.29%) subclusters (P < 0.05, Chi-square test).
The different recovery of Slackia and Sutterella found in the stool samples of the celiac subjects at the baseline strongly correlated with the presence/absence of 33-mer in the K-means clusters (end of the study), i.e., the almost total absence of the 33-mer peptide in K3 and its significant presence in K2 and K1 (ESI Table 2†).
Therefore, it can be postulated that Slackia and Sutterella interfered with the digestion of GF in 53% of celiac subjects in K1 and 27% of celiac subjects in K2, where 33-mer was detected. Furthermore, Slackia and Sutterella may have predisposed the intestines to inflammation even in the remaining celiac subjects from K1 and K2, where 33-mer was not detected. Such hypothesis seems to be corroborated by the uncoupling between VH/CrD decrease and 33-mer presence in K1 and K2. Our hypothesis is that in the presence of Slackia and Sutterella, even traces of the 33-mer peptide (below 40 ppm) not detectable by G12 immunoassay could cause detrimental effects on the gut mucosa. This is in line with previous studies, highlighting how celiac sensitivity to very low amounts of gluten may vary depending on the intestinal microenvironment.47
Conversely, in the presence of a healthy microbial ecology, as observed in the subjects in the K3 “Friendly” sub-cluster (Fig. 6), butyrate-producing species were able to thrive and celiac gut microbiota endopeptidases were able to digest GF without detrimental effects on celiac gut mucosa.
Gluten Friendly shapes celiac microbiota toward intestinal homeostasis
The last step of this research was to focus on how GF administration affected the microbiota over time. Our analysis evidenced significant variations in the recovery of the following bacteria: Agathobacters pp (Fig. 7a), Eubacterium coprostanoligenes (Fig. 7b), Tyzzerella spp (Fig. 7c), Lactobacillus spp (Fig. 7d), and Intestinimonas spp (Fig. 7e). The data should be read as recovery variation (increase or decrease) after 12 weeks. The data is reported as the box-plot pictures with medians and quartiles as it is not possible to assume that they follow a normal distribution. | ||
| Fig. 7 Gluten friendly shapes gut microbiota towards homeostasis. Figures above show changes (increase or decrease of abundance at the end of the trial) in the relative abundance (%) of Agathobacter spp. (A), Eubacterium coprostanoligenes spp. (B), Tyzzerella spp. (C), Lactobacillus spp. (D) and Intestinimonas (E) after 12 weeks of trial. K1, K2 and K3: clusters 1, 2 or 3 of K-means. the word friendly and free indicate the sub-cluster of cluster 3 and the subjects receiving GF bread and gluten free bread. | ||
Fig. 7a shows the changes in the recovery of Agathobacter spp. Cluster K1 evidenced a median value of −0.27%, while the 3rd and 1st quartiles were 0 and −1.25% (box), respectively, with a minimum value of −3.1%. Such distribution suggests that celiac subjects in K1 (at least 75% of them) experienced a decrease in the Agathobacter spp. population after 3 months. A similar trend was found in K2, where the median value of variation was 0%, 3rd and 1st quartiles of respectively 0% and −0.8%, with a minimum value of −2.1%. In K3, the “Friendly” data evidenced a significant difference. The median value was 0.4%. The 1st and 3rd quartiles were 0 and 0.8%, respectively, with a maximum value of 4.3%. The range from the 1st quartile to maximum (0–4.3%) suggests that for this cluster, 75% of the subjects in K3 “Friendly” experienced an increase in Agathobacter spp.
Fig. 7b shows the recovery variation of Eubacterium coprostanoligenes. The recovery variation was not significant. K1 experienced a strong variation, while the box of K3 “Friendly” (median, 1st, and 3rd quartiles around 0%) suggested a possible homeostasis for at least 50% of the celiac subjects.
Eubacterium coprostanoligenes is an anaerobic, Gram-positive coccobacillus with cholesterol-reducing properties.48
Fig. 7c shows the recovery variation of Tyzzerella spp., evidencing homeostasis for both K3 “Friendly” and “Free” clusters (box, maximum, and median to 0%), while K1 and K2 clusters evidenced a significant variation with an increasing trend; the differences between the “Friendly”/“Free” clusters and K2/K1 clusters were significant (P, 0.0488). Previous studies have linked distinct Tyzzerella species to the inflammatory milieu of Crohn's Disease and functional gastrointestinal disorders in autistic children,49 respectively.
Fig. 7d shows the recovery variation of Lactobacillus spp., showing an increase (P, 0.0217) in K2 (median and 1st quartile 0.03%; 3rd quartile, 0.15%; maximum, 1.0%).
Fig. 7e shows the recovery variation of Intestinimonas spp., showing an increasing trend in K1.
Lactobacillus is a well-known genus with various beneficial properties. It can contribute to gut re-equilibration and to shape the microbiota toward homeostasis, with a protective effect against inflammation. Moreover, Lactobacillus spp. might promote the homeostasis of the gut mucus layer through L-Ornithine (L-Orn) production. L-Orn stimulates L-tryptophan (Trp) metabolism to produce aryl hydrocarbon receptor (AhR) ligands in the gut epithelial cells, which induce the accumulation of RORγt(+) IL-22(+) ILC3 in the gut tissues.50
Significant variations of Lactobacillus spp. in the microbiota of celiac subjects, as observed in cluster K2, confirmed the in vivo bifidogenic effect of GF. In fact, in previous studies,8,9 GF has shown its in vitro ability to stimulate both Lactobacilli growth and metabolism. Significant Lactobacillus spp. increase could play a role in the unexpected uncoupling between VH/CrD vs. IELs and VH/CrD vs. 33-mer in K2, after 12 weeks of eating GF. This could also explain why celiac subjects experienced a consistent relief in the symptoms. Although the data on microbiota variations (Fig. 7d) show that K2 was not in homeostasis, it could be hypothesized that the Lactobacilli were metabolizing GF and exploiting it as a source of energy. In turn, Lactobacilli metabolism contributed to taming the inflammation. Studies have suggested that gut commensals, including Lactobacillus, Bifidobacterium, Bacteroides, Clostridium, and Streptococcus, as well as bacterial metabolites such as butyric acid and propionic acid, may program Treg cells in the intestine toward tolerogenicity, helping the host with the maturation and homeostasis of the immune system.51
Furthermore, the increase in Intestinimonas and Agathobacter in both K1 and the K3 “Friendly” cluster, seems to point in the same direction. Both the species produce butyrate, contributing to a “healthy ecology” of the gut and to its homeostasis. Agathobacter uses acetate to produce butyrate,52 while Intestinimonas grows by metabolizing amino acids, in particular, lysine and glutamate, and produces butyrate.53
Overall, these results confirm the hypothesis beyond clustering: time may be a crucial factor in enabling GF interaction with celiac gut mucosa by reshaping its microbiota toward homeostasis. The idea of a re-shaping, or at least beginning a reshaping, was supported by an overview of all the data of microbiota composition, at least for the most important genera for CD (ESI Table 5†). As reported elsewhere, the data did not follow a parametric distribution; therefore, the information was synthesized through position indices (median, quartiles, minimum, and maximum). A check of the quantitative composition for 54 genera/groups suggests for some of them a re-shaping from the beginning to the end of the trial; this information was also reported in ESI Fig. 6 and 7,† which reports the median values for all the genera.
For some genera, the figures show a shift in the median values to higher or lower values compared to the beginning for most. Sometimes, the differences were significant, while for other genera, there was a shift in the median and quartiles, while the maximum and minimum of distribution were not affected probably due to some subjects that did not experience a shift or restoration of the microbiota, for example, Alistipes, Blautia, (in ESI Fig. 6 part A;† the genera correspond to the numbers 10 and 20), Lachnoclostridium, Faecalibacterium, Escherichia/Shigella (in ESI Fig. 6 part B;† the genera correspond to the numbers 25, 39, 53). For other genera, there was a significant shift at least for the interquartile range; as an example, ESI Fig. 7† shows the results for Eisenbergiella (part A) and for Saccharimonadaceae (part B).
The chart of Eisengergiella confirms our hypothesis reported for Slackia and Sutterella; the initial printing of the microbiota in terms of the pro-inflammatory and beneficial microorganisms could affect the response to GF. At the baseline, the median and quartile 1 of cluster 1 were 0.0% and the quartile 3 was 0.053%, thus suggesting that Eisenbergiella was almost missing in at least 75% of the subjects of this cluster; on the other in cluster 3, at the baseline, the median was 0.109%, quartile 3 at 0.795%, and the maximum of the distribution was 1.376%, thus highlighting a relatively higher abundance in the subjects of cluster 3 receiving GF. Another interesting evidence was the trend of K2, which showed an intermediate trend between K1 and K3.
Eisenbergiella is probably related to eubiosis because it can produce butyrate, acetate, lactate, and succinate as major metabolic products, with a trophic effect on the mucosa.54 Its higher abundance in K3 and K2 subjects should be read in an opposite way of Slackia: while the latter genus is a marker of pro-inflammatory conditions, Eisenbergiella could be a marker of a possible positive effect of GF.
Fig. S7B† shows the shift in Saccharimonadaceae and the different trends between cluster 1 (K1) and cluster 3 (K3); for this family, GF exerted a modulating effect with a strong reduction after the trial in cluster 3 receiving Gluten Friendly, as suggested by median and quartile 3, both of which were 0.0% (family lacking in at least 75% of subjects). The role of Saccharimonadaceae was not clear but a recent evidence suggests a correlation of this family with inflammatory responses and high reactions of the immune system.55
Gut microbiota is a complex entity, composed of bacteria, fungi, and viruses; in this research, we have addressed only the effect of GF on the bacteria because of the preliminary experiments and their evidence on some target genera (such as Lactobacillus and Bifidobacterium). However, the fungi could also play a role in the modulation of the gut microbiota and on its positive or negative shifts; therefore, further investigations are required in the future to address this issue.
Conclusion
Gluten friendly has shown in vivo unprecedented characteristics that elude well-established literature on the gluten challenge. All 37 celiac subjects eating 3 g (Group B) or 6 g (Group C) of GF for 12 weeks experienced no symptoms or even an improvement of symptoms, according to CSI scores and GSRS. The 33-mer gluten peptide could not be found or was undetectable in 24 celiac subjects eating GF, suggesting the possibility of digestion. Based on histology and antibody seroconversion, GF appeared to be disease-inducing in a majority of study subjects over the course of 12 weeks. However, the GF data set did not show strong correlations typical of gluten-induced mucosal damage. For example, VH/CrD did not correlate with IELs >30%, 33-Mer in stools, and the symptoms. K-means analysis evidenced the possibility of celiac subjects clustering (K1, K2, K3) around unknown parameters independent of the GF dosage. Recent studies have highlighted the role of gut microbes in determining the gluten immunogenicity. The microbiota variations of specific proinflammatory (Slackia and Sutterella) and healthy and butyrate-producing bacteria (Lactobacillus spp, Agathobacter spp, and Intestinimonas spp) evidenced a correlation with the K-means clustering of celiac subjects, namely, lack of symptoms, no 33-mer detection, and IELs <30%. Proinflammatory gut ecology showed an adverse role in the digestion and tolerance of GF in the study subjects. However, our research suggests that GF shapes the microbiota toward gut homeostasis in celiac subjects by boosting healthy butyrate-producing populations. Time may be a crucial factor in enabling GF to immunomodulate the celiac gut mucosa, along with the initial composition of the microbiota. This result will be shared in the future with volunteers and subjects involved in other studies because our feeling is that further experiments are required to elucidate the mode of action of GF, its modulatory effect on the microbiota, the reasons for the uncoupling between the mucosal data, and other evidences (symptoms, IEL, and gluten tolerance).Apart from the results, another conclusion of this research is that, although not clinically validate yet, the quali-quantitative composition of the microbiota should be considered for such situations, such as CD.
Author contributions
Design research: Carmen Lamacchia, Angelo Andriulli, Antonio Bevilacqua, Orazio Palmieri and Maria Rosaria Corbo; Methodology: Carmen Lamacchia, Angelo Andriulli, Antonio Bevilacqua, Orazio Palmieri, Maria Rosaria Corbo and Milena Sinigaglia; Software: Antonio Bevilacqua, Orazio Palmieri, Michele Tricarico, Leonardo Petruzzi; Validation: Antonio Bevilacqua, Orazio Palmieri, Carmen Lamacchia; Formal analysis: Antonio Bevilacqua, Orazio Palmieri, Stefano Castellana, Tommaso Mazza and Leonardo Petruzzi; Conducted research: Orazio Palmieri, Anna Latiano, Rosanna Fontana, Domenica Gioffreda, Anna Panza, Claudia Menzaghi, Elvira Grandone, Lazzaro di Mauro, Ivana Decina, Michele Tricarico, Daniela Musaico, Alina Popp, Juha Taavela, Leonardo Petruzzi; Resources: Angelo Andriulli, Carmen Lamacchia, Markku Maki, Jorma Isola; Data curation: Angelo Andriulli, Orazio Palmieri, Antonio Bevilacqua and Carmen Lamacchia; Writing-original draft: Carmen Lamacchia, Antonio Bevilacqua and Maria Rosaria Corbo; Writing – review & editing: Carmen Lamacchia, Antonio Bevilacqua, Maria Rosaria Corbo, Milena Sinigaglia, Angelo Andriulli, Orazio Palmieri, Markku Maki, Jorma Isola, Juha Taavela and Daniela Musaico; Visualization: Carmen Lamacchia, Antonio Bevilacqua, Maria Rosaria Corbo; Supervision: Angelo Andriulli and Carmen Lamacchia; Project administration: Carmen Lamacchia and Angelo Andriulli; Funding acquisition: Carmen Lamacchia.Data and materials availability
All data is available in the main text or the ESI.† The raw data that is presented in this paper is available upon request from Ospedale Casa Sollievo della Sofferenza in San Giovanni Rotondo, Italy, without restrictions.Conflicts of interest
Angelo Andriulli, Orazio Palmieri, Anna Latiano, Rosanna Fontana, Domenica Gioffreda, Stefano Castellana, Tommaso Mazza, Anna Panza, Claudia Menzaghi, Elvira Grandone, Lazzaro di Mauro, Markku Mäki, Jorma Isola, Alina Popp, Juha Taavela declare no competing interest; Ivana Decina and Michele Tricarico declare that their institution has a patent related to this work: “Method for detoxification of gluten proteins from grains of cereals and related medical uses” (Italian Priority Patent No. 102015000084813, Patent Cooperation Treaty PCT/EP2016/0815899, 2016). Daniela Musaico, Antonio Bevilacqua, Leonardo Petruzzi, Milena Sinigaglia, Maria Rosaria Corbo declare that their institution has a patent related to this work: “Method for detoxification of gluten proteins from grain of cereals” (Patent Cooperation Ttreaty PCT/IB2013/000797, 2013); Carmen Lamacchia declares that she is a founder and a share holder, at about 60%, of New Gluten World s.r.l since april 2015. She serves as a CEO of the New Gluten World since April 2015. She is the inventor of the following patents related to this work: “Method for detoxification of gluten proteins from grain of cereals” (Patent Cooperation Treaty PCT/IB2013/000797, 2013) and “Method for detoxification of gluten proteins from grains of cereals and related medical uses” (Italian Priority Patent No. 102015000084813, Patent Cooperation Treaty PCT/EP2016/0815899, 2016).Acknowledgements
We thank all celiac volunteers and their families for participating in this study, as well as the physicians, nurses, and medical staff who helped with recruitment at Fondazione IRCCS Casa Sollievo della Sofferenza Hospital. We thank the following individuals for their support and assistance throughout the study: Corritore Giuseppe, Gentile Annamaria, Tiziana Latiano, Ercolino Filomena, Antonio Ippolito, Fulvio Spirito, Marco Gentile, Francesco Perri, Lucia Salvemini, Gennaro Vecchione, Mauro Truglio and Giuseppe Cappucci. Additional thanks to Storyhatch Srls for supporting the manuscript with editing and proof-reading. Obtained funding: This research was accomplished with the financial support of Casillo Group S.p.a. (Corato, Italy). This project has also received funding from the European Union's Horizon 2020 research and innovation program, under grant agreement no. 732640. Disclaimer: the contents of this publication are the sole responsibility of the authors, and do not necessarily reflect the opinion of the European Union.References
- I. Nadal, E. Donat, C. Ribes-Koninckx, Y. Calabuig and M. Sanz, Imbalance in the composition of the duodenal microbiota of children with coeliac disease, J. Med. Microbiol., 2007, 56, 1669–1674 CrossRef CAS PubMed.
- G. Marasco, A. R. Di Biase, R. Schiumerini, L. H. Eusebi, L. Iughetti, F. Ravaioli, E. Scaioli, A. Colecchia and D. Festi, Gut microbiota and celiac disease, Dig. Dis. Sci., 2016, 61, 1461–1472 CrossRef CAS PubMed.
- E. Sánchez, E. Donat, C. Ribes-Koninckx, M. Calabuig and Y. Sanz, Intestinal Bacteroides species associated with coeliac disease, J. Clin. Pathol., 2010, 63, 1105–1111 CrossRef PubMed.
- K. A. Bascuñán, M. Araya, L. Roncoroni, L. Doneda and L. Elli, Dietary gluten as a conditioning factor in celiac disease, Adv. Nutr., 2020, 11, 160–174 Search PubMed.
- C. Lamacchia, L. Landriscina and P. D'Agnello, Changes in wheat kernel proteins induced by microwave treatment, Food Chem., 2016, 197, 634–640 CrossRef CAS PubMed.
- C. Lamacchia, A. Di Luccia and C. Gianfrani, Method for the detoxification of gluten proteins from grains of cereals, Patent Cooperation TreatyPCT/IB2013/000797, 2015 Search PubMed.
- L. Landriscina, P. D'Agnello, A. Bevilacqua, M. R. Corbo, M. Sinigaglia and C. Lamacchia, Impact of Gluten-Friendly™ technology on wheat kernel endosperm and gluten protein structure in seeds by light and electron microscopy, Food Chem., 2017, 221, 1258–1268 CrossRef CAS PubMed.
- A. Bevilacqua, A. Costabile, T. Bergillos-Meca, I. Gonzalez, L. Landriscina, E. Ciuffreda and C. Lamacchia, Impact of Gluten-Friendly bread on the metabolism and function of in vitro gut microbiota in healthy human and coeliac subjects, PLoS One, 2016, 11, e0162770 CrossRef PubMed.
- A. Costabile, T. Bergillos-Meca, L. Landriscina, A. Bevilacqua, I. Gonzalez-Salvador, M. R. Corbo, L. Petruzzi, M. Sinigaglia and C. Lamacchia, An in vitro fermentation study on the effects of Gluten FriendlyTM bread on microbiota and short chain fatty acids of fecal samples from healthy and celiac subjects, Front. Microbiol., 2017, 8, 1722 CrossRef PubMed.
- C. Lamacchia, D. Musaico, M. E. Henderson, T. Bergillos-Meca, M. Roul, L. Landriscina, I. Decina, G. Corona and A. Costabile, Temperature-treated gluten proteins in Gluten-FriendlyTM bread increase mucus production and gut-barrier function in human intestinal goblet cells, J. Funct. Foods, 2018, 48, 507–514 CrossRef CAS.
- J. Gervasoni, A. Schiattarella, V. Giorgio, A. Primiano, C. Russo, V. Tesori, F. Scaldaferri, A. Urbani, C. Zuppi and S. Persichilli, Validation of an LC-MS/MS method for urinary lactulose and mannitol quantification: results in patients with irritable bowel syndrome, Dis. Markers, 2016, 2016, 5340386 CrossRef PubMed.
- C. Lamacchia, Method for detoxification of gluten protein from grains of cereals and related medical uses, Italian priority patent no102015000084813, Patent Cooperation Treaty PCT/EP2016/0815899, 2016 Search PubMed.
- J. Taavela, O. Koskinen, H. Huhtala, M. L. Lähdeaho, A. Popp, K. Laurila, P. Collin, K. Kaukinen, K. Kurppa and M. Mäki, Validation of morphometric analyses of small-intestinal biopsy readouts in celiac disease, PLoS One, 2013, 8, e76163 CrossRef CAS PubMed.
- V. J. Tuominen and J. Isola, Linking whole-slide microscope images with DICOM by using JPEG2000 interactive protocol, J. Digital Imaging, 2010, 23, 454–462 CrossRef PubMed.
- M. L. Lähdeaho, M. Mäki, K. Laurila, H. Huhtala and K. Kaukinen, Small-bowel mucosal changes and antibody responses after low- and moderate-dose gluten challenge in celiac disease, BMC Gastroenterol., 2011, 11, 129 CrossRef PubMed.
- V. J. Tuominen, S. Ruotoistenmäki, A. Viitanen, M. Jumppanen and J. Isola, ImmunoRatio: a publicly available web application for quantitative image analysis of estrogen receptor (ER), progesterone receptor (PR), and Ki-67, Breast Cancer Res., 2010, 12, R56 CrossRef PubMed.
- J. Gervasoni, A. Primiano, C. Graziani, F. Scaldaferri, A. Gasbarrini, A. Urbani and S. Persichilli, Validation of UPLC-MS/MS method for determination of urinary lactulose/mannitol, Molecules, 2018, 23, 2705 CrossRef PubMed.
- 16S metagenomic sequencing library preparation, 2013, https://support.illumina.com/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf.
- D. A. Leffler, M. Dennis, J. E. George, S. Jamma, E. F. Cook, D. Schuppan and C. P. Kelly, A validated disease-specific symptom index for adults with celiac disease, Clin. Gastroenterol. Hepatol., 2009, 7, 1328–1334 CrossRef PubMed.
- D. A. Revicki, M. Wood, I. Wiklund and J. Crawley, Reliability and validity of the Gastrointestinal Symptom Rating Scale in patients with gastroesophageal reflux disease, Qual. Life Res., 1998, 7, 75–83 CrossRef CAS PubMed.
- M. Srinivas, V. Srinivasan, M. Jain, C. S. R. Shanthi, V. Mohan and V. Jayanthi, A cross-sectional study of stool form (using Bristol stool chart) in an urban South Indian population, JGH Open, 2019, 3, 464–467 CrossRef PubMed.
- S. Andrews, FastQC: A quality control tool for high throughput sequence data, http://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
- M. P. Cox, D. A. Peterson and P. J. Biggs, SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data, BMC Bioinf., 2010, 11, 485 CrossRef PubMed.
- J. G. Caporaso, J. Kuczynski, J. Stombaugh, K. Bittinger, F. D. Bushman, E. K. Costello, N. Fierer, A. G. Peña, J. K. Goodrich, J. I. Gordon, G. A. Huttley, S. T. Kelley, D. Knights, J. E. Koenig, R. E. Ley, C. A. Lozupone, D. McDonald, B. D. Muegge, M. Pirrung, J. Reeder, J. R. Sevinsky, P. J. Turnbaugh, W. A. Walters, Y. T. Widmann, J. Zaneveld and R. Knight, QIIME allows analysis of high-throughput community sequencing data, Nat. Methods, 2010, 7, 335–336 CrossRef CAS PubMed.
- B. J. Callahan, P. J. McMurdie, M. J. Rosen, A. W. Han and A. J. Johnson, Holmes S P. DADA2: High-resolution sample inference from Illumina amplicon data, Nat. Methods, 2016, 13, 581–583 CrossRef CAS PubMed.
- P. Yilmaz, L. W. Parfrey, P. Yarza, J. Gerken, E. Pruesse, C. Quast, T. Schweer, J. Peplies, W. Ludwig and F. O. Glöckner, The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks, Nucleic Acids Res., 2014, 42, 643–648 CrossRef PubMed.
- M. J. Anderson, Permutational Multivariate Analysis of Variance (PERMANOVA), Wiley StatsRef: Statistics Reference Online, 2017, vol. 2017, pp. 1–15 Search PubMed.
- S. Mandal, W. Van Treuren, R. A. White, M. Eggesbø, R. Knight and S. D. Peddada, Analysis of composition of microbiomes: a novel method for studying microbial composition, Microb. Ecol. Health Dis., 2015, 26, 27663 Search PubMed.
- D. Leffler, D. Schuppan, K. Pallav, R. Najarian, J. D. Goldsmith, J. Hansen, T. Kabbani, M. Dennis and C. P. Kelly, Kinetics of the histological, serological and symptomatic responses to gluten challenge in adults with coeliac disease, Gut, 2013, 62, 996–1004 CrossRef CAS PubMed.
- C. Catassi, E. Fabiani, G. Iacono, C. D'Agate, R. Francavilla, F. Biagi, U. Volta, S. Accomando, A. Picarelli, I. De Vitis, G. Pianelli, R. Gesuita, F. Carle, A. Mandolesi, I. Bearzi and A. Fasano, A prospective, double-blind, placebo-controlled trial to establish a safe gluten threshold for patients with celiac disease, Am. J. Clin. Nutr., 2007, 85, 160–166 CrossRef CAS PubMed.
- A. Rostom, J. A. Murray and M. F. Kagnoff, American Gastroenterological Association (AGA) Institute technical review on the diagnosis and management of celiac disease, Gastroenterology, 2006, 131, 1981–2002 CrossRef PubMed.
- M. N. Marsh, Crowe P T. Morphology of the mucosal lesion in gluten sensitivity, Baillieres Clin. Gastroenterol., 1995, 9, 273–293 CrossRef CAS PubMed.
- K. C. Niranjan, A. Tayaar, G. S. Kumar, R. Krishnapillai, K. Hallikeri and S. Hunasgi, Immunohistochemical expression of cyclin B1 in epithelial hyperplasia, dysplasia and oral squamous cell carcinomas - a comparative study, J. Clin. Diagn. Res., 2016, 10, 85–90 Search PubMed.
- J. König, J. Wells, P. D. Cani, C. L. García-Ródenas, T. MacDonald, A. Mercenier, J. Whyte, F. Troost and R. J. Brummer, Human intestinal barrier function in health and disease, Clin. Transl. Gastroenterol., 2016, 7, e196 CrossRef PubMed.
- D. Cardoso-Silva, D. Delbue, A. Itzlinger, R. Moerkens, S. Withoff, F. Branchi and M. Schumann, Intestinal barrier function in gluten-related disorders, Nutrients, 2019, 11, 2325 CrossRef CAS PubMed.
- J. A. Silvester, S. Kurada, A. Szwajcer, C. P. Kelly, D. A. Leffler and D. R. Duerksen, Tests for serum transglutaminase and endomysial antibodies do not detect most patients with celiac disease and persistent villous atrophy on gluten-free diets: a metaanalysis, Gastroenterology, 2017, 153, 689–701 CrossRef PubMed.
- B. L. Kelsall and M. Rescigno, Mucosal dendritic cells in immunity and inflammation, Nat. Immunol., 2004, 5, 1091–1095 CrossRef CAS PubMed.
- A. Fasano, T. Not, W. Wang, S. Uzzau, I. Berti, A. Tommasini and S. E. Goldblum, Zonulin, a newly discovered modulator of intestinal permeability, and its expression in coeliac disease, Lancet, 2000, 355, 1518–1519 CrossRef CAS.
- J. F. Ludvigsson, C. Ciacci, P. H. Green, K. Kaukinen, I. R. Korponay-Szabo, K. Kurppa, J. A. Murray, K. E. A. Lundin, M. J. Maki, A. Popp, N. R. Reilly, A. Rodriguez-Herrera, D. S. Sanders, D. Schuppan, S. Sleet, J. Taavela, K. Voorhees, M. M. Walker and D. A. Leffler, Outcome measures in coeliac disease trials: the Tampere recommendations, Gut, 2018, 67, 1410–1424 CrossRef CAS PubMed.
- A. M. Chander, H. Yadav, S. Jain, K. Bhadada and D. K. Dhawan, Cross-talk between gluten, intestinal microbiota and intestinal mucosa in celiac disease: recent advances and basis of autoimmunity, Front. Microbiol., 2018, 9, 2597 CrossRef PubMed.
- S. Reddel, F. Del Chierico, A. Quagliariello, S. Giancristoforo, P. Vernocchi, A. Russo, A. Fiocchi, P. Rossi, L. Putignani and M. El Hachem, Gut microbiota profile in children affected by atopic dermatitis and evaluation of intestinal persistence of a probiotic mixture, Sci. Rep., 2019, 9, 4996 CrossRef PubMed.
- K. H. Allin, V. Tremaroli, R. Caesar, B. A. H. Jensen, M. T. F. Damgaard, M. I. Bahl, T. R. Licht, T. H. Hansen, T. Nielsen, T. M. Dantoft, A. Linneberg, T. Jørgensen, H. Vestergaard, K. Kristiansen, P. W. Franks, IMI-DIRECT consortium, T. Hansen, F. Bäckhed and O. Pedersen, Aberrant intestinal microbiota in individuals with prediabetes, Diabetologia, 2018, 61, 810–820 CrossRef PubMed.
- J. L. Round and S. K. Mazmanian, The gut microbiota shapes intestinal immune responses during health and disease, Nat. Rev. Immunol., 2009, 9, 313–323 CrossRef CAS PubMed.
- O. O. Coker, Z. Dai, Y. Nie, G. Zhao, L. Cao, G. Nakatsu, W. K. Wu, S. H. Wong, Z. Chen, J. J. Y. Sung and J. Yu, Mucosal microbiome dysbiosis in gastric carcinogenesis, Gut, 2018, 67, 1024–1032 CrossRef CAS PubMed.
- A. Caminero, H. J. Galipeau, J. L. McCarville, C. W. Johnston, S. P. Bernier, A. K. Russell, J. Jury, A. R. Herran, J. Casqueiro, J. A. Tye-Din, M. G. Surette, N. A. Magarvey, D. Schuppan and E. F. Verdu, Duodenal bacteria from patients with celiac disease and healthy subjects distinctly affect gluten breakdown and immunogenicity, Gastroenterology, 2016, 151, 670–683 CrossRef CAS PubMed.
- P. M. Smith, M. R. Howitt, N. Panikov, M. Michaud, C. A. Gallini, M. Bohlooly-Y, J. N. Glickman and W. S. Garrett, The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis, Science, 2013, 341, 569–573 CrossRef CAS PubMed.
- G. Serena, S. Yan, S. Camhi, S. Patel, R. S. Lima, A. Sapone, M. M. Leonard, R. Mukherjee, B. J. Nath, K. M. Lammers and A. Fasano, Proinflammatory cytokine interferon-gamma and microbiome-derived metabolites dictate epigenetic switch between forkhead box protein 3 isoformsin coeliac disease, Clin. Exp. Immunol., 2017, 187, 490–506 CrossRef CAS PubMed.
- L. Li, S. M. Batt, M. Wannemuehler, A. Dispirito and D. C. Beitz, Effect of feeding of a cholesterol-reducing bacterium, Eubacterium coprostanoligenes, to germ-free mice, Lab. Anim. Sci., 1998, 48, 253–255 CAS.
- R. A. Luna, N. Oezguen, M. Balderas, A. Venkatachalam, J. K. Runge, J. Versalovic, J. Veenstra-VanderWeele, G. M. Anderson, T. Savidge and K. C. Williams, Distinct microbiome-neuroimmune signatures correlate with functional abdominal pain in children with autism spectrum disorder, Cell. Mol. Gastroenterol. Hepatol., 2016, 3, 218–230 CrossRef PubMed.
- H. Qi, Y. Li, H. Yun, T. Zhang, Y. Huang, J. Zhou, H. Yan, J. Wei, Y. Liu, Z. Zhang, Y. Gao, Y. Che, X. Su, D. Zhu, Y. Zhang, J. Zhong and R. Yang, Lactobacillus maintains healthy gut mucosa by producing L-Ornithine, Commun. Biol., 2019, 2, 171 CrossRef PubMed.
- Y. Furusawa, Y. Obata, S. Fukuda, T. A. Endo, G. Nakato, D. Takahashi, Y. Nakanishi, C. Uetake, K. Kato, T. Kato, M. Takahashi, N. N. Fukuda, S. Murakami, E. Miyauchi, S. Hino, K. Atarashi, S. Onawa, Y. Fujimura, T. Lockett, J. M. Clarke, D. L. Topping, M. Tomita, S. Hori, O. Ohara, T. Morita, H. Koseki, J. Kikuchi, K. Honda, K. Hase and H. Ohno, Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells, Nature, 2013, 504, 446–450 CrossRef CAS PubMed.
- J. A. Rosero, J. Killer, H. Sechovcová, J. Mrázek, O. Benada, K. Fliegerová, J. Havlík and J. Kopečný, Reclassification of Eubacterium rectale (Hauduroy, et al. 1937) Prévot 1938 in a new genus Agathobacter gen. nov. as Agathobacter rectalis comb. nov., and description of Agathobacter ruminis sp. nov., isolated from the rumen contents of sheep and cows, Int. J. Syst. Evol. Microbiol., 2016, 66, 768–773 CrossRef CAS PubMed.
- T. P. Bui, J. Ritari, S. Boeren, P. de Waard, C. M. Plugge and W. M. de Vos, Production of butyrate from lysine and the Amadori product fructoselysine by a human gut commensal, Nat. Commun., 2015, 6, 10062 CrossRef CAS PubMed.
- I. Amir, P. Bouvet, C. Legeay, U. Gophna and A. Weinberger, Eisenbergiella tayi gen. nov., sp. nov., isolated from human blood, Int. J. Syst. Evol. Microbiol., 2014, 64, 907–914 CrossRef CAS PubMed.
- D. A. Elhag, M. Kumar and S. Al-Khodor, Exploring the triple interaction between the host Genome, the epigenome, and the gut microbiome in type 1 diabetes, Int. J. Mol. Sci., 2021, 22, 125 CrossRef CAS PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d1fo00490e |
| ‡ A. Andriulli, A. Bevilacqua and O. Palmieri equally contributed to this work, while C. Lamacchia and M.R. Corbo are co-corresponding authors. |
| This journal is © The Royal Society of Chemistry 2022 |