
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceThe influence of pH on dissolved organic matter fluorescence in inland waters†
M.
Groeneveld
 a,
N.
Catalán
b,
K.
Einarsdottir
a,
A. G.
Bravo
c and
D. N.
Kothawala
*a
a,
N.
Catalán
b,
K.
Einarsdottir
a,
A. G.
Bravo
c and
D. N.
Kothawala
*a
aDepartment of Ecology and Genetics/Limnology, Evolutionary Biology Center, Uppsala University, Norbyvägen 18D, 75236, Uppsala, Sweden
bLaboratoire des Sciences du Climat et l'Environnement (LSCE), CNRS-UMR 8212, L'Orme des Merisiers-bat.714, 91190 Gif-sur-Yvette, France
cDepartament de Biologia Marina i Oceanografia Institut de Ciències del Mar, CSIC, Pg Marítim de la Barceloneta 37-49, E08003 Barcelona, Catalunya, Spain
First published on 17th March 2022
Abstract
Fluorescence is an easily available analytical technique used to assess the optical characteristics of dissolved organic matter (DOM). Despite widespread use, there has been some confusion about how robust fluorescence spectroscopy is to differences in solution pH. Here we assess fluorescence characteristics of three natural water samples and one commercially available standard (Nordic Reservoir) by modifying the pH across a range from 3.5 to 9.0 at 0.5 pH increments. We used two statistical approaches to assess if fluorescence intensity shifted significantly across this pH range. We identified that humic-like and protein-like fluorescence was largely stable within the pH range of 5.5 to 7.5, which represents 80% of Swedish lakes and streams. Likewise, we found that the three commonly used fluorescence indices were robust across the full pH range tested with the exception of the humification index, which had a narrower range of stability. The commerical humic substance sample was highly unstable with changes to pH in the regions of protein-like fluorescence being particularly sensitive. One of our conclusions is that differences in fluorescence intensity in the pH range of 5.5 to 7.5, typical for most inland waters, are generally minor. We recommend adjusting the pH when samples fall outside this region and to be especially careful in interpreting results from commercial humic substances.
Introduction
Fluorescence spectroscopy is a widely used and simple technique used to characterize the molecular characteristics of dissolved organic matter (DOM) in freshwaters, soils and marine environments.1–3 Fluorescence scans produce a three-dimensional spectrum expressing the fluorescence intensity across a range of excitation and emission wavelengths simultaneously. This resulting spectrum is referred to as an excitation–emission matrix (EEM). By comparing the intensity of fluorescence in distinct regions of the EEM, one can obtain information regarding the origin of DOM, which is broadly categorized into two types: humic-like fluorescence, which represents material of terrestrial origin, and protein-like fluorescence, which largely reflects aquatically produced material of microbial and algal origin. DOM is comprised of several hundred thousand molecules and not all DOM molecules contain a fluorophore, however fluorescence has proven to be a highly effective fingerprinting tool to track the source and processing of DOM.3–5 A mixture of DOM compounds can comprise a wide range of intrinsic fluorophores, from condensed aromatic moieties to unsaturated aliphatic chains.6 Consequently, each fluorescence spectrum incorporates the structural complexity and heterogeneity of numerous organic compounds comprising DOM. There are however several factors that need to be considered when measuring and interpreting DOM fluorescence, including the solution pH.7–9 The state of protonation can influence the effectiveness of DOM compounds to fluoresce in multiple ways. The hydrogen ion can potentially act as a quencher for specific fluorophore units, whereby fluorescence intensity decreases with lowering pH.10 When the pH of a solution is equal to the acid dissociation constant (pKa) of a compound, half of the acidic group binding sites are deprotonated with a negative charge and the other half are saturated with a hydrogen ion. When the pH is lower than the pKa, more sites are protonated and the opportunity for repulsion between two negative sites within a DOM molecule decreases. Thus, the conformational arrangement and hydrodynamic density of a molecule can be influenced by pH due to alterations in inter- and intra-molecular interactions. Accordingly, a given fluorophore may become more or less exposed to incoming light depending on the molecular configuration of DOM compounds.11,12 Ions other than H+ can also potentially affect DOM molecular configuration, inter- and intramolecular interactions, and so ionic strength may affect the DOM's light absorbing properties in a similar manner.13In studies using DOM fluorescence, it is a common practice to analyse and compare DOM from water samples spanning across a wide natural gradient of conditions including pH. Many factors can contribute to variability in ambient pH including differences in underlying bedrock composition, vegetation, seasonal flushing of spring snowpack, which can have a diluting effect, or anthropogenic inputs. Thus, it is of fundamental importance to understand the influence of pH on fluorescence intensity across the full EEM, and provide a comprehensive set of recommendations on how and when to pre-treat or interpret data. Additionally, it is useful to evaluate the influence of pH on commonly used fluorescence indices, which are ratios of different regions of the EEM, such as the fluorescence index,14,15 the freshness index16 and the humification index.17 In past studies, the influence of pH on the ratio of common fluorescence indices has been largely overlooked and requires investigation.
Our current understanding of pH effects on fluorescence spectra of DOM remains fraught with confusion. This is in part due to variable study designs producing contrasting results which results in different recommendations. For instance, a study using commercially available humic substances derived from soil and water claimed that pH had a strong influence on fluorescence spectra.18 Another study also found pH to have a strong influence on the fluorescence of commercial humic substances, but not for natural water samples.19 Several additional studies evaluating the influence of pH on humic-like fluorescence in natural waters, reported little change in the fluorescence intensity with pH.8,20 An important aspect to consider as we expand our understanding of pH effects on DOM fluorescence is to avoid an assumption that all regions of the EEM spectra respond similarly. Traditionally, many studies have restricted their interpretation to regions of the EEM associated with humic-like or fulvic-like fluorescence,8,20 thereby leaving it unclear whether the protein-like region behaves similarly. The fluorescence intensity of the protein-like region, including tyrosine-like and tryptophan-like fluorescence, is often independent of the humic-like region and is unlikely to be influenced by pH in a comparable manner. Reduced fluorescence intensity due to protonation of a fluorescing functional group can have a quenching effect, which has been observed with the pure amino acid tryptophan.21 However, these results should not be directly applied to tryptophan-like fluorescence observed in DOM spectra without being properly tested due to the possibility of other interactions. Many studies have gleaned important insight from protein-like fluorescence, which is considered to represent aquatically produced DOM in comparison to terrestrial DOM which tends to fluoresce at longer emission wavelengths.22,23 Thus, it is particularly relevant to test the sensitivity of tryptophan-like fluorescence for these reasons. Currently, it remains unclear which regions of the EEM are most sensitive to changes in pH, and to what scale the different regions are affected across the pH gradient most relevant for inland waters. Furthermore, it is unclear why there might be discrepancies in results from natural lake waters relative to commercial humic standards, which were used more commonly in earlier studies to test pH effects.18,24 Approaches used to prepare commercial standards are variable, yet they often are dried to a powder and re-constituted in water with the final solution containing an ionic composition differing from natural conditions.
A methodological limitation of some previous studies investigating the effect of pH is a limited number of pH levels being tested and restricting analysis to extremely acidic or basic conditions that lie well outside of natural conditions (e.g. pH 2, 6 and 10;18 or original pH ± 2 pH units19). Several studies have used one pH unit increments,8,20,25 however 0.5 pH unit steps could be particularly insightful for determining where shifts occur along the pH gradient, and the magnitude of the change in fluorescence due to pH changes. Measurements at high resolution and with focus placed on the pH range typical of inland waters can help provide more concrete and meaningful recommendations.
This study aims to quantify the relative sensitivity of DOM fluorescence to changing pH across the full EEM, focusing on five commonly studied ‘peaks’ including two protein-like regions (tryptophan-like and tyrosine-like), and the relative sensitivity of three common fluorescence indices (FI, Fresh, HIX). The study compares and contrasts results from three natural water samples and one commercially available extract (Nordic Reservoir Humic Substance) across a pH from 3.5 to 9.0 with increments of 0.5 units, with focus placed on the pH range (5.5 to 7.5) most relevant to natural freshwaters.
Experimental
Samples
Natural lake water was collected, and filtered with 0.7 μm glass fiber filters (GF/F, Whatman), from three sites in South-Central Sweden; Strandsjön (25.5 mg L−1 DOC, ambient pH 7.1), Krycksjön (26.8 mg L−1 DOC, ambient pH 7.7) and Ängbybadet (15.5 mg L−1 DOC, ambient pH 7.6). The samples were collected in March and April, and after filtering stored in the dark at 4 °C and analyzed in May and June of 2014, after being re-filtered (0.7 μm GF/F). One commercially available extracted humic substance sample from a reservoir located in Norway was included, called Nordic Reservoir NOM (IHSS, Ref. 1R108N), and was prepared by dissolving it in Milli-Q water with a starting pH of 5.9 (15.0 mg L−1 DOC). The optical properties of all samples are given in ESI Table 1,† and are plotted relative to the distribution of lakes across Sweden in ESI Fig. 1.† All samples were analysed for non-purgeable organic carbon concentration after being acidified using a total carbon analyser (Shimadzu TOC-L), which we refer to as the dissolved organic carbon (DOC) concentration.Algal extract
To ensure all samples would produce measurable protein-like fluorescence, a pre-prepared algal extract was added to all samples. The green algae Scenedesmus sp. (Chlorophyceae) is a common member of planktonic communities around the world.26 The algae was cultured axenically for 14 days at 20 °C under a 12 h dark/12 h light photoperiod with a growth chamber (Nüve GC 400) in a Z8 medium consisting of essential nutrients (e.g. NO3−, K+, and Fe2+).27 The algal cultures were concentrated and dried to a powder form by centrifugation followed by freeze drying. Subsequently, the algal powder was added to the stock solution of each sample (0.06 g per 0.5 L sample). This stock solution was subsequently poured into individual 40 mL glass vials, and subsequently used for pH adjustments. The stock solution was stirred with a magnetic stirring rod in a 60 °C warm water bath for a few minutes until the powder dissolved, and re-filtered (GF/F, Whatman) to remove the residual particulate algal material. A separate algal stock solution was made for the pH experiments and ionic strength experiments, because the test for ionic strength was performed subsequent to the pH experiments.pH and ionic strength experiments
Experiments to test pH effects included three natural water samples and the commercial standard. Each of the four samples was allowed to reach room temperature (20 °C), and divided over 12 acid-washed and pre-combusted (450 °C, 4.5 hours) 40 mL glass vials. The pH of each sample was adjusted with either 1 M NaOH or 10% HCl, to span a pH range from 3.5 to 9.0, at 0.5 pH unit increments. The acid and base were added in precise steps of 1 to 5 μL, such that the total added volume remained <0.5% of the total sample volume. All pH measurements were recorded using a pH electrode (Metrohm) designed for the pH range 0 to 14, and temperatures from 0 to 80 °C and a pH meter, which was calibrated daily, and re-calibrated if necessary when standards were checked periodically (744 Metrohm, Switzerland). A magnetic stirring bar was used for a few seconds to properly mix the acid or base, but pH measurements were taken on still and equilibrated samples.A separate experiment was performed to test the influence of ionic strength on the commercially available Nordic Reservoir sample using four ionic strength concentrations (0.0, 0.5, 1.0 and 2.0 mM). A stock solution of 4.0 mM containing a combination of NaCl (40 mg L−1), CaCl2–2H2O (80 mg L−1) and K2SO4 (96 mg L−1) was made, and subsequently diluted to 0.5, 1.0 and 2.0 mM to represent the range typical of inland waters. Milli-Q water was used for the 0.0 mM solution which is used as the reference solution for statistical analysis since this is how the standard is typically prepared. An aliquot of concentrated Nordic Reservoir solution (DOC ≈ 100 mg L−1) was added to each solution to make the DOC concentration ≈15 mg L−1. A set of 12 vials containing each solution was adjusted for pH ranging from 3.5 to 9.0, at 0.5 pH increments, using the same protocol described above.
Fluorescence measurements
All solutions adjusted for pH and ionic strength were analysed in triplicate for fluorescence using a spectrofluorometer (FluoroMax-4, Jobin Yvon, Horiba, Kyoto, Japan), and absorbance using a UV-visible spectrophotometer (Lambda 25, PerkinElmer, Waltham, USA). The absorbance spectra (250 to 600 nm) were measured at 1 nm intervals in a 1 cm quartz cuvette and Milli-Q water was used as the instrument blank. Fluorescence was measured in triplicate for each pH and ionic strength adjusted sample at excitation 250–445 nm with 5 nm increments, and emission 300–600 nm with 4 nm increments. Fluorescence spectra were blank subtracted, corrected for instrument biases and inner filter effects, and normalized to the Raman area of water.10,28,29 Changes in fluorescence intensity (Fint) were examined at five commonly known regions of the excitation–emission matrix,3 including three humic-like peaks that are representative of terrestrially derived DOM and incorporate substances that are considered to be humic and fulvic; peak A (λex: 260 nm, mean Fint at λem: 400–460 nm), peak C (mean Fintλex: 320–360 nm, mean Fintλem: 420–460 nm), peak M (mean Fintλex: 290–310 nm, mean Fintλem: 372–412 nm) and two protein-like peaks, including the tryptophan-like peak, peak T (λex: 275 nm, mean Fintλem: 340–352 nm) and the tyrosine-like peak, peak B (λex: 275 nm, mean Fintλem: 305 nm). Additionally, three common fluorescence indices were evaluated; (i) the fluorescence index, indicating if the source of organic matter is derived from more microbial and algal processing (≈1.8), or terrestrial sources (≈1.2) (FI = λem of 470: λem 520 nm, at an λex of 370 nm),14,30 (ii) the freshness index, with higher values representing more recently produced DOM (Fresh = Fint at λem 380 nm divided by the maximum intensity between λem 420 and 435 nm, at an λex of 310 nm),16 and (iii) the humification index, indicating the degree of condensation (HIX = peak area λem 435–480 nm/peak area λem 300–345 nm at λex 254 nm).17Method detection limit
Original water samples at ambient pH were analysed ten times for fluorescence and absorbance to determine the method detection limit (MDL = 1 standard deviation/mean × 100%), which includes the background variability attributable to instrumental fluctuations and measurement error associated with each sample. The MDL was variable between samples and peaks (from 2% to 13%, ESI Table 2†) and thus interpreted individually to give an indication of the background variability in Fint that could be attributed to sample analysis.Statistical analysis
To assess the pH sensitivity of fluorescence peaks and indices, the percent change in Fint for the peaks was calculated between each triplicate of the pH adjusted samples and the mean of triplicate samples at a reference pH of 6.5. The results were plotted to identify the pH range where fluorescence intensity remained within 5% of the reference value. A reference pH value was required to make a statistical comparison of deviations and a pH of 6.5 was determined from assessing the full distribution of pH values across Swedish surface waters, whereby the mean pH is 6.5 (median 6.8), with the lower 10th (pH 5.5) and upper 90th (pH 7.5) percentiles lying within ±1.0 pH units of the median (ESI Fig. 2†). The pH distribution included 48![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 358 data points including 7691 Swedish lakes and rivers and was compiled from the long-term monitoring program of the Swedish Agricultural University.31 A second statistical approach assessed if values across the full pH range (3.5 to 9.0) were statistically similar to the reference pH of 6.5 using a paired comparison of means (Student's t-test; p < 0.05) with the p-value adjusted by the Benjamini–Hochberg method of false discovery rate (FDR).32 When performing multiple comparisons, there is a chance of mistakenly identifying statistically significant relationships because some fraction of discoveries are false. The FDR therefore conceptualizes these type I errors for multiple comparisons as done here using the t-test. In summary, all comparisons are to test for similarity to values at pH 6.5, which represents 0% change in fluorescence intensity.
358 data points including 7691 Swedish lakes and rivers and was compiled from the long-term monitoring program of the Swedish Agricultural University.31 A second statistical approach assessed if values across the full pH range (3.5 to 9.0) were statistically similar to the reference pH of 6.5 using a paired comparison of means (Student's t-test; p < 0.05) with the p-value adjusted by the Benjamini–Hochberg method of false discovery rate (FDR).32 When performing multiple comparisons, there is a chance of mistakenly identifying statistically significant relationships because some fraction of discoveries are false. The FDR therefore conceptualizes these type I errors for multiple comparisons as done here using the t-test. In summary, all comparisons are to test for similarity to values at pH 6.5, which represents 0% change in fluorescence intensity.
To assess whether the exceptionally high pH sensitivity of fluorescence in the Nordic Reservoir sample was affected by ionic strength, Fint for the peaks or the value of the index in the ionic strength amended aliquots was compared to an aliquot at the reference pH of 6.5 and the ambient ionic strength of 0 mM. Again, any values across the full pH range (3.5 to 9.0) that were statistically similar to the reference pH of 6.5 and ionic strength of 0 mM were identified using a paired comparison of means (Student's t-test; p < 0.05) with the p-values corrected for multiple testing using the Benjamini–Hochberg method.
Results and discussion
pH sensitivity of humic-like peaks
For natural water samples, the difference in mean fluorescence intensity for humic-like peaks A, C and M remained largely within ±5% of the reference pH of 6.5, with two exceptions for peak A, (i) when the pH was <4.5 for Ängbybadet (Fig. 1a), and (ii) when the pH was <4.0 for Strandsjön (Fig. 1c). From the paired t-test comparison, fluorescence intensity of humic-like DOM in Strandsjön was not statistically different from the reference pH of 6.5 across the full pH range (3.5 to 9.0), while Ängbybadet and Krycksjön had narrower ranges whereby peak intensities were generally stable within a range of 5.0 to 7.0, with only a few exceptions (Table 1). Strandsjön had notably more variability in fluorescence intensities relative to the other samples, despite being handled like all the other samples, particularly so for peak A, with some erratic readings at pH 5.5, 8.0 and 9.0 (Fig. 1c). Likewise, there was greater variability in the background reading of the 10 Strandsjön replicates (ESI Table 2†), suggesting that the high variability in fluorescence was related to inherent sample characteristics, and possible interference from inorganic compounds that absorb at low excitation wavelengths (e.g. nitrate33 or iron34). Despite this erratic fluorescence, mean values for Strandsjön were generally within 5% of the 6.5 reference across the pH range for all five peaks, with the exception of peak A at pH 3.5 (Fig. 1c and g). In general, there was a systematic response for humic-like peaks across the pH range, with a quenching effect at extremely low pH (i.e. <4.0), and a slight increase at higher pH values, even if not statistically different from the reference pH. In the case of the extracted Nordic Reservoir samples, humic-like peaks A, C and M had reduced intensity at a slightly lower pH than the other samples (i.e. <5.0) and were stable across a pH range of 5.0 to 9.0 (Fig. 1d). The general increase in Fint with pH has been observed previously, even in earlier studies before well-known peaks associated with DOM (e.g. A, C, M, T and B) were defined, and when studies tended to focus on extracted humic substances rather than natural waters.18,25 The study by Mobed et al. (1996) declared significant pH effects based on experiments at three pH values (2, 6 and 10) which included acidic and basic conditions lying beyond the range of the current study (3.5 to 9.0). Pullin and Cabaniss et al. (1995) also focused on six extracted humic substances and made an interesting contribution by revealing non-uniform attenuation with lowering pH (from 10.5 to 3.0 in steps of 1–1.5 pH units) across a diagonal slice of the EEM spectra, using a wavelength offset of 20 nm (i.e. λem = λex + 20 nm). Since studies have progressively become inclined to use natural water samples, and test the effects of pH at narrower pH intervals of 1.0,8,20 they have obtained similar findings as we find here, whereby we find that pH has a limited effect within the range typical of natural waters (5.5 and 7.5, 10–90% percentiles, respectively, for Swedish lakes, ESI Fig. 2†). In fact, the pH distribution across Swedish freshwaters indicated that the vast majority (80%) of lakes lie within a range of two pH units around a median value of 6.8, with the upper 90% and lower 10% percentile having a pH of 7.5 and 5.5 (ESI Fig. 2†). Thus, our results confirm a similar conclusion expressing that little changes in spectrophotometric parameters are found across a pH range typical of inland waters.35 | ||
| Fig. 1 Percent change in fluorescence intensity relative to the reference pH 6.5 for three natural lakes (Ängbybadet, Krycksjön and Strandsjön) and one commercial standard (Nordic Reservoir), for three humic-like peaks (peaks A, C and M) (a)–(d) and two protein-like peaks (peaks T and B) (e)–(h). Dashed lines represent ±5% of the reference pH 6.5 value. Grey areas indicate the pH range of 5.5 to 7.5, and filled triangles show ambient pH. | ||

|
pH sensitivity of protein-like peaks
For protein-like peaks, a narrower range of pH stability was observed based on two assessment approaches used here, using the 5% deviation from the 6.5 reference value (Fig. 1) or the paired t-test approach with 6.5 reference pH (Table 1). For the extracted Nordic Reservoir sample, the protein-like peaks were especially sensitive to pH changes (Fig. 1h and Table 1), such that the fluorescence intensity dropped to half the reference value when the pH was <4.5 (Fig. 1h), and was only considered stable within a narrow 0.5 pH range from 6.5 to 7.0. The protein-like peaks of two natural waters, Ängbybadet and Strandsjön, were also attenuated, but to a lesser degree when the pH was lower than 5.5 (Fig. 1e and g), while the protein-like peaks for Krycksjön remained stable across a wider pH range from 4.0 to 8.5 (Fig. 1f and Table 1).The background intensities of peaks T and B were extremely low, or not detected, under ambient conditions for all test samples, and thus we experimentally added an algal extract to ensure we could test effects of pH on protein-like fluorescence. Therefore, pH sensitivity of protein-like peaks might be expected to be similar across samples because the algal material of the same origin was added in a consistent way for all samples, however interactions between the algal DOM and ambient DOM along with other ions in solution may have contributed to quenching effects and resulted in differing responses. The algal extract was made of Scenedesmus, which is a very common algae found anywhere in the world,26 is an important dietary source for freshwater crustaceans36 and is typical of eutrophic systems, and thus is often used as an indicator species.37 Thus, depending on the lake, Scenedesmus could be more or less representative of the local community. As Scenedesmus is culturable, it been used as part of a universal inoculum designed for DOM degradation studies.38 For these reasons, Scenedesmus was considered a good option for generating protein-like fluorescence. The pH sensitivity of tryptophan-like fluorescence observed here mimicked that of the pure amino acid (λex 280 nm; λem 300–350 nm). In fact, the fluorescence of pure tryptophan decreases due to proton quenching of the indole functional group, whereby the addition of a H+ ion reduces the electron density in the benzene ring.21 The reverse is true when the pH increases, whereby more basic conditions result in enhanced fluorescence intensity due to deprotonation of the amino group and increased electron density.21 Tryptophan has a higher quantum yield compared to tyrosine and even phenylalanine, and it is usually the dominant protein-like peak observed in natural waters. In Swedish surface waters, protein-like fluorescence is typically found at low intensities (<10%) relative to humic-like peaks, and often altogether absent,39 and thus the addition of an algal extract was useful to study the pH effects on tryptophan and tyrosine-like fluorescence here. The typically low intensity of protein-like peaks may explain why many previous studies have exclusively focused on the pH sensitivity of humic-like peaks.18,20 One study including the interpretation of tryptophan-like fluorescence found some unpredictable results, but acidic samples generally lost intensity in this area.19
Ionic strength does not affect pH sensitivity
A noteworthy observation here is the particularly high sensitivity of protein-like fluorescence for the Nordic Reservoir sample, which is intriguingly stronger than the natural water samples and suggest differing inter-molecular interactions with ambient humic-like DOM, or other matrix effects. In particular, the Nordic Reservoir humic substance is prepared by the IHSS for commercial purposes by initially concentrating the water sample using reverse osmosis,40 which includes the use of a cation exchange resin to reduce calcium, aluminium and iron that could otherwise cause fouling of the membrane by precipitation. In this concentration process, there is a chance of losing less soluble organic matter to precipitation. Accordingly, the far lower ionic composition and modified molecular composition of DOM in the solution of reconstituted Nordic Reservoir is not representative of natural lake water. If it is true that the lack of background ions was a contributing factor, we would expect to see an influence of ionic strength on the Fint of the Nordic Reservoir sample. Thus, we conducted an additional study to test the effect of ionic strength across four concentrations expected across inland waters ranging from 0 to 0.002 M. We found remarkably consistent intensities across all peaks, with very little effect of ionic strength (Fig. 2 and ESI Table 3†).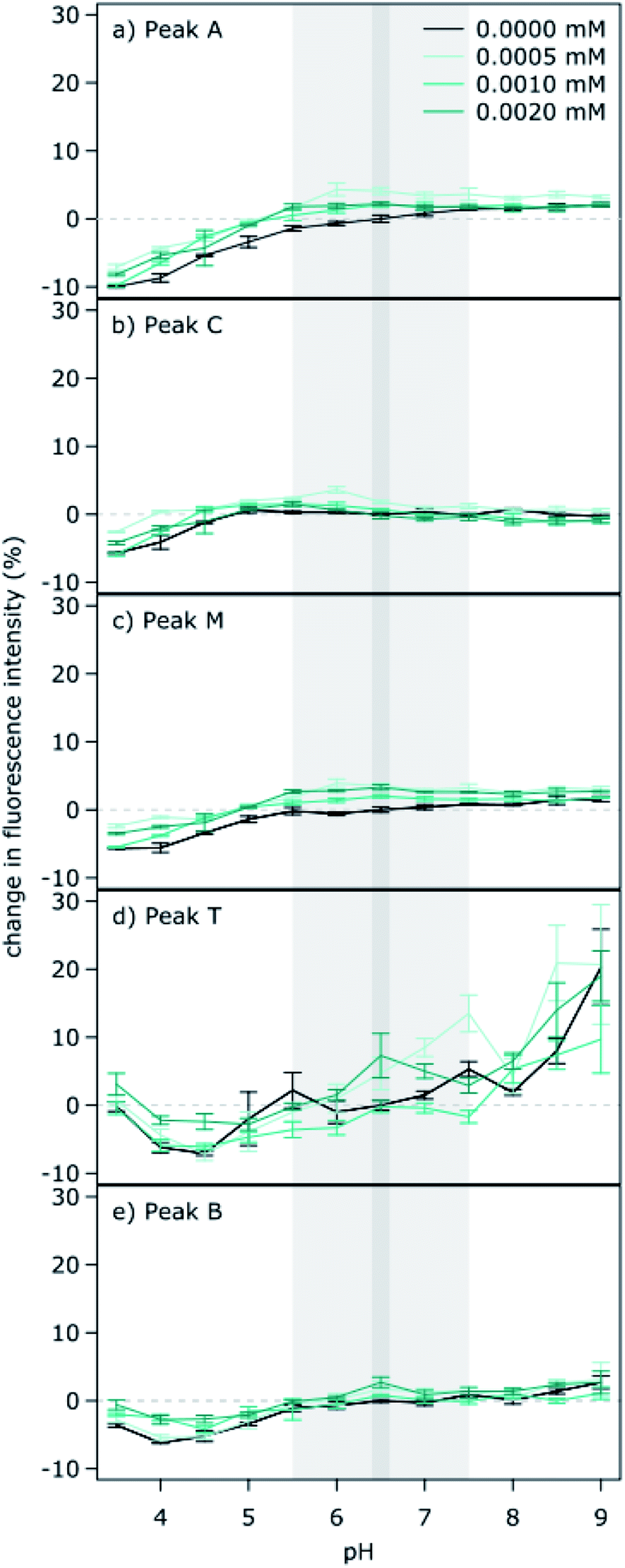 | ||
| Fig. 2 The influence of ionic strength on the fluorescence intensity of different regions of the spectra (a to e, Peaks A, C, M, T and B) for the Nordic Reservoir sample across a pH range from 3.5 to 9.0. All values are expressed as a percent deviation from the reference pH of 6.5 at an ionic strength of 0 mM. | ||
In particular, we found no directional shift in fluorescence intensity with increasing ionic strength. The lack of ionic strength effects (using KCl to modify ionic strength) has been previously reported by Mobed et al. (1996), and we confirm that ionic strength (using a mixture of NaCl, CaCl2 and K2SO4) was not an influencing factor for the strong pH sensitivity observed here for the Nordic Reservoir sample. Other studies have also noted that changes in ionic strength appear to have less effect on fluorescence than pH.41 However, we did not specifically aim to test the influence of ionic strength on DOM fluorescence, rather, we simply rule out that it was the main factor resulting in high sensitivity of protein-like peaks for the Nordic standard here. Others have noted decreases in fluorescence due to ionic strength conditions at higher levels than tested here which may be more relevant for soil solutions, estuaries and marine systems (0.001 to 0.1 M).42
However, it is possible that other ions such as metal ions (not tested here) could modify ionic strength and interact with pH to affect DOM fluorescence. For example, iron speciation and solubility are affected by pH, and iron is known to affect DOM fluorescence.43 This suggests a possible interaction between iron concentration, pH and DOM fluorescence. Although Poulin et al. (2014) measured lower fluorescence intensity for a commercial humic standard at low pH, quenching by iron was more pronounced under mildly acidic conditions than under highly acidic conditions, indicating that iron concentrations might not be the main factor influencing DOM fluorescence at low pH.
The ionic strength experiment conducted on the Nordic Reservoir sample was carried out as a supplemental experiment after recognizing the particularly strong pH effects. Consequently, the concentration of algal extract added to the Nordic Reservoir sample in the original pH experiment differed from the ionic strength experiment due to difficulties in solubilizing DOM from the original algal extract in an exactly consistent manner. Thus, while the pH sensitivity of protein-like peaks is observed for both the pH and ionic strength experiments, the percent change in fluorescence intensity differs between the original pH experiment (Fig. 1h) and subsequent ionic strength experiment (Fig. 2d and e).
Non-uniform pH sensitivity across EEM spectra
Based on the differing sensitivities observed between the commonly identified fluorescing regions (peaks A, C, M, T and B), we explored if there were in fact any consistent patterns in terms of pH sensitivity across the full EEM (Fig. 3). To do this, we focused on the narrower pH range (7.5 to 5.5) most relevant to boreal freshwaters (ESI Fig. 2†), and plotted the % change in fluorescence intensity due to a drop in pH from 7.5 to 5.5, based on a mean of triplicates (Fig. 3). Accordingly, we found distinct regions of the EEM displaying quenching effects and increases in fluorescence intensity with acidification from 7.5 to 5.5. Quenching was observed in the protein-like region (λex/λem = 280 nm/320 nm) as noted for peaks T and B, but more unexpectedly, we also observed quenching along the shoulder of peak C with longer excitation wavelengths (λex/λem = 420 nm/450 nm). Interpretation of pH effects on protein-like peaks assumes that fluorophores represent structures in the corresponding amino acid, either tryptophan or tyrosine. However, the interpretation of pH effects across humic-like regions is more complex because there are multiple underlying fluorophores that could individually respond distinctly and the observed net change is possibly a result of both quenching and enhanced fluorescence. Fluorescence across the humic-like region often includes fluorophores with two excitation peaks, as commonly observed for fluorophores in this region as well as PARAFAC analysis.44 Accordingly, fluorescence in the peak A region is not entirely independent of fluorescence in the peak C and M regions. This combination of factors makes the interpretation of pH effects on underlying humic-like fluorophores more complex than protein-like fluorescence. | ||
| Fig. 3 Illustration of non-uniform changes in fluorescence intensity due to a decrease in pH from 7.5 to 5.5 across the full excitation-emission matrix of three natural water samples (a–c) and one commercial humic standard (d). For reference, the location of Peaks A, C, M, T and B are marked in panel (a), and the location of fluorescence index (FI), freshness index (Fresh) and humification index (HIX) are marked on panel (b). | ||
pH sensitivity of three commonly used fluorescence indices
In the case of the three fluorescence indices, which are ratios of intensities across different regions of the EEM (Fig. 4), each index was plotted on it's own scale, because it is more meaningful to present shifts in actual values, rather than percent change from a reference value as done for the peaks. The fluorescence index (FI) varied across the four samples and remained fairly robust across the pH range, with the Nordic Reservoir sample expressing a maximum change of 0.1 units across the full pH range (Fig. 4a). When the FI was originally developed, it was recommended that individual studies test the influence of pH on the fluorescence index for possible quenching effects and acidify samples if necessary to ensure all samples are at the same pH.14 Our results would show that such a test might not be necessary, especially when the samples are in the pH range 5.5 to 7.5. The freshness index (Fresh) was especially robust to pH changes (Fig. 4b), with changes not exceeding 0.1 units across the studied pH range, while the humification index was more unstable, especially at a pH below 4.5 (Fig. 4c). The changes observed in the FI and Fresh across the full pH range are so small that they would not confound the interpretation with regard to the source of the DOM. For the HIX, more care should be taken when interpreting results from across a pH range, as the same DOM will give slightly different values for this index when measured at different pH values. | ||
| Fig. 4 Distribution of three fluorescence indices (a) fluorescence index, (b) freshness index and (c) humification index across the pH range 3.5 to 9.0 for three natural lakes (Ängbybadet, Strandsjön and Krycksjön) and one commercial reference humic substance (Nordic Reservoir). The grey bar shows the reference pH value of 6.5. | ||
Conclusions and recommendations
- Fluorescence intensity of the five main peaks was stable within the pH range of 5.5 to 7.5, which is common for natural water samples from inland waters, but may vary outside of this range. We recommend that pH is always reported, with particular emphasis placed on the maximum pH range across the dataset (i.e. all samples ranged from _._ to _._ pH units). If samples within a dataset are in this range and within 1.0 pH unit of each other, this point can be noted to alleviate concerns about pH interferences.- Humic-like peaks (peaks A, C and M) were robust to pH changes in the 5.5 to 7.5 range. One sample was robust to pH effects across the full pH range tested (3.5 to 9.0; Strandsjön). In contrast, when the pH of one sample (Ängbybadet) was adjusted to <4.0, a decrease of up to 55% in fluorescence was observed. Since the dataset used here was limited to three natural water samples, we encourage follow up studies including a wider range of samples from across ecosystems.
- Protein-like peaks (peaks T and B) were robust to pH changes in the 5.5 to 7.5 range, with peak T having a wider range of stability than peak B. For instance, two natural samples had a range of stability spanning from 4.0 to 9.0 for peak T.
- The commercially available Nordic Reservoir Humic Substance was highly sensitive to pH changes. The protein-like peaks decreased in fluorescence even within 0.5 pH units of the reference 6.5. The most extreme loss in fluorescence was observed at pH 4.0 for peak T, when a loss of 60% fluorescence was observed.
- The most commonly used indices seem to be robust across a pH range from 5.5 to 7.5. FI and Fresh are stable in the pH range 3.5–9, while HIX shows more variability across this wider pH range.
- If natural pH values are within the pH range of 5.5 to 7.5, we recommend analysis at ambient pH.
- If natural pH values fall outside the 4.5 to 8.5 range, we recommended adjusting the pH (refer to the methods section for details), or alternatively acidifying all samples (in accordance with McKnight et al. 2001), which would account for potential interferences due to metals like iron (Poulin et al. 2014).
- Commercial HS or DOM concentrates, particularly those obtained by reverse osmosis are likely to be more sensitive to pH changes. This should be considered when designing experiments, including samples with poor buffering capacity. We recommend that DOM solutions made with commercial substances or concentrates are prepared at intermediate pH (i.e. pH of 6.5) to facilitate comparison with natural samples.
- Ionic strength within the range of 0 to 0.002 M does not seem to influence the way DOM fluorescence varies across the pH range. Larger differences in ionic strength may be observed in other ecosystems such as soils, estuaries and marine systems thereby need further consideration, particularly if metal ions are included.
- Evaluation of the full EEM when changing the pH from 7.5 to 5.5 showed a decrease in fluorescence intensity in the protein-like region and along the shoulder of peak C with longer excitation wavelengths (Fig. 3). Variation in this region in samples of varying pH is not likely to affect the peaks and indices, but may affect the results of any identified components in this region as commonly done when using parallel factor analysis (PARAFAC). We therefore recommend that care must be taken when interpreting PARAFAC results pertaining to this region, even when the sample pH falls within the 5.5–7.5 range.
Author contributions
MG: investigation, data curation, formal analysis, methodology, visualization, writing-original draft, writing-review & editing. NC: conceptualization, writing-original draft, writing-review & editing. KE: investigation, writing-review & editing. AGB: resources, writing-review & editing. DNK: conceptualization, formal analysis, methodology, data curation, supervision, funding acquisition, project administration, visualization, writing-original draft, writing-review & editing.Conflicts of interest
The authors declare there are no conflicts of interest.Acknowledgements
This work was supported by funding from the Swedish National Science Foundation (D. N. K.) Vetenskapsrådet starting grant (VR 2016-04108), open call (VR 2020-03249), the Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning (FORMAS 201800778), an early career research award from King Carl XVI Gustaf of Sweden, a Marie Sklodowska Curie Action of the European Commission to N. C. (CHROME – 839709) and the Knut and Alice Wallenberg Foundation (Wallenberg Scholar KAW 2018-0191). We would like to thank Christoffer Bergvall for lab support and Karla Münzner for discussions regarding algae.References
- P. G. Coble, S. A. Green, N. V. Blough and R. B. Gagosian, Nature, 1990, 348, 432–435 CrossRef CAS.
- C. A. Stedmon, S. Markager and R. Bro, Mar. Chem., 2003, 82, 239–254 CrossRef CAS.
- J. B. Fellman, E. Hood and R. G. M. Spencer, Limnol. Oceanogr., 2010, 55, 2452–2462 CrossRef CAS.
- K. R. Murphy, C. A. Stedmon, P. Wenig and R. Bro, Anal. Methods, 2014, 6, 658–661 RSC.
- U. J. Wunsch, R. Bro, C. A. Stedmon, P. Wenig and K. R. Murphy, Anal. Methods, 2019, 11, 888–893 RSC.
- N. Senesi, Anal. Chim. Acta, 1990, 232, 77–106 CrossRef CAS.
- R. W. P. M. Lane, Mar. Chem., 1982, 11, 395–401 CrossRef.
- R. G. M. Spencer, L. Bolton and A. Baker, Water Res., 2007, 41, 2941–2950 CrossRef CAS PubMed.
- A. Hartland, I. J. Fairchild, J. R. Lead and A. Baker, Sci. Total Environ., 2010, 408, 5940–5950 CrossRef CAS PubMed.
- J. R. Lakowicz, Principles of Fluorescence Spectroscopy, Springer-Verlag, New York, USA, 3rd edn, 2006 Search PubMed.
- N. Senesi, T. M. Miano, M. R. Provenzano and G. Brunetti, Soil Sci., 1991, 152, 259–271 CrossRef CAS.
- M. L. Pace, I. Reche, J. J. Cole, A. Fernandez-Barbero, I. P. Mazuecos and Y. T. Prairie, Biogeochemistry, 2012, 108, 109–118 CrossRef CAS.
- Y. Gao, M. Q. Yan and G. V. Korshin, Environ. Sci. Technol., 2015, 49, 5905–5912 CrossRef CAS PubMed.
- D. M. McKnight, E. W. Boyer, P. K. Westerhoff, P. T. Doran, T. Kulbe and D. T. Andersen, Limnol. Oceanogr., 2001, 46, 38–48 CrossRef CAS.
- R. M. Cory, M. P. Miller, D. M. McKnight, J. J. Guerard and P. L. Miller, Limnol. Oceanogr.: Methods, 2010, 8, 67–78 CAS.
- E. Parlanti, K. Worz, L. Geoffroy and M. Lamotte, Org. Geochem., 2000, 31, 1765–1781 CrossRef CAS.
- A. Zsolnay, E. Baigar, M. Jimenez, B. Steinweg and F. Saccomandi, Chemosphere, 1999, 38, 45–50 CrossRef CAS PubMed.
- J. J. Mobed, S. L. Hemmingsen, J. L. Autry and L. B. McGown, Environ. Sci. Technol., 1996, 30, 3061–3065 CrossRef CAS.
- A. Baker, S. Elliott and J. R. Lead, Chemosphere, 2007, 67, 2035–2043 CrossRef CAS PubMed.
- N. Patel-Sorrentino, S. Mounier and J. Y. Benaim, Water Res., 2002, 36, 2571–2581 CrossRef CAS.
- F. Sun, W. Zong, R. Liu, J. Chai and Y. Liu, Spectrochim. Acta, Part A, 2010, 76, 142–145 CrossRef PubMed.
- K. R. Murphy, C. A. Stedmon, T. D. Waite and G. M. Ruiz, Mar. Chem., 2008, 108, 40–58 CrossRef CAS.
- C. A. Stedmon, B. Seredynska-Sobecka, R. Boe-Hansen, N. Le Tallec, C. K. Waul and E. Arvin, Water Res., 2011, 45, 6030–6038 CrossRef CAS PubMed.
- M. J. Pullin and S. E. Cabaniss, Environ. Sci. Technol., 1995, 29, 1460–1467 CrossRef CAS PubMed.
- M. J. Pullin and S. E. Cabaniss, Geochim. Cosmochim. Acta, 2003, 67, 4067–4077 CrossRef CAS.
- P. F. M. Coesel and L. Krienitz, Biodiversity and Conservation, 2008, 17, 381–392 CrossRef.
- J. Kotai, Instructions for preparation of modified nutrient solution Z8 for algae, 1972 Search PubMed.
- A. Lawaetz and C. Stedmon, Appl. Spectrosc., 2009, 63, 936–940 CrossRef CAS PubMed.
- D. N. Kothawala, K. R. Murphy, C. A. Stedmon, G. A. Weyhenmeyer and L. J. Tranvik, Limnol. Oceanogr.: Methods, 2013, 11, 616–630 CrossRef.
- R. M. Cory, D. M. McKnight, Y. P. Chin, P. Miller and C. L. Jaros, J. Geophys. Res.: Biogeosci., 2007, 112, G04S51 Search PubMed.
- J. Fölster, R. K. Johnson, M. N. Futter and A. Wilander, Ambio, 2014, 43, 3–18 CrossRef PubMed.
- Y. Benjamini and Y. Hochberg, J. Roy. Stat. Soc. B Stat. Meth., 1995, 57, 289–300 Search PubMed.
- J. E. Birdwell and A. S. Engel, J. Cave Karst Stud., 2009, 71, 144–156 CAS.
- J. L. Weishaar, G. R. Aiken, B. A. Bergamaschi, M. S. Fram, R. Fujii and K. Mopper, Environ. Sci. Technol., 2003, 37, 4702–4708 CrossRef CAS PubMed.
- R. G. M. Spencer and P. Coble, in Aquatic organic matter fluorescence, ed. P. Coble, J. Lead, A. Baker, D. M. Renyolds and R. G. M. Spencer, Cambridge University Press, New York, 2014 Search PubMed.
- C. L. Wang, X. L. Chang, Q. Y. Shi and X. Zhang, Environ. Sci. Technol., 2018, 52, 12133–12141 CrossRef CAS PubMed.
- G. Phillips, A. Lyche-Solheim, B. Skjelbred, U. Mischke, S. Drakare, G. Free, M. Jarvinen, C. de Hoyos, G. Morabito, S. Poikane and L. Carvalho, Hydrobiologia, 2013, 704, 75–95 CrossRef.
- A. Pastor, N. Catalan, N. Nagar, T. Light, C. M. Borrego and R. Marce, Limnol. Oceanogr.: Methods, 2018, 16, 421–433 CrossRef CAS.
- D. N. Kothawala, C. A. Stedmon, R. A. Muller, G. A. Weyhenmeyer, S. J. Kohler and L. J. Tranvik, Global Change Biology, 2014, 20, 1101–1114 CrossRef PubMed.
- L. Sun, E. M. Perdue and J. F. McCarthy, Water Res., 1995, 29, 1471–1477 CrossRef CAS.
- C. L. Osburn, R. Del Vecchio and T. J. Boyd, in Aquatic organic matter fluorescence, ed. P. Coble, J. Lead, A. Baker, D. M. Renyolds and R. G. M. Spencer, Cambridge University Press, New York, 2014 Search PubMed.
- K. Ghosh and M. Schnitzer, J. Soil Sci., 1979, 30, 735–745 CrossRef CAS.
- B. A. Poulin, J. N. Ryan and G. R. Aiken, Environ. Sci. Technol., 2014, 48, 10098–10106 CrossRef CAS PubMed.
- K. R. Murphy, C. A. Stedmon, D. Graeber and R. Bro, Anal. Methods, 2013, 5, 6557–6566 RSC.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d1ay01702k |
| This journal is © The Royal Society of Chemistry 2022 |