
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceNOAH-(15N/13C)-CEST NMR supersequence for dynamics studies of biomolecules†
Rodrigo
Cabrera Allpas
 a,
Alexandar L.
Hansen
b and
Rafael
Brüschweiler
*abc
a,
Alexandar L.
Hansen
b and
Rafael
Brüschweiler
*abc
aDepartment of Chemistry and Biochemistry, The Ohio State University, Columbus, Ohio 43210, USA. E-mail: bruschweiler.1@osu.edu
bCampus Chemical Instrument Center, The Ohio State University, Columbus, Ohio 43210, USA
cDepartment of Biological Chemistry and Pharmacology, The Ohio State University, Columbus, Ohio 43210, USA
First published on 29th July 2022
Abstract
An NMR supersequence is introduced for the rapid acquisition of 15N-CEST and methyl-13C-CEST experiments in the same pulse sequence for applications to proteins. The high sensitivity and accuracy allows the simultaneous quantitative characterization of backbone and side-chain dynamics on the millisecond timescale ideal for routine screening for alternative protein states.
Conformational dynamics in biomolecules can occur on a wide range of timescales from picosecond to milliseconds and beyond and they often play a key role for protein function. Over the years, NMR has proven to be extraordinarily powerful to monitor and quantify the conformational dynamics of biomolecules at atomic detail.1,2 An increasingly popular method is the Chemical Exchange Saturation Transfer (CEST) experiment, in which the spin magnetization is selectively saturated by a relatively long, weak radio-frequency (rf) pulse while conformational exchange occurs, providing detailed information about lowly populated conformational states along with their Boltzmann population, chemical shifts, and kinetic exchange rates kex (typically from 20 to 200 s−1).3,4
For biomolecular applications, CEST is performed as a pseudo-3D NMR experiment where the direct detection F2 dimension is along 1H, the indirect F1 dimension is along the heteronucleus, such as 15N or 13C, and the pseudo-dimension is the frequency offset of the saturation field that is systematically sampled across the spectral range of interest. Hence, a CEST experiment involves the acquisition of many 2D 1H–15N or 1H–13C HSQC spectra, each of them being acquired with a saturation field applied using a variable frequency offset. The number of frequency offsets used along the pseudo-dimension can be rather large, usually in the hundreds, leading to prolonged measurements that can last between a day and a week per experiment depending on the sample concentration and NMR hardware used. Therefore, new approaches that substantially shorten this process are needed.
Recently, a strategy to acquire multiple NMR experiments in sequence has been introduced, known as “NMR by Ordered Acquisition using 1H detection” or NOAH.5 NOAH combines multiple homo- and hetero-nuclear 2D NMR experiments in a single pulse sequence resulting in a “supersequence” where a single recovery delay is placed between sets of scans, thereby significantly reducing the experimental time when compared with the standalone pulse experiments that require a recovery delay after each scan (see also Fig. 1).5–9 So far, NOAH has been mostly applied to small organic molecules in concentrated purified samples combining sets of experiments used for resonance assignments and molecular structure elucidation.6 NOAH has also been extended to complex metabolomics mixture analysis.10,11 For proteins, sequential acquisition of correlation spectra has been reported for resonance assignments.12,13 As of now, the NOAH strategy has not been utilized to benefit protein dynamics experiments.
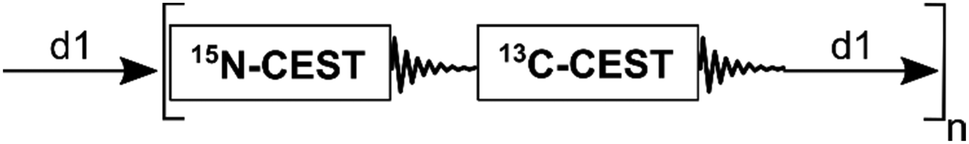 | ||
| Fig. 1 Modular representation of the NOAH-(15N/13C)-CEST supersequence. Delay d1 is the recovery delay and n is the number of times the sequence is repeated. The 15N-CEST module starts with 1H magnetization, while the 13C-CEST module starts with thermal 13C magnetization, which remains largely unperturbed during the first module. The complete pulse sequence is depicted in Fig. S1 (ESI†). | ||
A critical prerequisite for NOAH is the need to preserve magnetization for the immediately succeeding pulse sequence inside the supersequence in order to guarantee good sensitivity.5 Applications of NOAH to small organic molecules have been conducted at 13C natural abundance. For example, using 1.1% of 1H magnetization bound to a 13C nucleus for an HSQC-type experiment and using the remaining 98.9% 1H magnetization for a homonuclear experiment such as TOCSY, as the second experiment, will result in a HSQC/TOCSY supersequence. In addition, specific pulse sequence elements, such as ZIP10 and ASAP,14 are useful to preserve or recover magnetization in NOAH sequences used as starting points in subsequent experiments.6 An advantage of NOAH is that each individual standalone sequence typically requires only minor modifications inside the supersequence resulting in spectra that closely resemble those of the corresponding standalone sequences. Another advantage is that most NMR pulse sequence parameters remain unchanged when a sequence is incorporated into a supersequence, thereby facilitating both testing and application.
Here, we introduce a new supersequence, which was created by concatenating a 15N-CEST and a 13C-CEST for the purpose of studying protein dynamics in biomolecules along the backbone (15N-CEST) and in the methyl groups of amino-acid side-chains (13C-CEST) (Fig. 1 and Fig. S1, ESI†). We refer to this supersequence as NOAH-(15N/13C)-CEST. Results of the standalone 15N-CEST and 13C-CEST experiments are first presented and compared against a supersequence built from these individual sequences for the colicin E7 immunity protein (Im7) and ubiquitin. Since protein NMR samples are prepared in uniformly 15N and 13C-labeled form, some of the NOAH strategies successfully used for small organic molecules at natural abundance do not apply. Here, we start with the 15N-CEST experiment during which 13C magnetization is preserved and used as starting magnetization for the following 13C-CEST during the 2nd half of the supersequence. We show that both 15N-CEST and the 13C-CEST have good sensitivity despite using thermal 13C starting magnetization for the second part of the sequence when compared to a regular standalone 13C-CEST using 1H starting magnetization. The resulting CEST profiles and CEST fitting results are then compared with those of the standalone sequences.
By preserving 13C magnetization during 15N-CEST, the 13C-CEST can immediately follow without a separate recovery delay as implemented in the supersequence scheme of Fig. 1. The 13C magnetization employed is at thermal equilibrium and proportional in size to the gyromagnetic ratio (compared to 1H, γC/γH = 1/4). It therefore does not originate from 1H magnetization by an INEPT-type transfer used in standard 13C-CEST. In the context of NOAH, this has the advantage that the starting 13C magnetization of the 2nd experiment is not affected by depletion of 1H magnetization during the course of the 1st experiment. To assess the sensitivity of this “13C-start” 13C-CEST with the standard 13C-CEST, we measured it both as a standalone experiment and as the 2nd part of the supersequence of Fig. 1. All NMR experiments were performed with a recovery delay d1 of 2 s on an 850 MHz Bruker Ascend magnet equipped with an Avance III HD console and a triple resonance inverse cryoprobe and processed using NMRPipe.15 The standalone direct 13C-CEST and direct 13C-CEST in the supersequence have on average, based on mean cross-peak amplitudes determined by cross-peak fitting for ubiquitin and Im7, a 16% and 11% lower sensitivity than the standard 13C-CEST experiment, respectively. The loss of sensitivity for the 15N-CEST when being part of the supersequence was on average 12% and 7% for ubiquitin and Im7, respectively, compared to the standalone 15N-CEST. For Im7, the 15N-CEST experiment was acquired in 38 hours and the 13C-CEST experiment in 36 hours, for a total of 74 hours for both experiments together. By comparison, the NOAH-(15N/13C)-CEST experiment, including both 15N-CEST and 13C-CEST, was acquired in 45 hours. Therefore, the supersequence produced both experiments, but in 29 hours less time corresponding to 39% saving of spectrometer time. Even if the supersequence is ran longer to have 13C-CEST sensitivity equivalent to that of the standalone sequence, there is still a substantial net gain in experiment time, while the 15N-CEST experiment will have better sensitivity than the reference experiment. The average signal-to-noise ratios along with the experiment times of each sequence are summarized in Table 1.
| Experiment | S/Nc | Individual measurement time (hr) | Combined measurement time (hr) |
|---|---|---|---|
| a Standard 13C-CEST with starting 1H magnetization. b Modified 13C-CEST with starting 13C magnetization and additional 1H decoupling pulses. c Average S/N for all non-overlapping protein cross-peaks. | |||
| 15N-CEST | 205 | 38 | |
| 13C-CESTa | 179 | 36 | 74 |
| 13C-CESTb | 156 | 42 | 80 |
| NOAH 15N-CEST | 191 | — | 45 |
| NOAH 13C-CEST | 160 | ||
The standalone 13C-CEST with starting 13C magnetization has the same sensitivity as the one in the supersequence, which shows that the addition of heat compensation pulses, implemented as decoupling pulse trains applied to 1H, results in an increase in sensitivity when using the 13C magnetization as starting magnetization. This is likely caused by the concurrent heteronuclear Overhauser effect (hetNOE) enhancement via cross-relaxation of 1H to 13C magnetization within the methyl groups during the heat compensation pulses. As can be seen in Table 1, the addition of 1H decoupling pulses to the modified 13C-CEST with 13C starting magnetization leads to a similar sensitivity as the 13C-CEST in the supersequence and it is not far below the signal-to-noise ratio obtained with a regular 13C-CEST with 1H starting magnetization. This enhancement is notable as it opens the possibility of incorporating other methyl-13C-start pulse sequences into new types of supersequences.
After having established that the new supersequence for 15N-CEST and 13C-CEST saves experiment time with only minor sensitivity loss, we tested whether the information obtained by these NOAH-(15N/13C)-CEST profiles is identical to what is obtained by the standalone experiments. Fig. 2 compares CEST profiles of selected residues of Im7 for 15N-CEST with no conformational exchange (top) or two-site exchange (bottom). It demonstrates that the CEST profiles obtained by the two methods are identical. Even small features, such as the shoulder peak of Arg61 (around 121 ppm) on the left of the main dip can be accurately reproduced.
 | ||
| Fig. 2 Representative 15N-CEST profiles for Im7 residues A13, E12, Y55, and R61 from the supersequence (red, solid line) and the standalone sequence (blue, dashed line). Top panels show residues without exchange and the bottom panels show residues that undergo two-site exchange with a 2nd minimum or “shoulder” feature visible in the profile. | ||
The analogous comparison is shown for 13C-CEST in Fig. 3. As for 15N-CEST, the profiles are identical for the standalone experiment vs. supersequence. Small decoupling sidebands can be seen in the bottom panels, which slightly vary since different decoupling sequences were used. Moreover, it can be seen in the bottom panels that the supersequence is able to highly accurately reproduce two-site exchange features manifested as shoulder peaks.
 | ||
| Fig. 3 Representative 13C-CEST profiles for Im7 residues V27Cγ, A28Cβ, V42Cγ, and L3Cδ from the supersequence (red, solid line) and the standalone sequence (blue, dashed line). Top panels show residues without exchange and the bottom panels show residues that undergo two-site exchange with a 2nd minimum or “shoulder” feature clearly visible in the profile. | ||
From the CEST profiles, relaxation and exchange model parameters were extracted using ChemEx (https://www.github.com/gbouvignies/chemex)3 by non-linear least squares fitting and quantitatively compared with the model parameters between the sequences. The parameters extracted are the longitudinal and transverse relaxation rates R1 and R2 of the major state and in the case of two-site exchange also ΔΩ (chemical shift difference between the two states), pA (population of major state) and the exchange rate constant kex = kforward + kreverse. Fig. 4 shows a correlation plot between the fitting results for ΔΩ, R1 and R2 of the major state based on the standalone 15N-CEST and the NOAH 15N-CEST for Im7. Residues with strong overlap were not included in the analysis. Linear regression yields a slope close to 1 with a very small intercept and a high correlation coefficient reflecting the equivalence between the fitting results for the standalone and the supersequence. For the 2-site exchange model applied to the CEST profiles obtained with the standalone 15N-CEST, the fitted pA and kex values were 0.99 and 853 s−1, respectively, while for the NOAH 15N-CEST, these parameters were 0.99 and 757 s−1, respectively. For the standalone 13C-CEST, the fitted pA and kex values were 0.99 and 664 s−1, respectively, while for the NOAH 13C-CEST, these parameters were 0.99 and 644 s−1, respectively. In practice, by measuring CEST profiles at a second B1 field, as is frequently done, robustness of the fitting results should improve and thereby further decrease the difference between the fitted model parameters.
 | ||
| Fig. 4 Correlation plots between standard standalone and the NOAH-(15N/13C)-CEST supersequence for Im7, (A) ΔΩ standalone 15N-CEST vs. ΔΩ NOAH 15N-CEST, (B) ΔΩ standalone 13C-CEST vs. ΔΩ NOAH 13C-CEST, (C) R1 standalone 15N-CEST vs. R1 NOAH 15N-CEST, (D) R1 standalone 13C-CEST vs. R1 NOAH 13C-CEST, (E) R2 standalone 15N-CEST vs. R2 NOAH 15N-CEST, (F) R2 standalone 13C-CEST vs. R2 NOAH 13C-CEST. The linear regression relationships together with Pearson correlation coefficients R2 are displayed in each panel. | ||
In principle, the experimental parameters used to run a NOAH-(15N/13C)-CEST experiment in a 15N and 13C-labeled protein are the same as those when each sequence is executed as a standalone sequence, which facilitates the implementation of the CEST supersequence introduced here. A constraint of this experiment is that both 15N and 13C-CEST supersequence blocks use the same number of rf offsets. Alternatively, it is possible to perform one CEST block with half (or one third) the number of rf offsets compared to the other by repeating for one CEST block the same offset twice (or three times). In practice, however, this limitation should be immaterial as it is outweighed by the substantial time saving allowing one to obtain the entire 13C-CEST experiment essentially for free. Average 13C-T1 relaxation times of Im7 and ubiquitin were 0.62 ± 0.33 s. Larger proteins with longer T1‘s will require a longer recovery delay d1 (Fig. 1), but the relative time saving with respect to standalone CEST experiments will remain.
An attractive application of the NOAH-(15N/13C)-CEST supersequence is the high-throughput screening of protein dynamics, for example, to analyze proteins in the absence vs the presence of ligands (complex) or wild-type vs a mutant protein,16 where it will substantially reduce the NMR time when both backbone and side-chain properties are of interest. Furthermore, it has been shown that using CEST-derived 15N R1 and R2 exchange parameters a lean model-free approach (L-MFA) can be applied for the extraction of S2 order parameters with high accuracy.17 Considering that the supersequence results are equivalent to those of standalone CEST, the supersequence not only provides information about conformational substates exchanging on the ms time scale, but also on the ps–ns timescale reflected in S2 order parameters. This promises to further speed up NMR measurement time for the comprehensive characterization of protein dynamics on time scales from ps to ms for both the protein backbone and side-chains.17
In summary, extended experiment times, which poses a significant bottleneck for pseudo-3D NMR experiments to study biomolecular dynamics of both backbone and side-chain moieties, has been addressed here using the NOAH strategy by combining the 15N-CEST and 13C-CEST into one supersequence. This allows time savings of about 39%, or more than an entire day, when compared to running these sequences in their traditional standalone mode. This new supersequence makes CEST experiments amenable for the screening of protein dynamics under variable experimental conditions, such as temperature, buffer, ligands, or pressure, for the more rapid identification of mutants or substrates of interest involving dynamics changes along the backbone and the side-chains. Furthermore, we showed that the resulting CEST profiles obtained from standalone and NOAH-(15N/13C)-CEST experiments are identical (Fig. 2 and 3), leading to accurate CEST fitting results both in the absence and presence of exchange (Fig. 4). Given the nature of some biomolecules to degrade over time, the application of NOAH for CEST experiments could prove to be a valuable alternative for measuring backbone and side-chain dynamics in significantly less time. Moreover, longer acquisition times can also be more affected by temperature variations, shimming effects, and other fluctuations, which the supersequence could help mitigate. The combination of NOAH-(15N/13C)-CEST with other time-saving techniques, such as multi-frequency excitation,18,19 and non-uniform20 and absolute minimal sampling21 should result in even faster acquisition times. For NOAH experiments of small organic molecules, the computer-assisted generation of supersequences from a library of common NMR experiments has recently been introduced.22 As NOAH experiments start being explored for biomolecular applications, this type of approach may be also feasible for the modular, customized design of new protein NMR experiments.
Rodrigo Cabrera Allpas: developed pulse sequence, analyzed results, wrote original manuscript draft; Alexandar L. Hansen: developed pulse sequence, advised on analysis, edited manuscript; Rafael Brüschweiler: conceived project, wrote manuscript, managed project.
This work was supported by the U.S. National Science Foundation (grant MCB-2103637 to R. B.). All NMR experiments were performed at the Campus Chemical Instrument Center NMR facility at Ohio State University.
Conflicts of interest
There are no conflicts to declare.Notes and references
- T. R. Alderson and L. E. Kay, Cell, 2021, 184, 577–595 CrossRef CAS PubMed.
- A. G. Palmer and H. Koss, in Methods in Enzymology, ed. A. J. Wand, Academic Press, 2019, vol. 615, pp. 177–236 Search PubMed.
- P. Vallurupalli, G. Bouvignies and L. E. Kay, J. Am. Chem. Soc., 2012, 134, 8148–8161 CrossRef CAS PubMed.
- P. Vallurupalli, A. Sekhar, T. Yuwen and L. E. Kay, J. Biomol. NMR, 2017, 67, 243–271 CrossRef PubMed.
- E. Kupče and T. D. W. Claridge, Angew. Chem., Int. Ed., 2017, 56, 11779–11783 CrossRef PubMed.
- T. D. W. Claridge, M. Mayzel and E. Kupče, Magn. Reson. Chem., 2019, 57, 946–952 CrossRef CAS PubMed.
- E. Kupče and T. D. W. Claridge, J. Magn. Reson., 2019, 307, 106568 CrossRef PubMed.
- E. Kupče and T. D. W. Claridge, Chem. Commun., 2018, 54, 7139–7142 RSC.
- E. Kupče, J. R. J. Yong, G. Widmalm and T. D. W. Claridge, JACS Au, 2021, 1, 1892–1897 CrossRef PubMed.
- A. L. Hansen, E. Kupče, D.-W. Li, L. Bruschweiler-Li, C. Wang and R. Brüschweiler, Anal. Chem., 2021, 93, 6112–6119 CrossRef CAS PubMed.
- F. Tang and E. Hatzakis, Anal. Chem., 2020, 92, 11177–11185 CrossRef CAS PubMed.
- P. Bellstedt, Y. Ihle, C. Wiedemann, A. Kirschstein, C. Herbst, M. Görlach and R. Ramachandran, Sci. Rep., 2014, 4, 4490 CrossRef PubMed.
- C. Wiedemann, P. Bellstedt, A. Kirschstein, S. Häfner, C. Herbst, M. Görlach and R. Ramachandran, J. Magn. Reson., 2014, 239, 23–28 CrossRef CAS PubMed.
- E. Kupče and R. Freeman, Magn. Reson. Chem., 2007, 45, 2–4 CrossRef PubMed.
- F. Delaglio, S. Grzesiek, G. W. Vuister, G. Zhu, J. Pfeifer and A. Bax, J. Biomol. NMR, 1995, 6, 277–293 CrossRef CAS PubMed.
- T. Xie, T. Saleh, P. Rossi and C. G. Kalodimos, Science, 2020, 370, eabc2754 CrossRef CAS PubMed.
- Y. Gu, A. L. Hansen, Y. Peng and R. Brüschweiler, Angew. Chem., Int. Ed., 2016, 55, 3117–3119 CrossRef CAS PubMed.
- T. Yuwen, L. E. Kay and G. Bouvignies, ChemPhysChem, 2018, 19, 1707–1710 CrossRef CAS PubMed.
- M. Leninger, W. M. Marsiglia, A. Jerschow and N. J. Traaseth, J. Biomol. NMR, 2018, 71, 19–30 CrossRef CAS PubMed.
- G. Jameson, A. L. Hansen, D. Li, L. Bruschweiler-Li and R. Brüschweiler, J. Am. Chem. Soc., 2019, 141, 16829–16838 CrossRef CAS PubMed.
- K. Kazimierczuk and V. Orekhov, Magn. Reson. Chem., 2015, 53, 921–926 CrossRef CAS PubMed.
- J. R. J. Yong, E. Kupče and T. D. W. Claridge, Anal. Chem., 2022, 94, 2271–2278 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Details of the NOAH-(15N/13C)-CEST sequence and details of the acquisition of the experiments and methods employed. See DOI: https://doi.org/10.1039/d2cc02015g |
| This journal is © The Royal Society of Chemistry 2022 |