
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Assessment of presumed small-molecule ligands of telomeric i-DNA by biolayer interferometry (BLI)†
Hugues
Bonnet
a,
Maéva
Morel
a,
Alexandre
Devaux
a,
Joseph
Boissieras
b,
Anton
Granzhan
 b,
Benjamin
Elias
c,
Thomas
Lavergne
*a,
Jérôme
Dejeu
*ad and
Eric
Defrancq
*a
b,
Benjamin
Elias
c,
Thomas
Lavergne
*a,
Jérôme
Dejeu
*ad and
Eric
Defrancq
*a
aUniversité Grenoble Alpes, Département de Chimie Moléculaire, UMR CNRS 5250, CS 40700-38058 Grenoble, France. E-mail: eric.defrancq@univ-grenoble-alpes.fr; jerome.dejeu@univ-grenoble-alpes.fr; thomas.lavergne@univ-grenoble-alpes.fr
bInstitut Curie, CNRS UMR 9187, INSERM U1196, Université Paris Saclay, 91405 Orsay, France
cInstitut de la Matière Condensée et des Nanosciences (IMCN), Molecular Chemistry, Materials and Catalysis (MOST), Université catholique de Louvain (UCLouvain), Place Louis Pasteur 1, 1348 Louvain-la-Neuve, Belgium
dFEMTO-ST Institute, CNRS UMR-6174, Université de Bourgogne Franche-Comté, F-25000 Besançon, France
First published on 30th March 2022
Abstract
Biolayer interferometry (BLI) and circular dichroism (CD) spectroscopy were used to investigate the interaction between previously reported i-motif DNA (i-DNA) ligands and folded or unfolded i-DNA in acidic (pH 5.5) and near-neutral (pH 6.5) conditions. We observed that although several ligands, in particular macrocyclic bis-acridine (BisA) and pyridostatin (PDS), showed good affinities for the telomeric i-motif forming sequence, none of the ligands displayed selective interactions with the i-DNA structure nor was able to promote its formation.
i-Motifs of DNA (hereafter, i-DNA), known in vitro for nearly three decades,1 are unusual four-stranded structures, in which cytosines are intercalated via a stack of hemi-protonated C–C base pairs (CH+:C).2 Some of these DNA structures have been well characterized in vitro and, because i-DNA may mirror other four-stranded G-rich structures (G-quadruplexes or G4) present in gene promoters or at telomeres, their biological relevance has been investigated.3 Several studies indicated that i-DNA formation modulates transcription,4 and a recent study using a specific antibody (iMab) revealed their presence in the nuclei of human cells, co-localizing with either telomeres or transcription factors.5 Trantirek and colleagues also demonstrated, using in-cell NMR, that exogenous i-DNA are stable in the nuclei of human cells.6
However, our knowledge about i-DNA biology is still limited, in part due to the lack of suitable small-molecule probes to interrogate the biological roles of these structures. For example, most of our knowledge on the impact of four-stranded DNA secondary structures, such as G4, on cell metabolism resulted from the use of selective G4 ligands and the characterization of proteins that bind or modulate the formation of such structures.7,8 In contrast, relatively few molecules were reported to interact with i-DNA.9 The main issues in this regard are the strong pH-dependency, flexibility, polymorphism and complex folding behavior of i-DNA, which introduce potential bias into screening methods. In particular, low-pH conditions used to induce the formation of i-DNA lead to the protonation of many ligands, which can strongly increase their non-specific nucleic acid binding. This latter point is particularly critical because the use of small molecules to study the biological functions of such structures is essential. To date, molecules including TMPyP4,10 mitoxantrone,11 IMC-48,12 [Ru(phen)2dppz]2+,13 berberine,14 and others such as PhenDC3, BRACO-19 and PDS have been reported as putative i-DNA ligands. However, a strong controversy persists with regard to their binding mode, affinity, and selectivity for i-DNA with respect to single-stranded, duplex or G4-DNA forms.15,16 The identification and evaluation of i-DNA ligands are hampered by the lack of robust, uniform assays such as FRET-melting and FID assay which became routine in the field of G4 ligands. In fact, these methods have strong biases and provide untrustworthy results with i-DNA, as evidenced in recent publications,17,18 prohibiting direct comparison of the ligands described by different groups.
In this context, we have previously assembled two peptide-DNA conjugates that form i-motif structures,19,20 with one of them, namely conjugate 2 (Fig. 1), being able to fold into a stable i-motif at room temperature and, most importantly, at near-neutral pH.19 This stabilized mimic of the i-motif adopted by the h-telo sequence may be used to study the interaction of the i-motif with putative ligands at a more physiologically relevant pH. As a proof of concept, we report herein a comparative biolayer interferometry (BLI) study of the interaction of several of the most promising i-DNA ligands reported to date, with the native i-motif structure of telomeric DNA (h-telo 1 in Fig. 1) along with the stabilized DNA-peptide conjugate 2. BLI has recently emerged as a powerful optical technique for isothermal characterization and quantification of interactions between molecules, in particular because it does not require any microfluidic set-up and enables real-time and label-free characterization of the interactions with the determination of affinity and selectivity in a 96-well microplate format. It has already been used to study the interactions of ligands with G-quadruplex DNA.21–23
 | ||
| Fig. 1 Structure of the native telomeric i-motif h-telo 1, i-motif forming DNA-peptide conjugate 2, hairpin control 3 and single-stranded control 4. | ||
The interactions with putative ligands were investigated at two distinct pH values, an acidic pH of 5.5 at which both h-telo 1 and mimic 2 are folded into an i-motif structure as confirmed by CD analysis and a near-neutral pH of 6.5 at which the native telomeric sequence does not form an i-motif unlike the stabilized i-motif 2 (Fig. S20, ESI†).24 Hairpin (HP) 3 and single-stranded (SS) DNA 4 were also used in this study as controls. To perform BLI analysis, biomolecular systems 1–4 were anchored on the BLI sensors through biotin–streptavidin interactions. Nine ligands (Fig. 2) including the aforementioned state-of-the-art i-motif binders and previously reported i-DNA interactors were investigated for their interaction with the DNA structures 1–4. The acquired sensorgrams allowed for the determination of the equilibrium dissociation constants of the interaction from the responses obtained at the steady state (Δλ) based on a 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1 interaction model and after fitting of the Langmuir isotherm (Table 1 and Fig. S2–S19, ESI†).
:1 interaction model and after fitting of the Langmuir isotherm (Table 1 and Fig. S2–S19, ESI†).
 | ||
| Fig. 2 Structures of the ligands investigated in the study. | ||
| Ligand | H-telo i-DNA 1 | Constrained i-DNA 2 | HP control 3 | SS control 4 | ||||
|---|---|---|---|---|---|---|---|---|
| pH 5.5 | pH 6.5 | pH 5.5 | pH 6.5 | pH 5.5 | pH 6.5 | pH 5.5 | pH 6.5 | |
| PhenDC3 | 2 ± 0.3 | 6 ± 0.2 | 3.5 ± 1 | 6 ± 0.4 | 6.8 ± 0.9 | 4.5 ± 0.9 | 2 ± 0.5 | 2.3 ± 0.6 |
| PDS | 0.12 ± 0.04 | 0.32 ± 0.02 | 0.26 ± 0.04 | 0.36 ± 0.06 | 1.2 ± 0.2 | 1.3 ± 0.1 | 0.12 ± 0.03 | 0.10 ± 0.03 |
| TMPyP4 | 4.8 ± 0.6 | 8.7 ± 2 | 14 ± 2 | 10.7 ± 3 | 4.1 ± 1.6 | 8.8 ± 3 | 3.4 ± 0.3 | 9.8 ± 2 |
| Braco-19 | 4.6 ± 0.7 | 11 ± 3 | 9.5 ± 2 | 8.5 ± 2 | 15 ± 3 | 18.2 ± 4 | 4.1 ± 0.9 | 4.6 ± 0.9 |
| Berberine | n.d. | n.d. | n.d. | n.d. | 44 ± 12 | n.d. | n.d. | n.d. |
| [Ru(Phen)2dppz]2+ | n.d. | n.d. | n.d. | n.d. | 37 ± 16 | 54 ± 10 | 9.6 ± 1 | 18 ± 2 |
| RHPS4 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| BisA | 0.06 ± 0.01 | 0.22 ± 0.06 | 0.13 ± 0.06 | 0.29 ± 0.1 | 2.1 ± 0.2 | 8.45 ± 3 | 0.21 ± 0.04 | 0.35 ± 0.08 |
| Mitoxantrone | 11 ± 2 | 12 ± 0.9 | 19 ± 3 | 13 ± 4 | 72 ± 17 | 87 ± 21 | 10 ± 2 | 11 ± 2 |
Previous studies, performed with native, non-stabilized, i-motif h-telo sequence 1, were mostly done at the acidic pH of 5.5 to ensure the formation of the i-motif. Our current data for h-telo 1 at pH 5.5 are in part concordant with these previous reports. Specifically, for mitoxantrone, we found a KD value of 11 μM which agrees with the previously determined affinity using SPR measurements.11 Likewise, PhenDC3 showed a KD value around 2 μM which is consistent with the value of 4 μM obtained with related phenanthroline compounds using fluorometric titrations.25 For TMPyP4, a previous report suggested 1 μM affinity for the human telomeric i-motif at pH 5.2, which is quite close to the KD value of 4.8 μM observed in the present study.26 It should also be noted that the techniques used for the determination of affinity (UV and CD titrations) differ from our BLI method. For berberine, a KD value of 19.6 μM was previously reported by using fluorescence titration, while no significant signal variation was measured by BLI and the Langmuir isotherm did not converge,27 thus confirming the very weak affinity of berberine for i-DNA. We were unable to measure a KD value for [Ru(phen)2dppz]2+ (racemic mixture), whereas an interaction of this complex with i-DNA was reported in the literature by using UV melting and luminescence.13 However, the i-DNA sequences used for this study were not the same and the authors concluded that [Ru(phen)2dppz]2+ did not stabilize short-looped sequences but were able to stabilize the relatively less stable, long-looped i-motifs. Therefore, our results further confirm the null to weak affinity of [Ru(phen)2dppz]2+ for short-looped i-motif forming sequences. For Braco-19, a well-known G4 ligand, the relatively low affinity observed (4.6 μM) associated with poor selectivity against control hairpin would preclude the use of such a molecule as an i-motif binder. The same conclusion can be reached for RHPS4 (also a known G4 binder) for which no binding could be determined in the range of used concentrations. Finally, BisA and PDS appeared to be the best ligands for i-DNA 1 with KD values of 60 nM and 120 nM, respectively.
Promisingly, we found virtually no difference (within three-fold) between the affinities of the studied ligands for the native i-DNA 1 and the constrained i-DNA 2 at pH 5.5 which appears to confirm the structural similarity of both i-DNA in these acidic conditions and the absence of any detrimental effect of the cyclic peptide scaffold on the recognition properties as already reported.28 Encouragingly, the affinities measured with 2 at pH 6.5 were found to be very similar to the ones measured at pH 5.5 for both i-DNA 1 and 2. This indicates that constrained i-DNA 2 is also a potent mimic of i-motif DNA at a higher pH. Surprisingly, by using 1 at pH 6.5, i.e., pH at which the sequence is not expected to fold into an i-motif, we also observed affinities similar to those obtained at pH 5.5 as well as for the interaction of the ligands with constrained i-DNA 2.
Similar affinities for the interaction of the ligands with presumably folded (pH 5.5) and unfolded (pH 6.5) structures could be interpreted in multiple ways: (i) i-DNA 1 might be partially or fully unfolded by the ligands at pH 5.5 and the resulting structure would compare to the one obtained at pH 6.5; (ii) conversely, the interaction of the ligands with the unfolded i-DNA 1 at pH 6.5 may trigger folding into an i-motif structure such as the one observed at pH 5.5 and present in i-DNA 2; (iii) the measured affinities do not depend on the presence of the folded i-motif structure but merely reflects interactions with the cytosine-containing sequences or (iv) the interactions are due to non-specific interactions with the DNA backbone.
To further investigate the effect of the ligands, we used CD analysis to gauge the folding status of i-DNA 1 in the presence of each ligand (5 molar equivalents)29 at both pH 5.5 and 6.5. At pH 5.5 (Fig. S21, ESI†) we observed that all the compounds induced a hypochromic effect of the band at 288 nm which was particularly marked in the case of Braco-19, PDS, PhenDC3 and mitoxantrone. This agrees with the observations of Randazzo and colleagues and could suggest that the interaction with some ligands induces partial unfolding of the i-motif structure.30 Nevertheless, the i-motif appears to remain the preponderant structure in the presence of the ligands at pH 5.5 ruling out the first hypothesis. Inversely, the CD spectra obtained at pH 6.5 (Fig. S22, ESI†) demonstrate that no ligand could efficiently promote the folding of the unstructured i-DNA 1 sequence into a well-defined i-motif structure, as the spectra remain unchanged after addition of the ligands, ruling out the second hypothesis. Finally, we recorded CD spectra under intermediate conditions (pH 6.2, i.e., close to pHT) at which i-DNA 1 was found in an equilibrium between folded and unfolded forms, hypothesizing that in this case even small effects of the ligands would manifest by shifting the equilibrium towards either i-motif or single-stranded forms (Fig. S23, ESI†). In these experiments, the ratio of CD signals at 285 nm (maximum of the i-motif form) and 275 nm (maximum of the single-stranded form) was used as a metric of the ligand-induced effect. The results (Fig. 3) showed that, among the tested compounds, PDS and Braco-19 (at 2 or 5 molar equiv.), BisA (at 5 molar equiv.) as well as PhenDC3 (at 2 molar equiv. only) seemingly increased the θ285nm/θ275nm ratio, suggesting an induction of the i-motif form. However, a close inspection of the CD spectra (Fig. S23, ESI†) revealed that, in all these cases, addition of the ligand reduced the intensity of the signal at 275 nm without a concomitant increase of the i-motif peak.
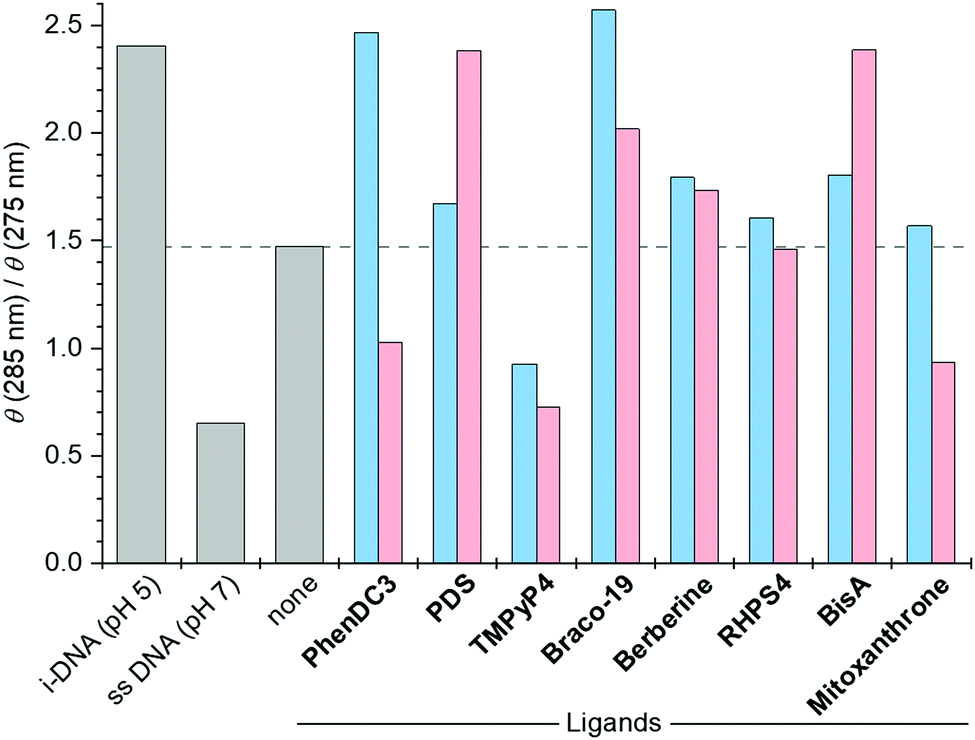 | ||
| Fig. 3 Ratio of CD intensity at 285 nm (i-DNA) and 275 nm (single-stranded form) of solutions of h-telo DNA 1 at pH 6.2 in the absence and in the presence of 2 (blue bars) and 5 (pink bars) molar equiv. of ligands. | ||
In all other cases, the addition of ligands clearly shifted the equilibrium towards the single-stranded form (Fig. 3 and Fig. S23, ESI†), which might suggest that those ligands may bind to single-stranded DNA. To further explore these hypotheses, we performed BLI analysis of the interactions between single-stranded (poly dT) control 4 (Fig. 1) and all ligands at both pH 5.5 and 6.5 (Table 1). Surprisingly, at both pH values, KD values similar to the ones obtained with i-motif forming DNA 1 and 2 were measured for most of the ligands, further suggesting a strong interaction of the ligands with the single-stranded oligonucleotide. Thus, the binding constants observed with substrates 1 and 2 insinuate the absence of specific recognition of i-motif structural features. Interestingly, the inspection of CD spectra recorded in equilibrium conditions (pH 6.2, Fig. S23, ESI†) revealed the presence of induced CD (iCD) bands in the region of ligand absorption in the case of PhenDC3, [Ru(phen)2dppz]2+ and Braco-19. As iCD is typically considered as solid evidence of the ligand–DNA interaction, we additionally recorded the iCD spectra of these ligands at pH 5.5 and 7.3, in order to elucidate whether these signals arise from binding of ligands to i-DNA or unfolded DNA, respectively (Fig. S24, ESI†). In the case of PhenDC3, the iCD signal observed at pH 6.2 matched the one observed at pH 7.3, giving evidence of binding to unfolded h-telo. Conversely, in the case of Braco-19, the iCD signal observed at pH 6.2 was similar to the one observed at pH 5.5, which is in favor of the interaction of this ligand with folded i-DNA. Finally, in the case of [Ru(phen)2dppz]2+, no iCD was observed either at pH 5.5 or at pH 7.3, giving evidence of a complex binding behavior.
In summary, we investigated the affinities of nine state-of-the-art small-molecule binders of unusual nucleic acid structures, previously reported as i-motif targeting agents. BLI analysis performed with folded (at pH 5.5 and 6.5) and unfolded (at pH 6.5) i-motifs derived from the human telomeric sequence uncovered massive binding promiscuity. Indeed, even though some of the ligands (BisA and PDS) demonstrated good affinities for the i-motif forming C-rich sequence and selectivity over hairpin DNA, none of the ligands were shown to discriminate between folded and unfolded i-motif structures, nor shift the equilibrium towards the folded i-motif as confirmed by CD.
Importantly most of the studied ligands showed comparable affinity to single-strand DNA not able to form i-motifs. Even though the analysis was not performed at physiological pH at which the physical properties of the ligands may vary, this result still sheds further light on the discrepancy that surrounds the endeavor to identify specific i-motif ligands. In this context, the use of constrained i-DNA that ensures an i-motif folding represents an interesting alternative to identify unambiguous (i.e., affine and specific) i-DNA-interacting ligands.
H. B., M. M, A. D., J. B.: investigation; A. G., B. E.: resources, writing, review and editing; T. L: supervision, writing, review and editing; J. D.: formal analysis, supervision, writing, review and editing; E. D.: supervision, funding acquisition, writing, review and editing.
This work was partially supported by the “Agence Nationale de la Recherche” (ANR-21-CE44-0005-02), the University Grenoble Alpes Graduate School (ANR-17-EURE-0003), and the CNRS through the MITI interdisciplinary program (PRIME’80). ICMG (UAR 2607) is acknowledged for facilities access. We thank Dr M.-P. Teulade-Fichou for a generous gift of BisA.
Conflicts of interest
There are no conflicts to declare.Notes and references
- K. Gehring, J.-L. Mergny and M. Guéron, Nature, 1993, 363, 561 CrossRef CAS PubMed.
- J.-L. Mergny and D. Sen, Chem. Rev., 2019, 119, 6290 CrossRef CAS PubMed.
- H. Abou Assi, M. Garavis and M. J. Damha, Nucleic Acids Res., 2018, 46, 8038 CrossRef PubMed.
- S. Kendrick, H.-J. Kang, M. P. Alam, M. M. Madathil, P. Agrawal, V. Gokhale, D. Yang, S. M. Hecht and L. H. Hurley, J. Am. Chem. Soc., 2014, 136, 4161 CrossRef CAS PubMed.
- M. Zeraati, D. B. Langley, P. Schofield, A. L. Moye, R. Rouet, W. E. Hughes, T. M. Bryan, M. E. Dinger and D. Christ, Nat. Chem., 2018, 10, 631 CrossRef CAS PubMed.
- S. Dzatko, M. Krafcikova, R. Hansel-Hertsch, T. Fessi, R. Fiala, T. Loja, D. Krafcik, J.-L. Mergny, S. Foldynova-Trantirkova and L. Trantirek, Angew. Chem., Int. Ed., 2018, 57, 2165 CrossRef CAS PubMed.
- J. Carvalho, J.-L. Mergny, G. F. Salgado, J. A. Queiroz and C. Cruz, Trends Mol. Med., 2020, 26, 848 CrossRef CAS PubMed.
- R. Hansel-Hertsch, M. Di Antonio and S. Balasubramanian, Nat. Rev. Mol. Cell Biol., 2017, 18, 279 CrossRef CAS PubMed.
- S. L. Brown and S. Kendrick, Pharmaceuticals, 2021, 14, 96 CrossRef CAS PubMed.
- L. Martino, B. Pagano, I. Fotticchia, S. Neidle and C. Giancola, J. Phys. Chem. B, 2009, 113, 14779 CrossRef CAS PubMed.
- E. P. Wright, H. A. Day, A. M. Ibrahim, J. Kumar, L. J. E. Boswell, C. Huguin, C. E. M. Stevenson, K. Pors and Z. A. E. Waller, Sci. Rep., 2016, 6, 39456 CrossRef CAS PubMed.
- H.-J. Kang, S. Kendrick, S. M. Hecht and L. H. Hurley, J. Am. Chem. Soc., 2014, 136, 4172 CrossRef CAS PubMed.
- B. J. Pages, S. P. Gurung, K. McQuaid, J. P. Hall, C. J. Cardin and J. A. Brazier, Front. Chem., 2019, 7, 744 CrossRef CAS PubMed.
- L. Xu, S. Hong, N. Sun, K. Wang, L. Zhou, L. Jia and R. Pei, Chem. Commun., 2016, 52, 179 RSC.
- H. A. Day, P. Pavlos and Z. A. E. Waller, Bioorg. Med. Chem., 2014, 22, 4407 CrossRef CAS PubMed.
- S. S. Masoud and K. Nagasawa, Chem. Pharm. Bull., 2018, 66, 1091 CrossRef CAS PubMed.
- V. B. Tsvetkov, A. V. Turaev, N. A. Petrunina, D. M. Melnik, Y. M. Khodarovich, G. E. Pozmogova, T. S. Zatsepin, A. M. Varizhuk and A. V. Aralov, Analyst, 2021, 146, 4436 RSC.
- P. Alberti, J. Ren, M.-P. Teulade-Fichou, L. Guittat, J.-F. Riou, J. B. Chaires, C. Helene, J.-P. Vigneron, J.-M. Lehn and J.-L. Mergny, J. Biomol. Struct. Dyn., 2001, 19, 505 CrossRef CAS PubMed.
- A. Devaux, L. Bonnat, T. Lavergne and E. Defrancq, Org. Biomol. Chem., 2020, 18, 6394 RSC.
- R. Bonnet, P. Murat, N. Spinelli and E. Defrancq, Chem. Commun., 2012, 48, 5992 RSC.
- L. Bonnat, M. Dautriche, T. Saidi, J. Revol-Cavalier, J. Dejeu, E. Defrancq and T. Lavergne, Org. Biomol. Chem., 2019, 17, 8726 RSC.
- M. Gillard, J. Weynand, H. Bonnet, F. Loiseau, A. Decottignies, J. Dejeu, E. Defrancq and B. Elias, Chem. – Eur. J., 2020, 26, 13849 CrossRef CAS PubMed.
- J. Weynand, H. Bonnet, F. Loiseau, J.-L. Ravanat, J. Dejeu, E. Defrancq and B. Elias, Chem. – Eur. J., 2019, 25, 12730 CrossRef CAS PubMed.
- The positive band observed at 288 nm and the negative one at 260 nm are characteristic of an i-motif folding topology: K. Guo, V. Gokhale, L. H. Hurley and D. Sun, Nucleic Acids Res., 2008, 36, 4598 CrossRef CAS PubMed.
- L. Wang, Y. Wu, T. Chen and C. Wei, Int. J. Biol. Macromol, 2013, 52, 1 CrossRef CAS PubMed.
- S. Fernandez, R. Eritja, A. Avino, J. Jaumot and R. Gargallo, Int. J. Biol. Macromol., 2011, 49, 729 CrossRef CAS PubMed.
- R. Gargallo, A. Avino, R. Eritja, P. Jarosova, S. Mazzini, L. Scaglioni and P. Taborsky, Spectrochim. Acta, Part A, 2021, 248, 119185 CrossRef CAS PubMed.
- P. Murat, R. Bonnet, A. Van der Heyden, N. Spinelli, P. Labbe, D. Monchaud, M.-P. Teulade-Fichou, P. Dumy and E. Defrancq, Chem. – Eur. J., 2010, 16, 6106 CrossRef CAS PubMed.
- Due to the absence of measured affinity in the BLI analysis, berberine, [Ru(Phen)2dppz]2+ and RHPS4 ligands were not studied by CD.
- A. Pagano, N. Iaccarino, M. A. S. Abdelhamid, D. Brancaccio, E. U. Garzarella, A. Di Porzio, E. Novellino, Z. A. E. Waller, B. Pagano, J. Amato and A. Randazzo, Front. Chem., 2018, 6, 1 CrossRef PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Sensorgrams, Langmuir isotherm, experimental details, CD analysis, and calculations. See DOI: 10.1039/d2cc00836j |
| This journal is © The Royal Society of Chemistry 2022 |