
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Glutathione sensitive vesicles prepared from supramolecular amphiphiles†
Emily B.
Mobley
,
Natalie
Byrd
,
Matthew G.
Yim
,
Rachel
Gariepy
,
Maya
Rieder
and
Sandra
Ward
 *
*
Department of Chemistry and Biochemistry, California Polytechnic State University, 1 Grand Avenue, San Luis Obispo, California 93401, USA. E-mail: sward07@calpoly.edu
First published on 27th September 2021
Abstract
Glutathione (GSH) sensitive vesicles were prepared by the self-assembly of amphiphilic inclusion complexes. These novel chemically sensitive supramolecular amphiphiles are anticipated to have applications in drug delivery; the nanocarriers can protect the encapsulated cargo and release it via triggered degradation in high concentrations of GSH. Additionally, the sensitivity of the vesicles to GSH indicates that the dynamic covalent disulfide bond at the vesicle surface can be used for post-modification of the nanocarrier via a thiol–disulfide exchange, a strategy that can be exploited to introduce targeting moieties to increase treatment specificity. Supramolecular amphiphiles containing a dynamic covalent disulfide bond were prepared via the host–guest inclusion complexes between alkylated β-cyclodextrin (β-CD) hosts and adamantane terminated polyethylene glycol derivatives. The significant difference between the critical micelle concentrations of the supramolecular amphiphiles and the individual host and guest components confirmed that a unique supramolecular amphiphile was formed. Fluorescence experiments and dynamic light scattering (DLS) revealed that the supramolecular amphiphiles self-assembled into vesicles of 130 nm diameter which were stable for 8 months. Degradation of the vesicles after incubation with GSH was monitored using DLS and by the release of encapsulated 5,6-carboxyfluorescein (CF), observed by an increase in fluorescence intensity. Degradation of the nanocarrier was faster at intracellular GSH concentrations than at extracellular GSH concentrations.
Introduction
In aqueous solutions, amphiphiles can act as molecular building blocks and spontaneously self-assemble into nanostructures such as nanotubes, micelles, and vesicles. For drug delivery purposes, cargo such as drugs, proteins or other therapeutics can be encapsulated in the aqueous interiors or the lipophilic bilayer of the nanocarriers for controlled and sustained release. Nanocarriers are designed to degrade and release the therapeutics at the intended site of delivery by incorporating chemically sensitive bonds that respond to changes in pH or redox conditions. Synthesizing amphiphiles that incorporate design features such as chemically sensitive bonds or targeting moieties can add a significant synthetic burden by increasing the number of both the synthetic and tedious purification steps. Supramolecular chemistry can be exploited to simplify amphiphile synthesis by the spontaneous self-assembly of two or more molecular components through non-covalent interactions.Host–guest interactions are frequently employed as a driving force in the self-assembly of supramolecular amphiphiles.1,2 In aqueous solutions, small hydrophobic guest molecules are driven into the cavity of macrocyclic hosts by the entropically favoured hydrophobic effect.3–5 Vesicle nanostructures have been prepared from inclusion complexes with complexes with pillar[6]arenes,6–15 calix[4]arenes,16–21 cyclodextrins,22–35 cucurbit[6]urils35–38 and resorcin[4]arenes39 as the macrocyclic hosts. Triggered degradation and release of the encapsulated cargo have been achieved with supramolecular amphiphiles by changing the pH,6,8,10,12–15,33,39–41 or redox conditions.7–9,30,33,42
Degradation of supramolecular amphiphile vesicles can be triggered by forming or breaking bonds within the guest in the host–guest inclusion complex. For example, Wang et al. reported a glucose sensitive supramolecular vesicle that released insulin at high glucose concentrations to treat diabetes. The vesicle degraded when the hydrophobic/hydrophilic balance (HLB) of the inclusion complex was disrupted upon glucose binding to the pyridylboronic acid derivative guest.11 On the other hand, Lie et al., designed a supramolecular amphiphile in which the aliphatic chain of the guest was cleaved. Butyrlcholinestrase cleaved the ester bond of myristoylcholine, causing the vesicle to disassemble, again due to the disruption of the HLB of the supramolecular amphiphile.18
Glutathione can trigger release from drug delivery systems with disulfide bonds. Glutathione is a naturally abundant antioxidant that has significantly different intracellular (1–10 mM) and extracellular (2–20 μM) concentrations.43,44 The tripeptide has a cysteine residue with a free thiol that can undergo thiol–disulfide exchange to cleave disulfide bonds. Zhang et al. prepared micelles from a supramolecular amphiphile that covalently bound a therapeutic porphyrin to the guest molecule via a disulfide bond.45,46 Cleavage of the disulfide bond by GSH released the porphyrin. This system was limited to the delivery of porphyrins, and it would require to synthesize and characterize a new supramolecular amphiphile to deliver other desired therapeutics.
Here, we present a novel supramolecular amphiphile that self-assembles via non-covalent host–guest interactions to form glutathione responsive vesicles. This versatile, supramolecular amphiphile can be used to deliver either hydrophilic or hydrophobic therapeutic agents by encapsulating them into the aqueous vesicle interior or embedding them in the hydrophobic bilayer. Synthesis of the amphiphile building blocks was simplified by combining the hydrophobic thioalkylated β-CD hosts and hydrophilic adamantyl poly(ethylene glycol) (PEG) guests to form an amphiphilic inclusion complex (Fig. 1). A disulfide bond in the pegylated adamantyl guests was designed to cleave in the presence of GSH. Upon cleavage of the PEG tail on the supramolecular amphiphile, the disruption of the hydrophilic/hydrophobic balance caused degradation of the vesicle (Fig. 1). The supramolecular amphiphiles were confirmed and characterized by measuring the critical micelle concentration (CMC). The vesicles were shown to be remarkably stable (shelf-life of 8-months); however, they readily degraded in the presence of GSH. Degradation studies showed the vesicles were resistant to degradation at extracellular GSH concentrations but readily degraded when incubated with intracellular concentrations.
 | ||
| Fig. 1 (A) Schematic illustration of the inclusion complexes that form supramolecular amphiphiles and self-assemble into vesicles. Glutathione undergoes a thiol–disulfide exchange with the AdSSPEG guest, and cleavage of the hydrophilic PEG disrupts the hydrophilic/hydrophobic balance causing the vesicle to disassemble. (B) Structures of alkylated cyclodextrin hosts CD12 and CD14, and pegylated adamantyl guests AdSSPEG–OH and AdSSPEG–OMe. | ||
Results and discussion
The thioalkylated β-CD derivative hosts (Scheme 1) were synthesized via a synthetic route previously reported in the literature.47 A bromine was installed on the 6-position of a native cyclodextrin using an Appel reaction. A subsequent SN2 reaction with an appropriate thiolate yielded thioalkyl β-CD. | ||
| Scheme 1 Synthesis of CD12 and CD14 from native β-CD by an Appel reaction (80% yield) followed by an SN2 substitution with an appropriate thiolate (70–90% yield). | ||
The final host compound was isolated by precipitation in water. The 1H NMR confirmed the complete substitution on all seven glucose units by the single set of glucose signals from the symmetry of the compound. A triplet at 2.8 ppm confirmed the thioether linkage.
The synthesis of AdSSPEG–OH and AdSSPEG–OMe was adapted from the synthesis of asymmetric functionalization of 1,3-dithiopropanoic acid (Scheme 2) mentioned in previous reports.48,49 1,3-dithiopropanoic acid was cyclized into an anhydride using acetyl chloride. The cyclic anhydride was opened by a nucleophilic attack from PEG1000 or the corresponding methyl-ether PEG. The free carboxylic acid was coupled with adamantol using N,N′-dicyclocarbodiimide (DCC) as the coupling agent.
 | ||
| Scheme 2 AdSSPEG–OH and AdSSPEG–OMe were synthesized from 1,3,-dipropanoic acid. After cyclization into an anhydride, the ring was opened by either a methyl-ether or OH terminated PEG (MWavg = 1000 g mol−1), and finally 1-adamantol underwent an ester coupling using DCC. | ||
Supramolecular amphiphile preparation and critical micelle concentration
The supramolecular amphiphiles self-assembled into an inclusion complex by combining a host (CD12 or CD14) with a guest (AdSSPEG–OH or AdSSPEG–OMe) in a 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1 molar ratio in water. The critical micelle concentration (CMC) was determined by fluorescence spectroscopy using Nile red as the hydrophobic probe.
:1 molar ratio in water. The critical micelle concentration (CMC) was determined by fluorescence spectroscopy using Nile red as the hydrophobic probe.
The four unique supramolecular amphiphiles had CMCs in the range of 27.2–37.9 μM (Table 1, Fig. S1–S4, ESI†). The supramolecular amphiphiles formed using the CD14 host had lower CMCs than the supramolecular amphiphiles prepared using CD12 due to the increase in the hydrophobic alkyl chains. Supramolecular amphiphiles with AdSSPEG–OMe had a lower CMC than supramolecular amphiphiles with AdSSPEG–OH, which was attributed to the decrease in hydrogen bonding ability.
| Sample | CMC (μM) | Hydrodynamic radius (nm) | Δ % Fluorescenceb |
|---|---|---|---|
| a CMC not measured because of poor water solubility. b Change in fluorescence after addition of Triton-X. | |||
| CD12 ⊂ AdSSPEG–OMe | 30.5 | 55 ± 2 | 2203 |
| CD14 ⊂ AdSSPEG–OMe | 27.2 | 67 ± 1 | 1875 |
| CD12 ⊂ AdSSPEG–OH | 37.9 | 62 ± 1 | 2071 |
| CD14 ⊂ AdSSPEG–OH | 35.9 | 58 ± 4 | 1768 |
| AdSSPEG–OMe | 200.5 | — | 96 |
| AdSSPEG–OH | 870.9 | — | 328 |
| CD12 | —a | — | 209 |
| CD14 | —a | — | 370 |
The CMC of AdSSPEG–OMe and AdSSPEG–OH was measured individually to ensure that the aggregation observed was in fact due to a unique supramolecular amphiphile. The CMC of the two CD hosts was not measured because of very poor water solubility. The CMC of AdSSPEG–OMe and AdSSPEG–OH was significantly higher than the CMC of the inclusion complexes, 200.5 μM and 870.9 μM, respectively (Table 1, Fig. S5 and S6, ESI†). The considerably larger CMC of the guest compounds and the inability to measure the CMC of the host compounds confirmed that a supramolecular amphiphile was formed and the observed CMCs (27.2–37.9 μM) were not due to aggregation of the individual host and guest components.
Vesicle preparation and characterization
Vesicles were prepared via the thin film hydration method. Briefly, a thin film of the prepared supramolecular amphiphile inclusion complex was hydrated, sonicated, and extruded through a 200 nm membrane. The vesicles sizes were characterized by dynamic light scattering (DLS) and are summarized in Table 1. Changes in carbon chain length on the CD host or a different terminal functional group on PEG chain did not significantly affect the size or stability of the vesicles. All four supramolecular amphiphiles produced vesicles of consistent particle sizes with diameters between 110 and 134 nm (Table 1).The stability of the vesicles was monitored over time by DLS and showed that the vesicles had a long shelf life of up to 8 months. All four supramolecular amphiphiles formed vesicles that displayed similar stability (Fig. S7–S9, ESI†). Fig. 2A shows the DLS of vesicles formed from CD14 ⊂ AdSSPEG–OMe; the vesicles had a consistent particle size up to 6 months with small micelle peaks at ∼5–10 nm. At 8 months the initial peak significantly broadened and particles smaller than 5 nm were observed.
 | ||
| Fig. 2 (A) The hydrodynamic radius of CD14 ⊂ AdSSPEG–OMe vesicles over time; particles of ∼64 nm were the predominant species in solution, with small amounts of ∼5–10 nm micelles. Increasing polydispersity of the hydrodynamic radius of the vesicles at 8 months indicates that the highly ordered vesicle structure is beginning to break apart. (B) Fluorescence of 50 mM CF filled CD14 ⊂ AdSSPEG–OMe vesicles, where Triton-X was injected at 30 seconds and a significant increase in fluorescence was observed. | ||
Fluorescence experiments confirmed that the particles observed by DLS were vesicles. Fluorescent vesicles were prepared by hydrating a thin film of the supramolecular amphiphiles with a solution of 5,6-carboxyfluorescein (CF) at a self-quenching concentration (50 mM). When the vesicles were lysed by addition of Triton-X, the CF encapsulated in the aqueous interior of the vesicle was released and the dilution in CF caused a 2000% increase of the fluorescence signal (Table 1, Fig. 1B, Fig. S10, ESI†). The individual components of the supramolecular amphiphile (CD12, CD14, AdSSPEGOMe, AdSSPEG–OH, Fig. S11, ESI†) were also prepared according to the same procedure. Upon the addition of Triton-X the increase in fluorescence was 10-fold less, due to the lack of an aqueous interior indicating that the individual components of the supramolecular amphiphile do not self-assemble into vesicles and that the unique supramolecular amphiphiles are responsible for the observed vesicles.
Glutathione sensitivity monitored by dynamic light scattering
Next, the stimuli-responsiveness of the vesicles was examined. The disulfide bonds in the guest molecules were designed to cleave in the presence of the natural glutathione via a thiol–disulfide exchange. The vesicles were treated with 10 mM and 2 μM glutathione to mimic the intra and extracellular environments, respectively. All four supramolecular amphiphile vesicles displayed similar DLS results (Fig. S12–S14, ESI†). Fig. 3 shows the DLS results of CD14 ⊂ AdSSPEG–OMe after incubation with GSH. After 1 hour of incubation in both extra and intracellular GSH concentrations, all four vesicles showed a significant peak broadening on DLS. Samples incubated in 10 mM GSH had large particles (300–800 nm radius) after 1 hour, which were the result of the supramolecular amphiphile being modified by GSH and thereby altering the vesicle nanostructure. Similar observations have been reported by other studies exploring the stimuli triggered degradation of nanoparticles.45,50 | ||
| Fig. 3 Changes in DLS after incubation in (A) 2 μM GSH (extracellular) and (B) 10 mM GSH (intracellular). DLS after 1 hour and 12 hours showed slight changes, with the appearance of larger particles. A visible precipitate was observed after 1 week of incubation in intracellular GSH, precipitation was never observed for vesicles in extracellular GSH. | ||
Samples incubated in 2 μM GSH did not show additional peaks until after 12 hours of incubation. These initial changes indicated that the disulfide bond was responsive to GSH and structure changes occurred in the vesicles.
After one week in 10 mM GSH, all four supramolecular amphiphile vesicles showed a visible precipitate. The precipitate from the CD12 ⊂ AdSSPEG–OMe vesicles was isolated and 1H NMR confirmed that the structure was CD12. The precipitate was the result of the cleavage of the disulfide bond which removed the PEG chain from supramolecular amphiphile rendering CD12 insoluble in water.
Precipitation was never observed for vesicles incubated in extracellular concentration (2 μM GSH), even after several months of monitoring, indicating that the vesicles were much more sensitive to intracellular GSH concentrations compared to extracellular concentrations.
Glutathione sensitivity monitored by fluorescence
Fluorescence studies were performed on the vesicle prepared from the CD14 ⊂ AdSSPEG–OMe supramolecular amphiphile to confirm GSH responsiveness. Although the DLS and fluorescence experiments follow similar trends, the vesicles with encapsulated CF required an additional purification step with a Sephadex column to remove any free CF, which caused dilution of the vesicle and a slower degradation. Vesicles containing a self-quenching concentration of encapsulated CF (50 mM) were prepared and incubated with 2 μM and 10 mM of GSH. These vesicles were monitored and it was found that the fluorescence intensity increased over time due to CF being released as the GSH cleaved the disulfide bond in the supramolecular amphiphile (Fig. 4).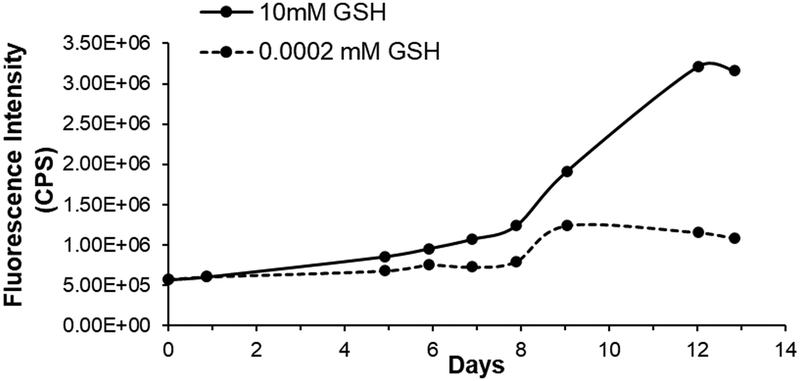 | ||
| Fig. 4 Emission of 50 mM filled CF vesicles incubated in 10 mM and 2 μM glutathione to mimic intracellular and extracellular GSH concentrations, respectively, monitored over two weeks. | ||
Small increases in fluorescence in both intra- and extracellular conditions were observed for the first 8 days. The fluorescence steadily increased after 9 days and levelled off after 13 days. Interestingly, the release of CF was much lower when the vesicles were incubated with the 2 μM extracellular GSH compared to the 10 mM intracellular GSH.
To confirm that maximum release was achieved, Triton-X was added to each sample to lyse the vesicles and release any CF that remained encapsulated at that time point. Fig. 5 shows the percent change in fluorescence (ΔF) calculated using eqn (1), where (If) and (Ii) are the emission intensities before and after the addition of Triton-X, respectively.
| ΔF = [(If – Ii)/(Ii)] × 100% | (1) |
 | ||
| Fig. 5 Δ F monitored over 13 days of 50 mM filled CF vesicles incubated in 10 mM and 2 μM glutathione to mimic intracellular and extracellular GSH concentrations, respectively. | ||
ΔF steadily decreased over time and levelled off after 9 days for vesicles in both the intra and extracellular GSH concentrations. For the intracellular samples, complete release of CF was apparent when ΔF < 0 after 9 days. The addition of Triton-X, after this point, had a dilution effect because there was no longer any vesicle present to lyse and release CF. The vesicles incubated in extracellular GSH concentration never achieved complete release of CF within two weeks of monitoring. It can be concluded that the vesicles have a higher sensitivity to intracellular GSH concentration than extracellular GSH concentration.
Conclusions
In summary, we prepared glutathione sensitive vesicles that have consistent hydrodynamic sizes and are stable up to 8 months. By measuring the CMC and performing fluorescence experiments, we confirmed that the vesicle nanostructure was a result of the self-assembly of supramolecular amphiphiles, rather than aggregation of the individual host and guest components. The degradation of the vesicles occurred faster when incubated at intracellular GSH concentrations than at extracellular GSH concentrations when monitored by DLS or CF release. The resistance of vesicles to degradation at a low extracellular GSH concentration makes them viable nanocarriers for drug delivery; the payload will be encapsulated and protected until exposure to high GSH concentrations. These results confirm that the vesicles are responsive to GSH and can undergo dynamic covalent thiol–disulfide exchange on the surface of the vesicle. Future studies will use this capability to modify the vesicles after fabrication, such as introducing targeting moieties to enhance selectivity.Conflicts of interest
There are no conflicts to declare.Acknowledgements
The authors thank the Bill and Linda Frost Fund, California State University Program for Education and Research in Biotechnology, and the Kenneth N. Edwards Western Coating Technology Center for financial support.References
- G. Yu, K. Jie and F. Huang, Chem. Rev., 2015, 115, 7240–7303 CrossRef CAS PubMed.
- L. García-Rio and N. Basílio, Curr. Opin. Colloid Interface Sci., 2019, 44, 225–237 CrossRef.
- B. Widom, P. Bhimalapuram and K. Koga, Phys. Chem. Chem. Phys., 2003, 5, 3085–3093 RSC.
- N. T. Southall, K. A. Dill and A. D. J. Haymet, J. Phys. Chem. B, 2002, 106, 521–533 CrossRef CAS.
- C. Tanford, The Hydrophobic Effect: Formation of Micelles and Biological Membranes, John Wiley & Sons Inc, New York, 1980 Search PubMed.
- J. Chen, Y. Zhang, Z. Meng, L. Guo, X. Yuan, Y. Zhang, Y. Chai, J. L. Sessler, Q. Meng and C. Li, Chem. Sci., 2020, 11, 6275–6282 RSC.
- T. Xiao, L. Qi, W. Zhong, C. Lin, R. Wang and L. Wang, Mater. Chem. Front., 2019, 3, 1973–1993 RSC.
- Q. Hao, Y. Kang, J. F. Xu and X. Zhang, Langmuir, 2020, 36, 4080–4087 CrossRef CAS PubMed.
- Y. Zhou, K. Jie and F. Huang, Org. Chem. Front., 2017, 4, 2387–2391 RSC.
- X. Liu, K. Jia, Y. Wang, W. Shao, C. Yao, L. Peng, D. Zhang, X. Y. Hu and L. Wang, ACS Appl. Mater. Interfaces, 2017, 9, 4843–4850 CrossRef CAS PubMed.
- L. Gao, T. Wang, K. Jia, X. Wu, C. Yao, W. Shao, D. Zhang, X. Y. Hu and L. Wang, Chem. – Eur. J., 2017, 23, 6605–6614 CrossRef CAS PubMed.
- Y. Cao, X. Y. Hu, Y. Li, X. Zou, S. Xiong, C. Lin, Y. Z. Shen and L. Wang, J. Am. Chem. Soc., 2014, 136, 10762–10769 CrossRef CAS PubMed.
- L.-B.-B Meng, W. Zhang, D. Li, Y. Li, X.-Y. Hu, L. Wang and L. Guigen, Chem. Commun., 2015, 51, 14381–14384 RSC.
- Y. Yao, J. Li, J. Dai, X. Chi and M. Xue, RSC Adv., 2014, 4, 9039–9043 RSC.
- X. Danyu, S. Liqing, X. Min and B. Shio, New J. Chem., 2016, 40, 9890–9894 RSC.
- C. Chen, X. Ni, H. W. Tian, Q. Liu, D. S. Guo and D. Ding, Angew. Chem., Int. Ed., 2020, 59, 10008–10012 CrossRef CAS PubMed.
- I. Pisagatti, L. Barbera, V. Giuseppe, G. Valentina, N. Micali, E. Fazio, M. F. P. Fortunato Neri and A. Nott, New J. Chem., 2019, 43, 7628–7635 RSC.
- D. S. Guo, K. Wang, Y. X. Wang and Y. Liu, J. Am. Chem. Soc., 2012, 134, 10244–10250 CrossRef CAS PubMed.
- V. Francisco, N. Basilio, L. Garcia-Rio, J. R. Leis, E. F. Maques and C. Vázquez-Vázquez, Chem. Commun., 2010, 46, 6551–6553 RSC.
- J. G. Harangozó, V. Wintgens, Z. Miskolczy, J. M. Guigner, C. Amiel and L. Biczók, Langmuir, 2016, 32, 10651–10658 CrossRef PubMed.
- N. Basilio, M. Martiín-Pastor and L. García-Río, Langmuir, 2012, 28, 6561–6568 CrossRef CAS PubMed.
- X. Zhong, C. Hu, X. Yan, D. Zhu, Q. Chen, W. Li, L. Feng and Y. Wei, RSC Adv., 2019, 9, 17281–17290 RSC.
- S. Fu, X. Su, M. Li, S. Song, L. Wang, D. Wang and B. Z. Tang, Adv. Sci., 2020, 7, 1–9 Search PubMed.
- M. Salih, C. A. Omolo, N. Agrawal, P. Walvekar, A. Y. Waddad, C. Mocktar, C. Ramdhin and T. Govender, Int. J. Pharm., 2020, 574, 118881 CrossRef CAS PubMed.
- B. Jing, X. Chen, X. D. Wang, C. J. Yang, Y. Z. Xie and H. Y. Qiu, Chem. – Eur. J., 2007, 13, 9137–9142 CrossRef CAS PubMed.
- Q. Yan, J. Yuan, Z. Cai, Y. Xin, Y. Kang and Y. Yin, Synthesis, 2010, 1–7 Search PubMed.
- Z. Du, K. Ke, X. Chang, R. Dong and B. Ren, Langmuir, 2018, 34, 5606–5614 CrossRef CAS PubMed.
- W. Tao, Y. Liu, B. Jiang, S. Yu, W. Huang, Y. Zhou and D. Yan, J. Am. Chem. Soc., 2012, 134, 762–764 CrossRef CAS PubMed.
- Y. P. Wang, N. Ma, Z. Q. Wang and X. Zhang, Angew. Chem., Int. Ed., 2007, 46, 2823–2826 CrossRef CAS PubMed.
- H. Zou, H. Sun, L. Wang, L. Zhao, J. Li, Z. Dong, Q. Luo, J. Xu and J. Liu, Soft Matter, 2016, 12, 1192–1199 RSC.
- S. K. Albert, H. V. P. Thelu, M. Golla, N. Krishnan and R. Varghese, Nanoscale, 2017, 9, 5425–5432 RSC.
- N. Nayak and K. R. Gopidas, J. Mater. Chem. B, 2015, 3, 3425–3428 RSC.
- H. C. Zhang, Z. N. Liu, F. F. Xin, W. An, A. Y. Hao, J. Y. Li, Y. Y. Li, L. Z. Sun, T. Sun, W. J. Zhao, Y. M. Li and L. Kong, Carbohydr. Res., 2011, 346, 294–304 CrossRef CAS PubMed.
- X. Y. Hu, J. Gao, F. Y. Chen and D. S. Guo, J. Controlled Release, 2020, 324, 124–133 CrossRef CAS PubMed.
- Y. J. Jeon, P. K. Bharadwaj, S. W. Choi, J. W. Lee and K. Kim, Angew. Chem., Int. Ed., 2002, 41, 4474–4476 CrossRef CAS PubMed.
- M. A. Horsch, Z. L. Zhang and S. C. Glotzer, Soft Matter, 2010, 6, 945–954 RSC.
- D. Jiao, J. Geng, X. J. Loh, D. Das, T.-C. Lee and O. A. Scherman, Angew. Chem., 2012, 124, 9771–9775 CrossRef.
- K. M. Park, M. Y. Hur, S. K. Ghosh, D. R. Boraste, S. Kim and K. Kim, Chem. Commun., 2019, 55, 10654–10664 RSC.
- Y. Zhou, K. Jie and Y. Yao, New J. Chem., 2017, 41, 916–919 RSC.
- T. Sun, Y. Li, H. Zhang, J. Li, F. Xin, L. Kong and A. Hao, Colloids Surf., A, 2011, 375, 87–96 CrossRef CAS.
- Z. Zhang, J. Ding, X. Chen, C. Xiao, C. He, X. Zhuang, L. Chen and X. Chen, Polym. Chem., 2013, 4, 3265–3271 RSC.
- W. Yuan, J. Ma, Z. Zhao and S. Liu, Macromol. Rapid Commun., 2020, 41, 5–9 CrossRef PubMed.
- M. Alton and M. E. Anderson, Annu. Rev. Biochem., 1983, 52, 711–760 CrossRef PubMed.
- H. F. Gilbert, Methods Enzymol., 1995, 251, 8–28 CAS.
- F. Liu, Y. Ma, L. Xu, L. Liu and W. Zhang, Biomater. Sci., 2015, 3, 1218–1227 RSC.
- Y. Chen, L. Rui, L. Liu and W. Zhang, Polym. Chem., 2016, 7, 3268–3276 RSC.
- C. C. Ling, S. Ward, O. Calderon, S. N. Keller, P. Zhang and V. Williams, J. Mater. Chem. C, 2014, 2, 4928–4936 RSC.
- L. Jia, Z. Li, D. Zhang, Q. Zhang, J. Shen, H. Guo, X. Tian, G. Liu, D. Zheng and L. Qi, Polym. Chem., 2013, 4, 156–165 RSC.
- P. Zhang, H. Zhang, W. He, D. Zhao, A. Song and Y. Luan, Biomacromolecules, 2016, 17, 1621–1632 CrossRef CAS PubMed.
- L. Xu, H. Wang, H. Tian, M. Zhang, J. He and P. Ni, Polym. Chem., 2021, 12, 2152–2164 RSC.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d1sm01238j |
| This journal is © The Royal Society of Chemistry 2021 |