
Mechanisms of growth-promotion and Se-enrichment in Brassica chinensis L. by selenium nanomaterials: beneficial rhizosphere microorganisms, nutrient availability, and photosynthesis†
Chuanxi
Wang
 ab,
Le
Yue
ab,
Bingxu
Cheng
ab,
Feiran
Chen
ab,
Xiaoli
Zhao
c,
Zhenyu
Wang
*ab and
Baoshan
Xing
d
ab,
Le
Yue
ab,
Bingxu
Cheng
ab,
Feiran
Chen
ab,
Xiaoli
Zhao
c,
Zhenyu
Wang
*ab and
Baoshan
Xing
d
aInstitute of Environmental Processes and Pollution Control, and, School of Environment and Civil Engineering, Jiangnan University, Wuxi, Jiangsu 214122, China. E-mail: wang0628@jiangnan.edu.cn
bJiangsu Key Laboratory of Anaerobic Biotechnology, Jiangnan University, Wuxi, Jiangsu 214122, China
cChinese Research Academy of Environmental Sciences, Beijing, 100012, China
dStockbridge School of Agriculture, University of Massachusetts, Amherst, MA 01003, USA
First published on 25th November 2021
Abstract
Maintaining a suitable selenium (Se) content in food is particularly important for human health. However, the mechanisms of uptake and enrichment of Se ENMs in crops are still unclear. Herein, Se engineered nanomaterials (Se ENMs) (size: 62.3 ± 14.6 nm and surface charge: −34.4 ± 1.4 mV) were synthesized and used as nanofertilizers for Se-fortified vegetables. The results demonstrated that the Se content and yield were increased by 338.0% and 19.8%, respectively, in Brassica chinensis L. through soil application of Se ENMs (0.5 mg kg−1). The Se content in vegetables increased up to 32.8 μg/100 g (7.5 μg/100 g for the control), which could provide the daily recommended Se intake (55–400 μg per day) for humans. Modification of the slightly alkaline soil with the Se ENMs improved beneficial rhizosphere microbiomes (Pseudomonas and Bacillus), which resulted in the plants accumulating more low molecular weight compounds (betaine, proline, glycine, norleucine, urocanic acid and indole-3-acrylic acid) with an increase in the Se content of the plant by 264.9%. Moreover, the nutrient accumulation in leaves promoted photosynthesis (16.7%) and increased the carbohydrate content (6.5%). Also, the expression of carbohydrate transport-related genes (BnSUC1,1, BnSUC1,4, and BnSWEET10,2) was up-regulated 52.2, 53.2 and 76.3-fold, respectively, promoting root growth and improving rhizosphere microbiome and nutrient availability. Therefore, such mutual benefits between leaves and roots using ENMs could provide an alternative model for cultivating Se-enriched crops.
Environmental significanceNano-enabled agriculture is providing solutions for sustainable agriculture. Our results showed that Se engineered nanomaterials, through soil application, could regulate the rhizosphere microbiome and nutrient availability to improve photosynthesis, the yield and the Se content of crops. Thus, Se engineered nanomaterials as nanofertilizers have the potential for replacing/reducing the use of traditional fertilizers in agriculture, while still ensuring crop yield and quality. These findings indicated that this Se ENM-enabled nanotechnology was beneficial to human health and shows promising sustainable application in crop production. |
1. Introduction
Selenium (Se) contributes to the synthesis of selenoproteins for improving the human antioxidant activity and immune system.1 To date, 25 selenoproteins have been identified, and their nutritional functions are achieved by the active center of selenocysteine in mammals.1,2 Se deficiency can lead to a variety of diseases including heart disease, weak immune system, and reproductive defects.3 Therefore, supplying adequate Se is important for human health. The World Health Organization (WHO) has established the lowest limit of acceptable adult daily intake at 50 μg, and the recommended daily allowance (RDA) in China is 55 μg for adults. Human intake of Se mainly comes from the diet, and the Se content in food widely depends on the crop growth conditions.4 The concentration of Se in soil ranges from 0.01 to 2.0 mg kg−1, with an average of ∼0.4 mg kg−1.5 However, the bioavailability of naturally occurring Se is very low for plants, limiting the Se content required for mammals through vegetable consumption. Globally, around 800 million people are deficient in Se.6 Therefore, enhancing the Se content in food is urgently needed for the human health.7At present, application of inorganic Se salts for producing Se-enriched food is an effective way to solve the Se deficiency problem in the diet.6 It was shown that sodium selenate (Na2SeO4, 40 mg L−1) can stimulate wheat growth, boost grain yield, increase Se content (5.99 μg kg−1), and enhance plant resistance under drought stress.8 Sodium selenite (Na2SeO3, 50 mg kg−1) can increase the content of chlorogenic acids (an important active component of anti-oxidation) in Lycium chinense leaves and elevate the level of chlorophyll and carotenoids in photosynthesis systems.9 However, Se inorganic salts as fertilizers are absorbed easily by soil minerals, resulting in low recovery efficiency (∼14%).10 So far, emerging studies reported that zero-valent Se engineered nanomaterials (Se ENMs) with higher mobility and solubility, prominent bioactivity and biosafety showed promising applications in agriculture.11–13 It has been reported that Se ENMs (50–100 mg kg−1) could promote callus organ formation and root growth in tobacco.11 Under low-temperature stress (10 °C), Se ENMs increased the chlorophyll content of tomato leaves by 27.5%, which outperformed Na2SeO4 (19.2%).14 Zahedi et al. reported that foliar spraying of Se ENMs could stimulate the growth and increase the yield of strawberry under salinity stress.15 These reports demonstrated that Se ENMs could promote crop growth and yield, but the key factors for improving Se availability remained to be identified. Moreover, the mechanism of promoting photosynthesis and yield by Se ENMs is unclear.
The rhizosphere microbiome plays a critical role in increasing nutrient availability and promoting plant growth.16–18 For instance, symbiotic associations between plants and nitrogen (N)-fixing bacteria converted atmospheric N into ammonium-N which can be easily taken up by plants.17 Phosphate (P)-solubilizing bacteria excrete organic acids and increase the dissolution of precipitated P, thereby improving the P availability.18 Studies demonstrated that nutrients, such as iron (Fe), magnesium (Mg), and P could improve photosystems and promote photosynthesis,19 and the resulting formation of carbohydrates could provide 11–40% carbon sources for microbes through root exudates.20 Recent research studies found that Ag (100 mg kg−1), SiO2 (100 mg kg−1), TiO2 (100 mg kg−1), and Fe3O4 ENMs (100 mg kg−1) could alter soil microbial community composition and metabolite profiles.21,22 Therefore, we hypothesized that soil application of Se ENMs could increase the abundance of beneficial rhizosphere microorganisms, improve nutrient availability, and increase the Se content in crops. The resulting increased nutrient accumulation in leaves could promote photosynthesis and even yield. The carbohydrates from photosynthesis transported from shoots to roots, which may further promote root growth, improve the rhizosphere microbiome and increase nutrient availability.
Brassica chinensis L. (B. chinensis) was selected as an experimental crop because it is an important vegetable and contains various vitamins for human health.23 The following three aspects were specifically explored: (1) alteration in the rhizosphere microbiome in response to Se ENMs; (2) the mechanisms of Se ENM uptake and Se-enrichment in B. chinensis; (3) the change in photosynthesis and yield of B. chinensis upon Se ENM exposure. This is the first investigation on the relationship among beneficial rhizosphere microorganisms, nutrient availability and photosynthesis of B. chinensis upon Se ENM application, which could provide an alternative model for cultivating Se-enriched crops.
2. Materials and methods
2.1 Synthesis and characterization of Se ENMs
The synthesis of Se ENMs was similar to that in a previous study.24 Briefly, a mixed solution of 4 × 10−5 M selenic acid (H2SeO3, ≥95%, Sinopsin Group Chemical Reagent, Ltd.) and raisin extracts was heated under reflux conditions (pH 5.9). Then, Se ENMs were purified and obtained through centrifugation at 17![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 280g for 20 min. The shape and size of Se ENMs were observed using a transmission electron microscope (TEM, JEM-2100, Nippon Electronics Co., JPN, operating at an acceleration voltage of 200 kV). The hydrodynamic diameter and zeta potential were measured using a Zetasizer Nano (ZEN3600, Malvern, UK), and the details are shown in ESI† Text S1.
280g for 20 min. The shape and size of Se ENMs were observed using a transmission electron microscope (TEM, JEM-2100, Nippon Electronics Co., JPN, operating at an acceleration voltage of 200 kV). The hydrodynamic diameter and zeta potential were measured using a Zetasizer Nano (ZEN3600, Malvern, UK), and the details are shown in ESI† Text S1.
2.2 Plant cultivation and ENM exposure
The soil was collected from Jiangsu Province, China, and the seeds of B. chinensis (no. 9, Suzhouqing) were obtained from the Jiangsu Academy of Agricultural Sciences. The soil was natural and its properties were as follows: a pH of 7.8, a redox potential of 318.5 mV, an electrical conductivity of 0.36 mS cm−1, total nitrogen of 23.6 g kg−1, total phosphorus of 0.38 g kg−1, total potassium of 12.5 g kg−1, total carbon of 61.1 g kg−1, and organic matter of 22.5 g kg−1.Soil was homogenized, by filtering through a 5.0 mm sieve to remove large plant residues and fragments. Then, the soil was blended vigorously with Se ENMs at different concentrations (0, 0.5, and 1 mg kg−1) or with Na2SeO3 (0.5 mg kg−1) to achieve a homogeneous mixture. Each pot was filled with 1 kg prepared soil without aging, and each treatment had five replicates. B. chinensis seeds were sown at a depth of 1 cm in soil. The seedlings were grown in a greenhouse for 60 days under the following conditions: 20/15 °C at day/night and an 18/6 h light/dark cycle. During the growth stage, no additional fertilizers were applied. Detailed soil parameters before seed sowing and after plant harvest are shown in Table S1.† Photosynthesis parameters of B. chinensis leaves were determined before sampling using a CIRAS-3 portable gas exchange system, Hansatech, USA. Root parameters (root length, surface area, average diameter, and number of tips) were analyzed using WinRHIZO Pro 2017b, Canada. The measurements of chlorophyll and carbohydrate content are shown in ESI† Text S2 and S3, respectively. The vitamin C content of B. chinensis leaves were determined by a titration method with 2,6-dichlorophenolindophenol, using 1 g of fresh tissue and HCl (2%).25
2.3 Mineral nutrient analysis in plants
The mineral nutrients and heavy metal (cadmium, Cd) contents in plant tissues using different treatments were analyzed by inductively coupled plasma mass spectrometry (ICP-MS, iCAP-TQ, Thermo-Fisher, USA).26,27 Briefly, dried tissues (25 mg) were digested (190 °C, 30 min) in a microwave accelerated reaction system (CEM Corp., Matthews, NC) with a mixture of HNO3 (GR, 65–68%) and ultrapure water (v/v = 4:3). The digested solution was filtered with a microporous membrane (0.22 μm) and diluted to 50 mL by using ultrapure water (Milli-Q) for ICP-MS analysis. For quality control (QC) and quality assurance (QA), the standard reference material (GBW 07602, bush twigs and leaves purchased from Nanjing Alida Biotechnology Co., Ltd., China) was digested and measured using the same procedures. The recoveries of all elements were between 84.9 and 102.9%.
2.4 Soil microbial community composition analysis
To characterize the soil microbial diversity and composition, high-throughput sequencing of soil bacterial 16S rRNA genes was performed on an Illumina MiSeq platform (Personal Biotechnology Co., Ltd. Shanghai, China). Briefly, the total DNA of soil microorganisms was extracted by using the Fast DNA SPIN extraction kits (MP Biomedicals, Santa Ana, CA, USA) according to the manufacturer's protocols. The concentration and quality of the total DNA were measured using a Nanodrop (Thermo Scientific, NC2000, USA) and 1.2% agarose gel electrophoresis (Invitrogen, AM9870, USA). The V3–V4 region of the microbial 16S rRNA gene was amplified with primers 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) using a quantitative PCR system (ABI, 2729, USA). The amplification system includes 5× reaction buffer (5 μL), 5× GC buffer (5 μL), dNTP (2.5 mM; 2 μL), a forward primer (10 μM; 1 μL), a reverse primer (10 μM; 1 μL), a DNA template (2 μL), ddH2O (8.75 μL), and Q5 DNA polymerase (0.25 μL). The quantitative real-time polymerase chain reaction (qRT-PCR) program parameters were 98 °C 2 min, denaturation 98 °C 15 s, annealing 55 °C 30 s, extension 72 °C 30 s, final extension 72 °C 5 min, 10 °C hold. 25–30 cycles. After the amplification, Illumina's TruSeq Nano DNA LT Library Prep Kit was used to prepare a sequencing library, and this library was selected and purified by 2% agarose gel electrophoresis. After completion of the amplification step, amplicons were pooled in equal amounts, and pair-end 2 × 300 bp sequencing was performed using the Illumina MiSeq platform with a MiSeq Reagent Kit v3 at Shanghai Personal Biotechnology Co., Ltd. (Shanghai, China).2.5 Quantitative real-time PCR (qRT-PCR) analysis
The qRT-PCR was used to investigate the regulation of key genes involved in Se translocation and carbohydrate transport. The total RNA of B. chinensis leaves (60 days) was extracted using a TaKaRa MiniBEST Plant RNA Extraction Kit according to the protocol of the manufacturer. The concentration of RNA was measured using an ultramicro-spectrophotometer (UltraM-QB200, Gallop tech Co., Ltd., Shanghai, China). cDNA was prepared from RNase-treated total RNA using a cDNA synthesis kit (CW Biotech Co., Ltd. Jiangsu, China) according to the manufacturer's protocols. Each PCR reaction system (50 μL final volume) contained a 2 μL cDNA template, a 1 μL forward primer (10 μM), a 1 μL reverse primer (10 μM) (Table S2†), and a 25 μL 2× Ultra SYBR mixture. The qRT-PCR was performed using a CFX96™ real-time system (BIO-RAD, USA). The reaction program was 10 min at 95 °C, followed by 40 cycles at 95 °C for 15 s and 60 °C for 1 min. Three replicates were done for each biological sample. The relative gene expression was calculated using the 2−ΔΔCT method.2.6 Statistical analysis
Biomass, photosynthesis, and root parameters were presented as the mean ± standard deviation. One-way ANOVA with the Fisher LSD test was used to analyze the variation among different treatments using Origin Statistics 2019b at p < 0.05. Sequence data analyses were mainly performed using QIIME and R packages (v3.5.0). All treatments were conducted at least three times.3. Results and discussion
3.1 Characterization and promoted crop growth of Se ENMs
The as-prepared Se ENMs showed a size distribution (Fig. 1a) of 20–90 nm with an average size of 62.3 ± 14.6 nm (Fig. 1b). The hydrodynamic diameter and zeta potential of the Se ENMs in ultrapure water were 648.9 ± 24.2 nm and −34.4 ± 1.4 mV (Table S3†), respectively. Previous studies demonstrated that the size and surface charge had a profound impact on the root uptake and translocation of ENMs.27–31 Although there are still some inconsistent results about the size, it is certain that nanometer-sized materials can be taken by plants.32 A previous study also demonstrated that negatively charged CeO2 ENMs were more efficiently translocated than the positively charged ones in roots due to electrostatic repulsion.30 Therefore, the nano-sized and negatively charged Se ENMs may have potential applications in agricultural production. The growth-promoting effect of Se ENMs after soil application was observed in B. chinensis (Fig. S1† and 1c). Both fresh and dry weights (FW and DW) of B. chinensis were increased (19.8% and 24.8%) upon exposure to 0.5 mg kg−1 Se ENMs as compared to the control (CK) (Fig. 1d and e). Particularly, the yield (FW, defined in ESI† Text S4) of B. chinensis increased by 19.8% upon 0.5 mg kg−1 Se ENM exposure (Fig. 1d). Although no significant difference in the FW of shoots was observed between the 1 mg kg−1 and 0.5 mg kg−1 treatments, the DW of shoots differed (Fig. 1d and e). Moreover, the Se content in roots and shoots of B. chinensis increased by 257.1% and 338%, respectively (Fig. 1f). Additionally, the vitamin C content in B. chinensis increased by 118.2% (Fig. S2†). Many other mineral nutrients had no statistical difference, while a significant decrease of the Cd content (23.41%) in edible parts was observed upon Se ENM (0.5 mg kg−1) exposure (Fig. S3†). The dissolution experiment demonstrated that the Se ENMs showed stability in soil because only 0.9% of Se could be released from the Se ENMs (ESI† Text S5 and Fig. S4). Moreover, the plant promotion effects of Se ENMs (0.5 mg kg−1) were better than those of Na2SeO3 (0.5 mg kg−1) (Fig. S5†). Therefore, we argued that the Se ENMs could be taken up by plants in its nanoparticle form, consistent with some previous research studies.28,33,34 For example, Wang et al. reported that CuO ENMs were taken up by maize and transported from roots to shoots.28 CeO2 ENMs were detected in roots, nodules, stems, leaves, and pods of soybeans.33 In addition, previous studies showed that Se could be taken up by roots and transported to shoots, then accumulated in plant leaves and be metabolized in chloroplasts.35–37 Therefore, most of the increased Se content may accumulate in the chloroplasts. It has been noted that the sublethal toxic concentrations of Se in food were 1–30 μg g−1 dry weight,38 and the Se content was only up to 0.33 μg g−1 in this study. Therefore, the Se ENMs (0.5 mg kg−1) applied in B. chinensis production in the soil studied were safe and beneficial. These results suggested that the as-prepared negatively charged Se ENMs have a promising potential for producing Se-enriched crops. Moreover, given that the bioavailability of the Se ENMs at 1 mg kg−1 was not superior to that of the Se ENMs at 0.5 mg kg−1, further experiments were focused on the effects of the Se ENMs at 0.5 mg kg−1. | ||
| Fig. 1 Characterization and promoted crop growth of the Se ENMs. (a) TEM image of the Se ENMs; (b) size distribution of the Se ENMs; (c) photos of the harvested B. chinensis exposed to the Se ENMs at 0, 0.5, and 1.0 mg kg−1, respectively; (d) shoot FW and (e) shoot DW of B. chinensis; (f) Se content in B. chinensis (n = 3, p < 0.05). | ||
3.2 Alteration in the rhizosphere microbiome of B. chinensis in response to the Se ENMs
The rhizosphere microbiome could improve the bioavailability of ENMs through mediating the dissolution and anti-aggregation39 and also absorb metal ions by electrostatic attraction.40 In this study, a change of rhizosphere microbial community composition was observed after Se ENM (0.5 mg kg−1) exposure. A total of 1012536 quality sequences of the six soil samples were obtained from the analysis of rhizosphere bacterial community composition. The read lengths varied from 50 to 442 bp with an average of 382 bp at the 97% similarity level. No significant difference was observed in the Shannon (H) and Simpson indexes of Se ENM and CK exposed treatments (Fig. 2a and S6,†p < 0.05), suggesting that the Se ENMs at 0.5 mg kg−1 did not alter the rhizosphere microbial diversity. Meanwhile, the beta-diversity of rhizosphere bacteria was changed distinctly according to the principal coordinates analysis (PCoA) and non-metric multidimensional scaling (NMDS) based on the weighted UniFrac distance (Fig. 2b and S7†). The PCoA showed 78.2% and 8.6% of the total variation for the bacterial data, demonstrating that the bacterial species of CK was obviously different from that of the Se ENM-treated soil. Meanwhile, the stress value of NMDs smaller than 0.2 further demonstrated the significant composition difference between CK and Se ENM-exposed groups. These results suggested that the Se ENMs (0.5 mg kg−1) significantly affected the rhizosphere bacterial community composition. Other research indicated that metal and metal oxide ENMs in soil could alter rhizosphere microbial community compositions.21,22 He et al. reported that FeO ENMs (1 mg kg−1) could promote the microbial growth via providing essential nutrients (C and N).41 However, another study demonstrated that CuO ENMs could reduce the microbial activities.42 Therefore, the interaction between ENMs and rhizosphere microbiome warrants further detailed research.
 | ||
| Fig. 2 Alteration in the rhizosphere microbiome of B. chinensis in response to Se ENMs. (a) Alpha diversity index (Simpson); (b) principal coordinates analysis (PCoA); (c) relative abundances of phylum; (d) heat map clustering of microbial species (n = 3, p < 0.05). | ||
In our studies with Se ENMs at the phylum level, Proteobacteria, Bacteroidetes, Actinobacteria, Chloroflexi, Acidobacteria, Gemmatimonadetes, Deinococcus–Thermus, Patescibacteria, Firmicutes, and Verrucomicrobia were the dominant phyla in both CK and Se ENM (0.5 mg kg−1) treated rhizosphere soils (Fig. 2c). Notably, the Se ENMs increased the abundance of Proteobacteria (3.3%), Bacteroidetes (3.3%), and Deinococcus–Thermus (1.7%). It has been demonstrated that Proteobacteria can act as plant growth-promoting rhizobacteria for improving plant growth.43Bacteroidetes is known to degrade high molecular weight compounds including proteins and polysaccharides, which might positively affect the sugar and organic acid metabolism in soil.21 It had been reported that the amount of Deinococcus–Thermus increased under hydrocarbon contamination44 and had the ability of absorbing heavy metals, such as lead (Pb), and Cd.40 This could be the reason for the reduction in the Cd content in the harvested B. chinensis in this study (Fig. S4d†).
At the genus, the relative abundance of Truepera, Chryseolinea and Xanthomarina increased by 1.6%, 0.3% and 0.7%, respectively (Fig. S8†). Chryseolinea is responsible for the oxidization and utilization of diverse mono- and disaccharides as well as polysaccharides and organic acids.45 The increased abundance of Chryseolinea might enhance the recycling of carbon in the rhizosphere. Xanthomarina is involved in phosphate solubilization and indole-3-acetic acid and siderophore production.46Truepera affiliated to the Deinococcus–Thermus phylum, which has potential to absorb heavy metals,40 may reduce the adverse effect of heavy metals on plant growth.
Furthermore, the species composition heat map (Fig. 2d) indicates that the Se ENMs (0.5 mg kg−1) changed the rhizosphere microbiome, compared to CK. This result confirmed the change of rhizosphere bacterial composition, which was identified with the PCoA and NMDS (Fig. 2b and S7†). The Se ENMs significantly increased the abundance of rhizosphere PGPR (Fig. S9†), such as Cellvibrio, Pseudomonas, Bacillus, and Micromonospora. The abundance of Pseudomonas and Bacillus increased upon Se ENM exposure, implying increased colonization in the rhizosphere which could stimulate plant growth. It is reported that Pseudomonas had the potential for enhancing the growth of lettuce,47 and the root length of tomato seedlings can be significantly improved in the presence of Pseudomonas.48 Chowdhury et al.49 reported that Bacillus was able to effectively reduce the disease severity (DS) of bottom rot on lettuce. A similar study found that Bacillus obviously promoted the height of peppers by 27.2–54.5% under the stress of Fusarium wilt.50
3.3 Increased uptake and accumulation of Se in B. chinensis
As discussed above, the abundance of beneficial rhizosphere bacteria (Pseudomonas and Bacillus) increased in B. chinensis upon Se ENM exposure. Rhizosphere microorganisms could produce low molecular weight compounds (e.g., oxalic acid, malic acid, and citric acid),51 which can increase the bioavailability of soil nutrients.18 Sura-de Jong et al. found that Pseudomonas promoted the growth of Se hyper-accumulators (Brassicaceae, Fabaceae).52Bacillus can produce antimicrobial metabolites to inhibit phytopathogens and promote plant growth and Se bio-fortification in plants.53 Durán et al.54 demonstrated that Bacillus enhanced the Se content in wheat grain after co-inoculation of selenobacteria and arbuscular mycorrhizal fungi. In this study, after exposure to Se ENMs (0.5 mg kg−1), the dissolved organic carbon (DOC) content in the rhizosphere increased by 29.2% (Fig. 3a) and the rhizosphere soil pH decreased from 7.8 to 7.3 (Fig. S10†). Fig. 3b and S11† demonstrates that the contents of low molecular weight compounds (betaine, proline, glycine, norleucine, urocanic acid and indole-3-acrylic acid) increased significantly in the rhizosphere of B. chinensis (the details for low molecular weight compound analysis are described in ESI† Text S6, Table S4 and Fig. S12), which could improve the availabilities of nutrients via promoting their solubility in soil.17,18,39,55 In addition, low-molecular weight organic acids could reduce homo-aggregation and/or hetero-aggregation of ENMs in the rhizosphere,56,57 resulting in an increase in their bioavailability. Our results confirmed that the uptake efficiency (taking the Se amount per unit length of the root) of Se increased by 264.9% using the 0.5 mg kg−1 Se ENMs compared to that using the control (Fig. 3c). Previous studies have shown that Se could be taken up and translocated by a high-affinity sulfate transporter due to a similarity in the chemical properties of Se and sulfate.4 In our study, the gene expression of a high-affinity sulfur transporter (BnSULTR1,1) was up-regulated by 149.0% (Fig. 3d), accompanied by a 338.0% increase in the Se content in B. chinensis (Fig. 3e). Therefore, Se ENMs could increase the abundance of beneficial rhizosphere microorganisms and the total organic carbon and improve the Se-uptake efficiency, contributing to the enrichment of Se in B. chinensis (Fig. 3f).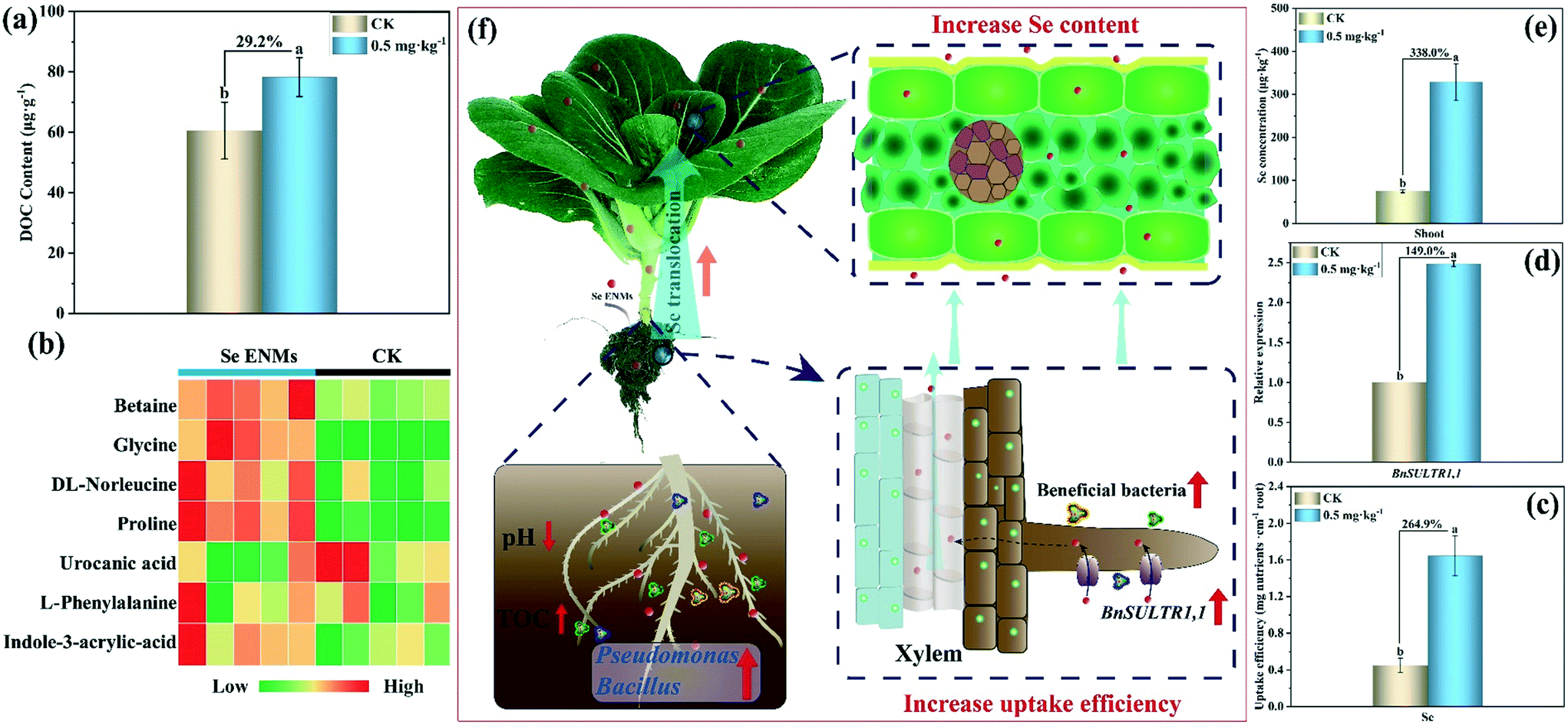 | ||
| Fig. 3 The mechanism of the uptake and enrichment of the Se ENMs. (a) The DOC content in the rhizosphere; (b) low molecular weight compounds increased significantly in the rhizosphere soil after exposure to the Se ENMs (0.5 mg kg−1) (n = 5, p < 0.05); the uptake efficiency (c), transporter activity (d) and Se enrichment (e) in B. chinensis; (f) schematic diagram of the uptake and enrichment of the Se ENMs in B. chinensis (Fig. 3a and c–e, n = 3, p < 0.05). | ||
Se, an essential element, has great importance in anti-oxidation, detoxification and prevention of diabetes for humans.2,3 The appropriate Se content in edible food is beneficial for human health. The Se content significantly increased to 32.8 μg/100 g in B. chinensis upon Se ENM (0.5 mg kg−1) exposure, while this value was only 7.5 μg/100 g using CK (Fig. 3e). According to China's dietary guidelines, a daily consumption of vegetables should reach 300–500 g which accounts for 24.6–27.2% of the total food intake. Therefore, consuming this Se-enriched B. chinensis could meet the recommended daily Se intake (55–400 μg per day) for humans. Moreover, the Se ENMs may be methylated and integrated into cysteine and methionine in plants,58,59 which were both beneficial for human health. However, it should be noted that high Se intake could cause selenosis and result in some diseases, such as brittle hair and abnormal hair loss, broken nail walls, and motor and sensory abnormalities.60
3.4 Promoted photosynthesis and transport of carbohydrates by the Se ENMs
As stated previously, the Se ENMs in the soil changed the rhizosphere microbiome and increased the DOC content in the rhizosphere soil, indicating that the nutrient availability could be improved. It is reported that nutrients such as Mg, P, and Fe have a key role in enhancing photosystems.19 In this study, these photosynthesis beneficial elements, namely, Mg, P, and Fe in the B. chinensis leaves increased by 24.2%, 22.3%, and 2.5% upon 0.5 mg kg−1 Se ENM exposure compared to those using CK, respectively (Fig. 4a). Meanwhile, their higher utilization efficiencies in roots and shoots were observed (Fig. S13†); thus, these nutrients have a major role in plant growth. Mg and Fe are the components of chlorophyll and are involved in photosynthesis.61,62 P, the component of the chloroplast bilayer membrane, granule and adenosine triphosphate (ATP), plays a significant role in energy conversion.63 After exposure to Se ENMs (0.5 mg kg−1) (Fig. 4a), the contents of chlorophyll a and b, which compose the light harvesting complex,6,64 increased by 16.7% and 18.9%, which may be induced by the increased content of Mg and P in the B. chinensis leaves. Thus, the Se ENMs (0.5 mg kg−1) had huge potential for improving the light harvesting of B. chinensis. As shown in Fig. 4a, the electron transfer rate (ETR) increased by 27.4% in the photosystem. Consequently, photosynthesis would be enhanced, since the increase of Mg, P, and Fe contents by the rhizosphere microbiome possibly resulted in the increase of the chlorophyll content. Studies showed that ENMs can increase the yield through improving photosynthesis.19 For example, CeO2 ENMs (100 mg kg−1) stimulated plant growth by enhancing the photosynthesis rate (54%),65 and nanochitin (6 mg L−1) ENMs can increase the grain yield by 25% through enhancing the photosynthesis rate (13.8%) (winter wheat), respectively.66 Herein, the net photosynthesis rate (Pn) of B. chinensis upon soil application of 0.5 mg kg−1 Se ENMs increased significantly by 16.7% (Fig. 4a), which could contribute to the increase in the yield (Fig. 1d). | ||
| Fig. 4 Effects of the Se ENMs on the B. chinensis growth, photosynthesis and transport of carbohydrates. The mechanisms of (a) enhancing photosynthesis (increasing the beneficial elements and chlorophyll content), (b) improving the transport of carbohydrates in the leaves, and (c) improving the root growth (root length, tip number and surface area) (n = 3, p < 0.05). | ||
Carbohydrate is a product of photosynthesis.67 As shown in Fig. 4b, the carbohydrate content increased by 6.5% upon 0.5 mg kg−1 Se ENM exposure. Sucrose is produced from glucose which serves as the principal product of photosynthesis and could be transported from source leaves to sink organs.68 SUTs/SUCs (sucrose transporters or sucrose carriers) and SWEETs (sugars will eventually be exported transporters) play significant central roles in sucrose translocation and are closely associated with crop yields.68 Herein, three key genes (BnSUC1,1, BnSUC1,4 and BnSWEET10,2) were selected to explore the expression of sucrose translocation genes.68 Their expressions were up-regulated 52.2, 53.2 and 76.3-fold in the B. chinensis leaves, respectively (Fig. S14†), suggesting that the activities of enzymes (SUCs and SWEETs) increased by exposure to Se ENMs. This suggested that increasing the carbohydrates produced more 3C compounds which could be transformed to sucrose and transported in phloem. Therefore, there is a possibility that more sucrose was translocated to the roots for supplying their growth. As shown in Fig. S15,† the roots became larger, and the FW and DW increased by 43.3% and 23.5% upon 0.5 mg kg−1 Se ENM exposure. Meanwhile, the root length, surface area and tip number distinctly increased by 65.0%, 73.1% and 17.7%, respectively (Fig. 4c), indicating that the uptake of water as well as nutrients by the roots might be stimulated by the Se ENMs. Taken together, soil application of the Se ENMs can improve the rhizosphere microbiome to increase the uptake of Mg, P and Fe, followed by an increase in chlorophyll a and b contents and the ETR, ultimately promoting the photosynthesis (Fig. 4a). The translocation of carbohydrates from shoots to roots was also accelerated (Fig. 4b), leading to the promoted root growth (Fig. 4c). The promoted roots could take up more nutrients such as Mg, P, Fe, and Se, which could be translocated to the leaves, further improving photosynthesis. In addition, the transport of carbohydrates from shoots to roots could improve the rhizosphere microbiome. Therefore, the Se ENMs promoted the yield and Se content of B. chinensis through increasing the abundance of rhizosphere beneficial microorganisms, promoting the nutrient uptake efficiency and enhancing photosynthesis. Moreover, the Se ENMs increased the microelements and beneficial components and decreased the heavy metal content in B. chinensis. This research provides a promising way for producing Se-enriched vegetables.
4. Conclusions
Herein, a novel Se ENM-enabled agricultural nanotechnology was developed to produce Se-enriched vegetables. Se ENMs (0.5 mg kg−1) in soil can increase the net photosynthesis rate (16.7%), carbohydrate content (6.5%), yield (19.8%), and Se content up to 32.8 μg/100 g. Moreover, the mechanism of uptake and enrichment of Se in B. chinensis with soil application of Se ENMs was investigated in detail. We found that the Se ENMs could boost photosynthesis efficiency, trigger the transport of carbohydrates from shoots to roots, and improve the beneficial rhizosphere microbiome. The rhizosphere microbiome (Pseudomonas and Bacillus) together with low molecular weight compounds (betaine, proline, glycine, norleucine, urocanic acid and indole-3-acrylic acid) enhanced the uptake efficiency of Se by 264.9%. The produced vegetables can meet the recommended daily Se intake (55–400 μg per day) for humans. Therefore, this study could provide an alternative ENM-enabled nanotechnology for producing Se-fortified crops, which is beneficial for human health and shows promising application in crop production.Conflicts of interest
The authors declare no competing financial interest.Acknowledgements
This research was supported by the National Natural Science Foundation of China (41820104009, 42077296, 41807378 and 41907304), the Agricultural Science and Technology Innovation Project of Jiangsu Province (CX(20)3080), the UMass Conti Faculty Fellowship, and the USDA Hatch program (MAS 00549).References
- M. P. Rayman, Selenium and human health, Lancet, 2012, 379, 1256–1268 CrossRef CAS.
- G. V. Kryukov, S. Castellano, S. V. Novoselov, A. V. Lobanov, O. Zehtab, R. Guigó and V. N. Gladyshev, Characterization of mammalian selenoproteomes, Science, 2003, 300, 1439–1443 CrossRef CAS PubMed.
- D. L. Hatfield, P. A. Tsuji, B. A. Carlson and V. N. Gladyshev, Selenium and selenocysteine: roles in cancer, health, and development, Trends Biochem. Sci., 2014, 39, 112–120 CrossRef CAS PubMed.
- Y. Zhu, E. A. H. Pilon-Smits, F. Zhao, P. N. Williams and A. A. Meharg, Selenium in higher plants: understanding mechanisms for biofortification and phytoremediation, Trends Plant Sci., 2009, 14, 436–442 CrossRef CAS.
- E. Marguí, G. H. Floor, M. Hidalgo, P. Kregsamer, G. Román-Ross, C. Streli and I. Queralt, Analytical possibilities of total reflection X-ray spectrometry (TXRF) for trace selenium determination in soils, Anal. Chem., 2010, 82, 7744–7751 CrossRef PubMed.
- M. Malagoli, M. Schiavon, S. Dall'Acqua and E. A. H. Pilon-Smits, Effects of selenium biofortification on crop nutritional quality, Front. Plant Sci., 2015, 6, 280 Search PubMed.
- K. H. Winther, M. P. Rayman, S. J. Bonnema and L. Hegedüs, Selenium in thyroid disorders-essential knowledge for clinicians, Nat. Rev. Endocrinol., 2020, 16, 165–176 CrossRef CAS.
- F. Nawaz, R. Ahmad, M. Y. Ashraf, E. A. Waraich and S. Z. Khan, Effect of selenium foliar spray on physiological and biochemical processes and chemical constituents of wheat under drought stress, Ecotoxicol. Environ. Saf., 2015, 113, 191–200 CrossRef CAS.
- J. Dong, Y. Wang, S. Wang, L. Yin, G. Xu, C. Lei and M. Zhang, Selenium increases chlorogenic acid, chlorophyll and carotenoids of Lyciumchinense leaves, J. Sci. Food Agric., 2013, 93, 310–315 CrossRef CAS PubMed.
- G. Lyons, Selenium in cereals: improving the efficiency of agronomic biofortification in the UK, Plant Soil, 2010, 332, 1–4 CrossRef CAS.
- E. Domokos-Szabolcsy, L. Marton, A. Sztrik, B. Babka, J. Prokisch and M. Fari, Accumulation of red elemental selenium nanoparticles and their biological effects in Nicotiniatabacum, Plant Growth Regul., 2012, 68, 525–531 CrossRef CAS.
- E. Domokos-Szabolcsy, N. Alladalla, T. Alshaal, A. Sztrik, L. Márton and H. El-Ramady, In vitro comparative study of two Arundo donax L. ecotypes' selenium tolerance, Int. J. Hortic. Sci., 2014, 20, 119–122 Search PubMed.
- H. El-Ramady, N. Abdalla, H. S. Taha, T. Alshaal, A. El-Henawy, E.-D. A. F. Salah, M. S. Shams, S. M. Youssef, T. Shalaby, Y. Bayoumi, N. Elhawat, S. Shehata, A. Sztrik, J. Prokisch, M. Fári, E. Domokos-Szabolcsy, E. A. Pilon-Smits, D. Selmar, S. Haneklaus and E. Schnug, Selenium and nano-selenium in plant nutrition, Environ. Chem. Lett., 2016, 14, 123–147 CrossRef CAS.
- M. Haghighi, R. Abolghasemi and D. S. J. A. Teixeira, Low and high temperature stress affect the growth characteristics of tomato in hydroponic culture with Se and nano-Se amendment, Sci. Hortic., 2014, 178, 231–240 CrossRef CAS.
- S. M. Zahedi, M. Abdelrahman, M. S. Hosseini, N. F. Hoveizeh and L. F. Tran, Alleviation of the effect of salinity on growth and yield of strawberry by foliar spray of selenium-nanoparticles, Environ. Pollut., 2019, 253, 246–258 CrossRef CAS.
- R. L. Berendsen, C. M. J. Pieterse and P. A. H. M. Bakker, The rhizosphere microbiome and plant health, Trends Plant Sci., 2012, 17, 478–486 CrossRef CAS.
- H. P. Bais, T. L. Weir, L. G. Perry, S. Gilroy and J. M. Vivanco, The role of root exudates in rhizosphere interactions with plants and other organisms, Annu. Rev. Plant Biol., 2006, 57, 233–266 CrossRef CAS.
- Y. Liu, X. Li and Y. Kou, Ectomycorrhizal Fungi: Participation in Nutrient Turnover and Community Assembly Pattern in Forest Ecosystems, Forests, 2020, 11, 453 CrossRef.
- Y. L. Liu, L. Yue, Z. Y. Wang and B. S. Xing, Processes and mechanisms of photosynthesis augmented by engineered nanomaterials, Environ. Chem., 2019, 16, 430–445 CrossRef.
- D. V. Badri and J. M. Vivanco, Regulation and function of root exudates, Plant, Cell Environ., 2009, 32, 666–681 CrossRef.
- H. Zhang, M. Huang, W. Zhang, J. L. Gardea-Torresdey, J. C. White, R. Ji and L. J. Zhao, Silver nanoparticles alter soil microbial community compositions and metabolite profiles in unplanted and cucumber-planted soils, Environ. Sci. Technol., 2020, 54, 3334–3342 CrossRef PubMed.
- W. Zhang, X. Jia, S. Chen, J. Wang, R. Ji and L. J. Zhao, Response of soil microbial communities to engineered nanomaterials in presence of maize (Zea mays L.) plants, Environ. Pollut., 2020, 267, 115608 CrossRef PubMed.
- H. An, X. Qi, M. L. Gaynor, Y. Hao, S. C. Gebken, M. E. Mabry, A. C. McAlvay, G. R. Teakle, G. C. Conant, M. S. Barker, T. Fu, B. Yi and J. C. Pires, Transcriptome and organellar sequencing highlights the complex origin and diversification of allotetraploid Brassica napus, Nat. Commun., 2019, 10, 2878 CrossRef.
- G. Sharma, A. R. Sharma, R. Bhavesh, J. Park, B. Ganbold, J. Nam and S. Lee, Biomolecule-mediated synthesis of selenium nanoparticles using dried Vitis vinifera (Raisin) extract, Molecules, 2014, 19, 2761–2770 CrossRef.
- H. Zhang, X. Si, X. Ji, R. Fan, J. Liu, K. Chen, D. Wang and C. Gao, Genome editing of upstream open reading frames enables translational control in plants, Nat. Biotechnol., 2018, 36, 894–898 CrossRef PubMed.
- Y. K. Rui, Q. Q. Yu, Y. H. Jin, J. Guo and Y. B. Luo, Application of ICP-MS to the detection of forty elements in wine, Guangpuxue Yu Guangpu Fenxi, 2007, 27, 1015–1017 CAS.
- Y. K. Rui, J. B. Shen and F. S. Zhang, Application of ICP-MS to determination of heavy metal content of heavy metals in two kinds of N fertilizer, Guangpuxue Yu Guangpu Fenxi, 2008, 28, 2425–2427 CAS.
- Z. Wang, X. Xie, J. Zhao, X. Liu, W. Feng, J. C. White and B. Xing, Xylem- and phloem-based transport of CuO nanoparticles in maize (Zea mays L.), Environ. Sci. Technol., 2012, 46, 4434–4441 CrossRef CAS PubMed.
- X. Ma, J. Geiser-Lee, Y. Deng and A. Kolmakov, Interactions between engineered nanoparticles (ENPs) and plants: phytotoxicity, uptake and accumulation, Sci. Total Environ., 2010, 408, 3053–3061 CrossRef CAS.
- J. Li, R. V. Tappero, A. S. Acerbo, H. Yan, Y. Chu, G. V. Lowry and J. M. Unrine, Effect of CeO2 nanomaterial surface functional groups on tissue and subcellular distribution of Ce in tomato (Solanum lycopersicum), Environ. Sci.: Nano, 2019, 6, 273–285 RSC.
- A. Avellan, J. Yun, Y. Zhang, E. Spielman-Sun, J. M. Unrine, J. Thieme, J. Li, E. Lombi, G. Bland and G. V. Lowry, Nanoparticle size and coating chemistry control foliar uptake pathways, translocation, and leaf-to-rhizosphere transport in wheat, ACS Nano, 2019, 13, 5291–5305 CrossRef CAS.
- Y. Su, V. Ashworth, C. Kim, A. S. Adeleye, P. Rolshausen, C. Roper, W. Jason and D. Jassby, Delivery, uptake, fate, and transport of engineered nanoparticles in plants: a critical review and data analysis, Environ. Sci.: Nano, 2019, 6, 2311–2331 RSC.
- J. A. Hernandez-Viezcas, H. Castillo-Michel, J. C. Andrews, M. Cotte, C. Rico, J. R. Peralta-Videa, Y. Ge, J. H. Priester, P. A. Holden and J. L. Gardea-Torresdey, In Situ Synchrotron X-ray Fluorescence Mapping and Speciation of CeO2 and ZnO Nanoparticles in Soil Cultivated Soybean (Glycine max), ACS Nano, 2013, 7, 1415–1423 CrossRef CAS.
- P. Miralles, T. L. Church and A. T. Harris, Toxicity, Uptake, and Translocation of Engineered Nanomaterials in Vascular plants, Environ. Sci. Technol., 2012, 46, 9224–9239 CrossRef CAS.
- T. G. Sors, D. R. Ellis and D. E. Salt, Selenium uptake, translocation, assimilation and metabolic fate in plants, Photosynth. Res., 2005, 86, 373–389 CrossRef CAS PubMed.
- H. El-Ramady, N. Abdalla, T. Alshaal, A. El-Henawy, E. D. F. Salah, M. S. Shams, T. Shalaby, Y. Bayoumi, N. Elhawat, S. Shehata, A. Sztrik, J. Prokisch, M. Fári, E. A. Pilon-Smits and É. Domokos-Szabolcsy, Selenium and its role in higher plants, Pollutants in Buildings, Water and Living Organisms, 2015, pp. 235–296 Search PubMed.
- M. Gupta and S. Gupta, An overview of selenium uptake, metabolism, and toxicity in plants, Front. Plant Sci., 2017, 7, 2074 Search PubMed.
- A. M. Debruyn and P. M. Chapman, Selenium toxicity to invertebrates: will proposed thresholds for toxicity to fish and birds also protect their prey?, Environ. Sci. Technol., 2007, 41, 1766–1770 CrossRef CAS PubMed.
- L. E. Barton, A. N. Quicksall and P. A. Maurice, Siderophore-mediated dissolution of hematite (α-Fe2O3): effects of nanoparticle size, Geomicrobiol. J., 2012, 29, 314–322 CrossRef CAS.
- B. R. Ginn and J. B. Fein, The effect of species diversity on metal adsorption onto bacteria, Geochim. Cosmochim. Acta, 2008, 72, 3939–3948 CrossRef.
- S. He, Y. Feng, J. Ni, Y. Sun, L. Xue, Y. Feng, Y. Yu, X. Lin and L. Yang, Different responses of soil microbial metabolic activity to silver and iron oxide nanoparticles, Chemosphere, 2016, 147, 195–202 CrossRef PubMed.
- M. Simonin, A. A. Cantarel, A. Crouzet, J. Gervaix, J. M. Martins and A. Richaume, Negative effects of copper oxide nanoparticles on carbon and nitrogen cycle micro-bial activities in contrasting agricultural soils and in presence of plants, Front. Microbiol., 2018, 9, 3102 CrossRef PubMed.
- M. Bruto, C. Prigent-Combaret, D. Muller and Y. Moënne-Loccoz, Analysis of genes contributing to plant-beneficial functions in plant growth-promoting rhizobacteria and related Proteobacteria, Sci. Rep., 2014, 4, 6261 CrossRef PubMed.
- D. J. Saul, J. M. Aislabie, C. E. Brown, L. Harris and J. M. Foght, Hydrocarbon contamination changes the bacterial diversity of soil from around Scott Base, Antarctica, FEMS Microbiol. Ecol., 2005, 53, 141–155 CrossRef PubMed.
- J.-J. Kim, M. Alkawally, A. L. Brady, W. I. C. Rijpstra, J. S. S. Damsté and P. F. Dunfield, Chryseolinea serpens gen. nov., sp. nov., a member of the phylum Bacteroidetes isolated from soil, Int. J. Syst. Evol. Microbiol., 2013, 63, 654–660 CrossRef PubMed.
- J. Crovadore, B. Cochard, G. Calmin, R. Chablais, T. Schulz and F. Lefort, Whole-genome sequence of Pseudomonas xanthomarina strain UASWS0955, a potential biological agent for agricultural and environmental uses, Genome Announc., 2016, 4, e01136-16 Search PubMed.
- M. A. Cipriano, M. Lupatini, L. Lopes-Santos, M. J. Da Silva, L. F. Roesch, S. A. Destéfano, S. S. Freitas and E. E. Kuramae, Lettuce and rhizosphere microbiome responses to growth promoting Pseudomonas species under field conditions, FEMS Microbiol. Ecol., 2016, 92, fiw197 CrossRef PubMed.
- R. Daniel and S. Gustavo, Evaluation of Bacillus and Pseudomonas to colonize the rhizosphere and their effect on growth promotion in tomato (Physalis ixocarpa Brot. ex Horm.), Rev. Chapingo Ser. Hortic., 2016, 22, 45–58 Search PubMed.
- S. P. Chowdhury, K. Dietel, M. Rändler, M. Schmid, H. Junge, R. Borriss, A. Hartmann and R. Grosch, Effects of Bacillus amyloliquefaciens FZB42 on lettuce growth and health under pathogen pressure and its impact on the rhizosphere bacterial community, PLoS One, 2013, 8, e68818 CrossRef.
- X. Yu, C. Ai, L. Xin and G. Zhou, The siderophore-producing bacterium, Bacillus subtilis CAS15, has a biocontrol effect on Fusarium wilt and promotes the growth of pepper, Eur. J. Soil Biol., 2011, 47, 138–145 CrossRef.
- N. Zhang, D. Wang, Y. Liu, S. Li, Q. Shen and R. Zhang, Effects of different plant root exudates and their organic acid components on chemotaxis, biofilm formation and colonization by beneficial rhizosphere-associated bacterial strains, Plant Soil, 2014, 374, 689–700 CrossRef CAS.
- M. Sura-de Jong, R. J. B. Reynolds, K. Richterova, L. Musilova, L. C. Staicu, I. Chocholata, J. J. Cappa, S. Taghavi, D. Van der Lelie, T. Frantik, I. Dolinova, M. Strejcek, A. T. Cochran, P. Lovecka and E. A. Pilon-Smits, Selenium hyperaccumulators harbor a diverse endophytic bacterial community characterized by high selenium resistance and plant growth promoting properties, Front. Plant Sci., 2015, 6, 113 Search PubMed.
- Ö. Baysal, D. Lai, H. Xu, M. Siragusa, M. Çalışkan, F. Carimi, J. A. T. Da Silva and M. A. Tör, Proteomic Approach Provides New Insights into the Control of Soil-Borne Plant Pathogens by Bacillus Species, PLoS One, 2013, 8, e53182 CrossRef PubMed.
- P. Durán, J. J. Acuña, M. A. Jorquera, R. Azcón, F. Borie, P. Cornejo and M. L. Mora, Enhanced selenium content in wheat grain by co-inoculation of selenobacteria and arbuscular mycorrhizal fungi: A preliminary study as a potential Se biofortification strategy, J. Cereal Sci., 2013, 57, 275–280 CrossRef.
- J. Xiong, J. Li, H. Wang, C. Zhang and M. S. Naeem, Fullerol improves seed germination, biomass accumulation, photosynthesis and antioxidant system in Brassica napus L. under water stress, Plant Physiol. Biochem., 2018, 129, 130–140 CrossRef.
- C. Layet, M. Auffan, C. Santaella, C. Chevassus-Rosset, M. Montes, P. Ortet, M. Barakat, B. Collin, S. Legros, M. N. Bravin, B. Angeletti, I. Kieffer, O. Proux, J. L. Hazemann and E. Doelsch, Evidence that soil properties and organic coating drive the phytoavailability of cerium oxide nanoparticles, Environ. Sci. Technol., 2017, 51, 9756–9764 CrossRef PubMed.
- H. Shang, H. Guo, C. Ma, C. Li, B. Chefetz, T. Polubesova and B. Xing, Maize (Zea mays L.) root exudates modify the surface chemistry of CuO nanoparticles: Altered aggregation, dissolution and toxicity, Sci. Total Environ., 2019, 690, 502–510 CrossRef.
- Y. Li, N. Zhu, X. Liang, L. Zheng, C. Zhang, Y. F. Li, Z. Zhang, Y. Gao and J. Zhao, A comparative study on the accumulation, translocation and transformation of selenite, selenate, and SeNPs in a hydroponic-plant system, Ecotoxicol. Environ. Saf., 2020, 189, 109955 CrossRef.
- G. Moreno-Martín, J. Sanz-Landaluze, M. E. León-González and Y. Madrid, Insights into the accumulation and transformation of Ch-SeNPs by Raphanus sativus and Brassica juncea: Effect on essential elements uptake, Sci. Total Environ., 2020, 725, 138453 CrossRef PubMed.
- M. Lemireb, A. Philibert, M. Fillion, C. J. S. Passos, J. R. D. Guimarães, F. B. Jr and D. Mergler, No evidence of selenosis from a selenium-rich diet in the Brazilian Amazon, Environ. Int., 2012, 40, 128–136 CrossRef.
- L. Zhao, L. Lu, A. Wang, H. Zhang, M. Huang, H. Wu, B. Xing, Z. Wang and R. Ji, Nano-biotechnology in agriculture: use of nanomaterials to promote plant growth and stress tolerance, J. Agric. Food Chem., 2020, 68, 1935–1947 CrossRef PubMed.
- M. H. Ghafariyan, M. J. Malakouti, M. R. Dadpour, P. Stroeve and M. Mahmoudi, Effects of magnetite nanoparticles on soybean chlorophyll, Environ. Sci. Technol., 2013, 47, 10645–10652 CAS.
- M. Li, L. Li, X. Shi, L. Lin and S. Lin, Effects of phosphorus deficiency and adenosine 5′-triphosphate (ATP) on growth and cell cycle of the dinoflagellate Prorocentrum donghaiense, Harmful Algae, 2015, 47, 35–41 CrossRef CAS.
- M. Delfani, M. B. Firouzabadi, N. Farrokhi and H. Makarian, Some physiological responses of black-eyed pea to iron and magnesium nanofertilizers, Commun. Soil Sci. Plant Anal., 2014, 45, 530–540 CrossRef CAS.
- Z. Ca, C. Stowers, L. Rossi, W. Zhang, L. Lombardini and X. Ma, Physiological effects of cerium oxide nanoparticles on the photosynthesis and water use efficiency of soybean (Glycine max (L.) Merr.), Environ. Sci.: Nano, 2017, 4, 1086–1094 RSC.
- W. Xue, Y. Han, J. Tan, Y. Wang, G. Wang and H. Wang, Effects of nanochitin on the enhancement of the grain yield and quality of winter wheat, J. Agric. Food Chem., 2018, 66, 6637–6645 CrossRef CAS.
- H. Wang, M. Zhang, Y. Song, H. Li, H. Huang, M. Shao, Y. Liu and Z. Kang, Carbon dots promote the growth and photosynthesis of mung bean sprouts, Carbon, 2018, 136, 94–102 CrossRef CAS.
- H. Jian, K. Lu, B. Yang, T. Wang, L. Zhang, A. Zhang, J. Wang, L. Liu, C. Qu and J. Li, Genome-wide analysis and expression profiling of the SUC and SWEET gene families of sucrose transporters in oilseed rape (Brassica napus L.), Front. Plant Sci., 2016, 7, 1464 Search PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d1en00740h |
| This journal is © The Royal Society of Chemistry 2022 |