
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceDistribution characteristics of nitrogen and the related microbial community in the surface sediments of the Songhua River†
Congyu Lia,
Zhen Zhongac,
Wenfu Wangd,
Haiyan Wang *a,
Guokai Yana,
Weiyang Donga,
Zhaosheng Chu*b,
Huan Wanga and
Yang Changa
*a,
Guokai Yana,
Weiyang Donga,
Zhaosheng Chu*b,
Huan Wanga and
Yang Changa
aResearch Center of Environmental Pollution Control Technology, Chinese Research Academy of Environmental Science, Beijing 100012, PR China. E-mail: licongyu1996@163.com; zhongzhen4503@163.com; wanghy@craes.org.cn; yangk@craes.org.cn; docreat@163.com; wanghuan989193@126.com; cy1100@126.com
bNational Engineering Laboratory for Lake Pollution Control and Ecological Restoration, Chinese Research Academy of Environmental Sciences, Beijing 100012, PR China. E-mail: chuzs@craes.org.cn
cChangZhou JiangRun Environmental Protection Technology Co., Ltd, Jiangsu 213200, PR China
dShangcai County Environmental Monitoring Station, Henan 463800, China. E-mail: 767487005@qq.com
First published on 5th August 2021
Abstract
Nitrogen in surface sediments is becoming an ecological risk to the river environment and it is essential to clarify the relationship between the different forms of nitrogen and related microorganisms. A survey was conducted to analyze the distribution characteristics of the nitrogen and related microbial community in the sediments of the Songhua River during normal season and dry season. In the surface sediments of the Songhua River, no total nitrogen (TN) pollution risk was observed according to the U.S. EPA assessment criteria (1000 mg kg−1) for sediment contamination, but TN in several sampling sites (554.9–759.7 mg kg−1) exceeded the alert values (550 mg kg−1) should be concerned according to the guidelines issued by the Ministry of Environment and Energy of Ontario, Canada. The average TN, NH4+–N, NO3−–N and total organic nitrogen (TON) in the surface sediments of the Songhua River during normal season were higher than those in the dry period. TON was the main form of nitrogen in the sediment of Songhua River, NO2−–N content was lowest and no obvious difference was observed between normal and dry seasons. The highest average NH4+–N of both seasons occurred in the Nenjiang River, and the highest average NO3−–N of both seasons were found in the main stream of the Songhua River. The community abundance of AOB genes (1.1 × 107 to 2.5 × 108 copies per g soil in normal season, 7.2 × 105 to 3.3 × 108 copies per g soil in dry season) was higher than that (1.2 × 106 to 9.7 × 107 copies per g soil in normal season, 6.6 × 104 to 3.2 × 107 copies per g soil in dry season) of AOA genes in both normal and dry seasons. The denitrifying nirS genes were predominant in both seasons, and their abundance (1.8 × 106 to 8.0 × 108 copies per g soil) in dry season was higher than that (9.7 × 105 to 4.6 × 108 copies per g soil) in normal season. Moreover, the moisture concentration, pH, dissolved oxygen and different formation of nitrogen were key factors affecting the variation of nitrogen-transformation microorganisms during normal and dry seasons. This research could help to explain the relationship between nitrogen transformation and the related microbial community in the surface sediment, which could provide a scientific basis for water ecological restoration and water environment improvement of Songhua River.
1. Introduction
Nitrogen is a core substance maintaining the ecological balance of water bodies. However, with the rapid social and economic development and the improvement in people's living standards, large amounts of domestic, industrial, and agricultural wastewater with nitrogenous substances are discharged into lakes and rivers, which leads to serious eutrophication and causes potential harm to human and animal health and leads to further ecological deterioration.1–3 Nitrogen pollution of water bodies has become one of the most urgent water environmental issues worldwide.4However, most of the nitrogen in a water body will be mineralized into ammonia nitrogen by sediment microorganisms and then adsorbed on the sediments.5 Sediment is the main nitrogen reservoir for deposited gaseous nitrogen and organic nitrogen, which is essential for natural water environment management.6,7 As the sink and source of nitrogen, the sediments will release nutrients to the overlying water with the changes of ambient conditions as temperature, dissolved oxygen (DO), pH, heavy metal8–10 and water disturbance, and then cause secondary pollution.11 The nitrogen forms in sediments are generally divided into total organic nitrogen (TON) and total inorganic nitrogen (TIN), while TON is predominant.12 Nitrogen distribution in the sediments varies greatly due to the different degrees of mineralization for TON,13 whose morphology mainly consists of animal and plant residues or humus, and the nitrogen cycle does not include all of the nitrogen forms in the sediments.14
Fixed ammonium, i.e., non-exchangeable ammonium, which refers to the NH4+–N in the 2![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1 aluminosilicate crystal layer of minerals and accounts for more than 80% of TIN and 10% of the total nitrogen (TN), cannot be exchanged with neutral salts.15 According to Huo,16 the fixed ammonium in lake sediments ranged from 99.45 to 329.02 mg kg−1, and it had negative correlation with nitrate nitrogen. Fixed ammonium and exchangeable nitrogen were mainly resulted from TON mineralization. In recent years, studies on sediment TON in lakes and reservoirs,16 oceans,17 estuarine wetlands17 and rivers18 are mainly focused on the exchangeable nitrogen, and that about river sediment is limited.18
:1 aluminosilicate crystal layer of minerals and accounts for more than 80% of TIN and 10% of the total nitrogen (TN), cannot be exchanged with neutral salts.15 According to Huo,16 the fixed ammonium in lake sediments ranged from 99.45 to 329.02 mg kg−1, and it had negative correlation with nitrate nitrogen. Fixed ammonium and exchangeable nitrogen were mainly resulted from TON mineralization. In recent years, studies on sediment TON in lakes and reservoirs,16 oceans,17 estuarine wetlands17 and rivers18 are mainly focused on the exchangeable nitrogen, and that about river sediment is limited.18
Microorganisms in the sediment can convert nitrogen into different forms by nitrification, denitrification and anammox process.19,20 The nitration process mainly consists of two steps, i.e., oxidation of ammonia and nitrite oxidation, and it is the first and foremost stage of nitrogen cycle. The microorganisms of ammonia oxidation archaea (AOA), ammonia oxidation bacteria (AOB) and nitrosation bacteria (NOB) work together to ensure the smooth progress of nitrification.21,22 AOB and AOA are considered as the most important bacteria and archaea for ammonia oxidation.23–25
AOA and AOB abundance varies in different environments, and AOB communities are dominant in river sediments,26 coastal wetlands27 and cultivated farmland.28 The number of amoA genes of AOA is higher than that of AOB in the hydrothermal spring sediments29 and marine environment.30 It is reported that low ammonia nitrogen concentration, low pH and anaerobic condition is suitable for AOA growth.31 When the circumstances are suitable for the growth of both AOA and AOB, the growth rate of AOB is much higher than that of AOA.32 In the nitrite oxidation stage, NO2− produced by ammonification is oxidized to NO3− by NOB, which interacts with AOB to promote the nitrification process. NOB concentration in different ecosystems changes obviously.33,34 In river sediments, nitrification bacteria are always dominated by Nitrospira group, whose abundance is greater than that of AOB.35 The role and concentration of AOA, AOB and NOB in river sediments, as well as their cooperative effects and impacting factors need to be further studied.
Denitrification is recognized as the best way to remove nitrogen from rivers,36 and the sediment and water interface is very suitable for the growth of denitrification microorganisms. Denitrification process consists of four consecutive steps, i.e., 2NO3− → 2NO2− → 2NO → N2O → N2, and nitrate reductase (encoding genes are mainly narG and napA), nitrite reductase (nirS and nirK), NO reductase (norB), and N2O reductase (nosZ) are essential to catalyze these steps. NarG, nirS, nirK and nosZ genes are the most important functional genes in the denitrification microorganisms,37 and nirS has higher genetic diversity than nirK in nitrite reductase related genes.38 It is reported that nirS related bacteria played more important role than nirK related bacteria in the denitrification process of San Francisco Bay estuary sediments, and the denitrification potential rate is positively correlated with nirS gene abundance.39 The investigation of denitrification rate and dissimilation efficiency of microorganisms is a hot issue.40
Previous studies on nitrogen in sediments are mostly focused on the sediments of sea, estuary and coastal zones, while the distribution characteristics of nitrogen in river sediments were seldom reported. Also, few analyses about the nitrogen morphology and denitrification related microorganisms were conducted. In this study, we seasonally collected surface sediment samples from main stream and tributaries along Songhua River. The characteristics of nitrogen distribution, related microbial community and their correlation in the surface sediments of the Songhua River during normal and dry seasons were extensively investigated in this paper, and the sediment TN pollution risk was also assessed, thus provided scientific basis for the nitrogen source analysis, water quality improvement and water ecological restoration.
2. Materials and methods
2.1 Studying region and site sampling
Located in the northeast of China, the Songhua River basin covers 556800 km2, which accounts for about 60% of the whole northeast China region. The river basin is surrounded by mountains on three sides, i.e., the Changbai Mountains in the east and southeast, the Greater Khingan Mountains in the west and northwest and the Lesser Khingan Mountains in the north, and the mountainous area accounts for more than 60%. The Songhua River has two headstreams, i.e., the Nenjiang River (NR) originated from the Greater Khingan Mountains and the Second Songhua River (SSR) originated from the Changbai Mountains, which converge at the Sancha River mouth and finally join the Heilong River (HR).41 Heilong River marks the border between China and Russia. In Songhua River basin, rainfall mainly occurs in July and August, and the average annual precipitation ranges from 400 mm to 700 mm. September and March were selected as the representative month of the normal season and the dry season respectively.42 In the normal water season of September, 2016, 43 surface sediment samples were obtained from the Songhua River basin, i.e., 8 from NR (NS1–NS8), 8 from SSR (DS1–DS6), 27 from main stream of Songhua River (MSSR) (SS1–SS27), and 2 from the HR (HS1–HS2), which is the reference sampling sites Fig. 1(a). 19 samples were taken in the dry season of March, 2017, i.e., NM2, NM4, NM6, NM8 from NR, DM1, DM2, and DM6 from SSR, SM1, SM2, SM6, SM7, SM8, SM9, SM10, SM18, SM20, and SM21 from MSSR, and HM1 and HM2 from HR (Fig. 1(b)), which were coincided with the corresponding sites in September, 2016. The sampling sites information is shown in the ESI Table S1,† and the same sample sites of normal and dry season (Fig. 1(c)) were selected for comparative analysis. The sites basically covered the entire river basin and the important cities along the route, and some of the sites located at the positions of structures such as bridges on the river which can change the flow conditions.
 | ||
| Fig. 1 Distribution diagram of sampling sites in the Songhua River (a) September, (b) March, (c) comparative analysis sites. | ||
2.2. Sample collection and pretreatment
After the removal of the gravel, plant roots, etc., the surface sediments (from the 20 cm upper part) were packed into polyethylene self-sealing bags and transported to the laboratory with ice bags as soon as possible.43 The samples were frozen at −20 °C, dried by freeze-dryer, grounded by mortar, and then passed through 100-mesh nylon sieve for storage. The samples for molecular biological analysis were stored at −80 °C for DNA extraction.2.3 Analytical method
| Gene | Primers | Primer sequence | References |
|---|---|---|---|
| AOA amoA | Arch-amoA26F | GACTACATMTTCTAYACWGAYTGGGC | 45 |
| Arch-amoA417R | GGKGTCATRTATGGWGGYAAYGTTGG | ||
| AOB amoA | amoA-1F | GGGGTTTCTACTGGTGGT | 45 |
| amoA-2R | CCCCTCKGSAAAGCCTTCTTC | ||
| narG (f/r) | narG-F | TCGCCSATYCCGGCSATGTC | 46 |
| narG-R | GAGTTGTACCAGTCRGCSGAYTCSG | ||
| nirK | nirK-583F | TCATGGTGCTGCCGCGKGACGG | 47 |
| nirK-909R | GAACTTGCCGGTPGCCCAGAC | ||
| nirS | nirS-cd3af | GTSAACGTSAAGGARACSGG | 47 |
| nirS-R3cd | GASTTCGGRTGSGTCTTGA | ||
| nosZ | nosZ-1527F | CGCTGTTCHTCGACAGYCA | 47 |
| nosZ-1773R | ATRTCGATCARCTGBTCGTT |
qPCR reaction conditions were as follows: pre-denaturation at 95 °C for 5 min, denaturation at 95 °C for 10 s, total 40 cycles, and finally extension at 60 °C for 34 s. PCR reaction mixtures contained 16 μL SYBR GreenI (GENEary, GK8020), 1 μL positive primers (10 pmol μL per a), 1 μL reverse primers (10 pmol μL per a) and 2 μL DNA template. The 10−2 to 10−6 diluents of AOA and AOB standard, narG, nirS and nosZ standard, and 10−3 to 10−7 diluents were selected to prepare the standard curve through pre-experiment. All the standard curves showed excellent correlations between the DNA template concentration and the crossing point with high coefficients of determination (R2 > 0.99).
2.4 The relationship between microorganisms and environmental factors
The relationship between microorganisms and environmental factors were analysed using Canoco 4.5 analysis software, and redundancy analysis (RDA) mainly included environmental and microbial data matrix. The microbial data matrix de-trend correspondence analysis (DCA) was performed, and then the sorting method was determined according to the gradient in the axis. In this study, the calculated length of axial gradient was less than 3, so RDA was chosen to analyze the relationship between microorganisms and environmental factors,48 and SPSS 22 was used for correlation analysis. The graphics were drawn using ArcGIS (Version 10.2, ESRI).3. Results and discussion
3.1 Temporal and spatial distribution of nitrogen in the Songhua River sediments
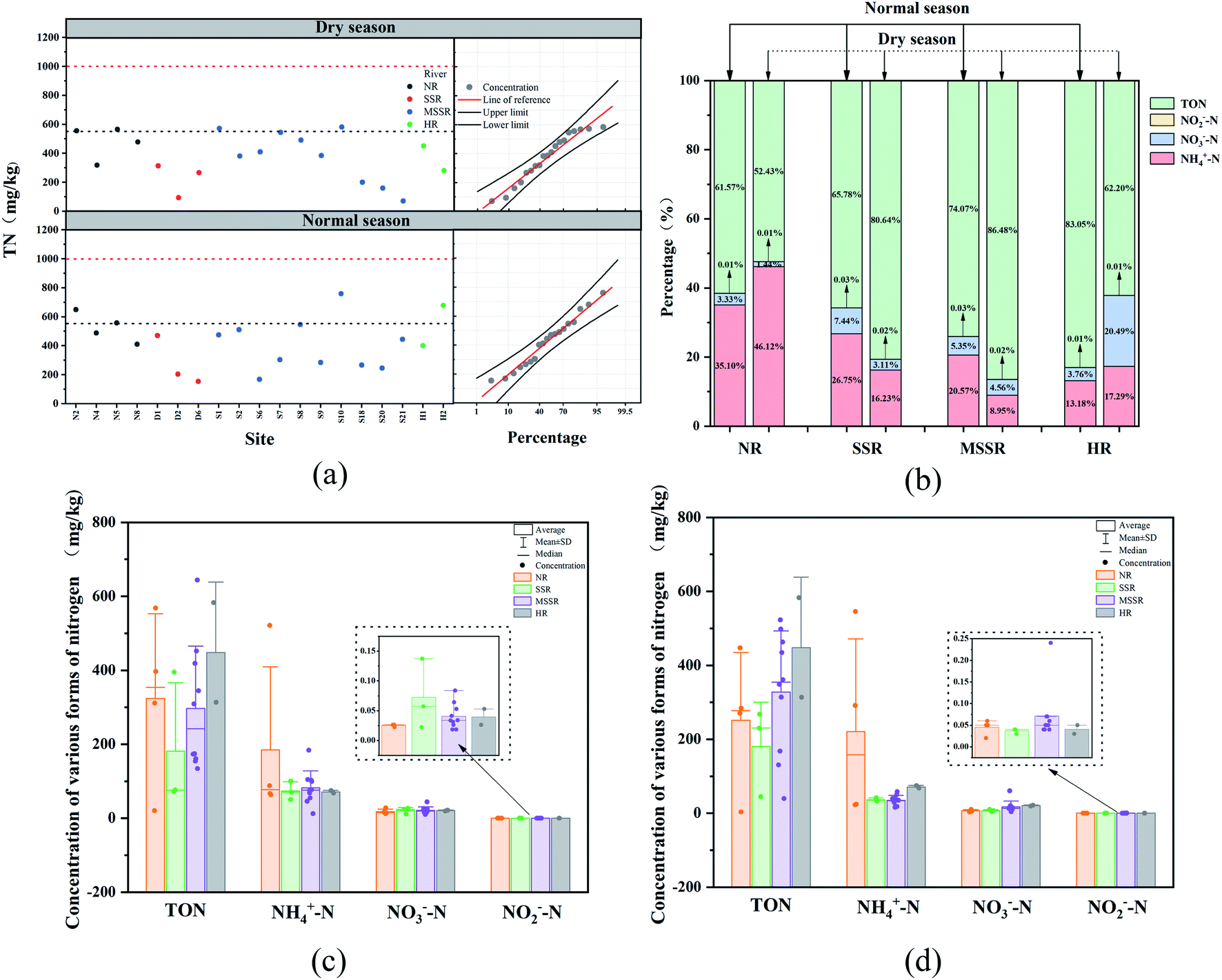 | ||
| Fig. 2 Nitrogen forms and concentration in surface sediments of the Songhua River during different water seasons (a) TN, (b) the ratios of different nitrogen forms in the 4 basins, (c) 4 forms of nitrogen in normal season, (d) 4 forms of nitrogen in dry season. | ||
TN variation in different sampling sites may be related to the different sources of nitrogen pollution and human activities along the Songhua River. TN concentration of the 8 sites in NR changed slightly with quite high value, which may be attributed to the numerous farmlands and its non-point source discharge along the river, moreover, industrial wastewater containing nitrogen might be discharged from Qiqihar and other industrial cities in NR basin. Besides the upstream inflow, the nitrogen pollution of MSSR was from Harbin, Kiamusze and other industrial cities, and the plenty of farmland non-point source in the river basin. Because of the water conservancy projects, the velocity of the main stream slows down, which leads to the decrease of dissolved oxygen (DO) and then the weakening of nitrogen purification capacity. Therefore, the sediment nitrogen in MSSR is rather high. Quite a lot petrochemical enterprises and heavy industries were unevenly distributed in SSR basin, so the sediment TN was quite different. Moreover, TN in SSR was lower than that in NR and MSSR, which indicated that non-point source pollution in SSR is not the main influencing factor.
Table 2 shows the TN concentration of surface sediments at different sampling sites in both dry and normal seasons in the Songhua River basin, from which the TN in normal season is higher than that in dry season. Compared with the sediment TN of lakes and rivers at home and abroad as Taihu Lake,53 Xuanwu Lake,54 Erhai Lake,11 Chaohu Lake,55 Han River56 and Tigris River,57 TN in the surface sediments of the Songhua River in both normal and dry season was lower. As a whole, surface sediments of the Songhua River were TN-pollution-free, but some sections need to be concerned.
| River basin | Water period | TN (mg kg−1) |
|---|---|---|
| Nenjiang River | Normal season | 410.6–649.0 |
| Dry season | 318.2–565.3 | |
| Second Songhua River | Normal season | 153.3–469.2 |
| Dry season | 93.0–312.8 | |
| Main stream of Songhua River | Normal season | 168.5–759.7 |
| Dry season | 70.5–582.0 | |
| Heilong River | Normal season | 400.3–678.8 |
| Dry season | 281.2–451.1 |
(1) NH4+–N. NH4+–N in the surface sediments of the Songhua River is shown in Fig. 3(a), which ranged from 12.2 mg kg−1 to 521.3 mg kg−1 with the average of 101.3 mg kg−1 in normal season. NH4+–N in the surface sediments of NR, SSR, MSSR, and HR were 184.7 mg kg−1, 73.6 mg kg−1, 82.4 mg kg−1, and 71.1 mg kg−1 respectively. During dry season, NH4+–N were changed between 15.8 mg kg−1 and 545.6 mg kg−1 with the average value of 76.8 mg kg−1, and NH4+–N in the surface sediments of NR, SSR, MSSR, and HR were 221.0 mg kg−1, 36.4 mg kg−1, 33.9 mg kg−1, and 63.3 mg kg−1 respectively. In general, sediment NH4+–N in normal season was higher than that in dry season, and the highest value appeared in Nenjiang River, which was resulted from the overlap of cultivation, surface runoff and increase utilization of nitrogenous fertilizer.58 Also, a higher concentration of NH4+–N in the surface sediment suggested that ammonification was stronger in the oxic surface sediment, which carried out by most heterotrophic bacteria.59
 | ||
| Fig. 3 Different forms of nitrogen in surface sediments of the Songhua River basin (a) NH4+–N, (b) NO3−–N, (c) NO2−–N, (d)TON. | ||
(2) NO3−–N. As shown in Fig. 3(b), the sediment NO3−–N of the Songhua River were between 10.1 mg kg−1 and 44.1 mg kg−1 during normal season with an average of 20.3 mg kg−1, and the average NO3−–N in the surface sediments of NR, SSR, MSSR, and HR were 17.5 mg kg−1, 20.5 mg kg−1, 21.4 mg kg−1, and 20.3 mg kg−1 respectively. In dry season, sediment NO3−–N ranged from 3.2 mg kg−1 to 146.5 mg kg−1 with an average of 19.6 mg kg−1, and NO3−–N in the surface sediments of NR, SSR, MSSR, and HR were 6.9 mg kg−1, 17.3 mg kg−1, 7.0 mg kg−1, and 75.0 mg kg−1. The concentration of NO3−–N in normal season was higher than that in dry season.
(3) NO2−–N. The mean value of sediment NO2−–N was 0.04 mg kg−1 (ranged from 0.02 to 0.14 mg kg−1) in normal season, and the average NO2−–N in the surface sediments of NR, SSR, MSSR, and HR were 0.03 mg kg−1, 0.07 mg kg−1, 0.04 mg kg−1, and 0.04 mg kg−1. In dry season, the sediment NO2−–N ranged from 0.02 mg kg−1 to 0.24 mg kg−1 with a mean value of 0.06 mg kg−1, and NO2−–N in the surface sediments of NR, SSR, MSSR, and HR were 0.04 mg kg−1, 0.04 mg kg−1, 0.07 mg kg−1, and 0.04 mg kg−1 respectively. Generally, the nitrite concentration in the surface sediments of the Songhua River is low, and its difference between the two water periods was not significant. Surface sediments were characterized by lower concentrations of nitrite and nitrate, which meant that bacterial assimilation and dissimilation (e.g., denitrification) contributes significantly to nitrogen removal,60 the nitrite or nitrate that were transformed to N2 by coupled nitrification and denitrification.61
(4) TON. Sediment TON was relatively high in normal season with an average of 300.0 mg kg−1 (ranged from 20.3 mg kg−1 to 644.2 mg kg−1), and the mean value in the surface sediments of NR, SSR, MSSR and HR were 324.0 mg kg−1, 181.0 mg kg−1, 296.5 mg kg−1, and 448.1 mg kg−1 respectively. Average TON was 278.1 mg kg−1 (ranged from 3.4 mg kg−1 to 522.7 mg kg−1) in dry season, which were 251.2 mg kg−1, 180.7 mg kg−1, 328.1 mg kg−1, and 227.7 mg kg−1, respectively. It can be seen from Fig. 4(d) that the sediment TON in HR was relatively high in normal season, but no significant TON difference was observed between normal and dry season for NR, SSR, and MSSR. Moreover, the range of TON concentration was quite wide in the whole Songhua River basin. TON was the main form of nitrogen in the sediment of Songhua River, which was consistent with Lu.12
 | ||
| Fig. 4 (I) Relative abundance and concentration of selected functional genes in the sediments (copies per g soil) and (II) (a) AOA (b) AOB genes copies in nitrification process and (c) narG, (d) nirK, (e) nirS, (f) nosZ in denitrification process during dry and normal seasons. | ||
3.2 Distribution characteristics of microbes related to nitrogen transformation in sediments
The relative abundance of the functional genes involved in main nitrogen cycles at all sampling sites are shown in Fig. 4. The abundance and distribution of nitrogen transformation related functional genes varied remarkably between normal and dry seasons, based on which the 19 sampling sites were classified into 3 distinct clusters by hierarchical cluster analysis and then illustrated as a dendrogram (Fig. 4), and then the same dominant microbe in different sites can distinguished.Fig. 4(a) shows the relevant microbial biomass during the nitrification process in both normal and dry seasons of the Songhua River, from which it can be seen that the sediment number of AOA genes (1.2 × 106 to 9.7 × 107 copies per g soil) in normal season was significantly higher than that (6.6 × 104 to 3.2 × 107 copies per g soil) in dry season. There was not much difference between AOB genes in dry season (ranged from 7.2 × 105 to 3.3 × 108 copies per g of soil) and those in normal season, i.e., from 1.1 × 107 to 2.5 × 108 copies per g of soil. Through the comparison of the quantities of sediment AOA and AOB genes, it can be concluded that AOB were the main ammonia-oxidation bacteria during the nitrification process in the sediments of the Songhua River. Considering the frozen water surface and low DO in dry season and the strong affinity of AOB with oxygen,62 the AOB genes had relatively high abundance in this research.
NH4+–N is a major factor affecting ammonia oxidation process, and AOA grow better than AOB in low NH4+–N environment.11 According to the analysis in Section 3.1, among all the nitrogen forms, TON had the highest concentration, followed by NH4+–N in the surface sediments of the Songhua River basin. Therefore, the relatively high sediment NH4+–N was conducive to the growth of AOB.
The number of microorganisms related to denitrification in normal and dry sesaons of the Songhua River basin were illustrated in Fig. 4. The number of narG genes in the sediments during normal season (4.3 × 105 to 5.8 × 107 copies per g soil) was greater than that in dry season (3.2 × 105 to 2.5 × 106 copies per g soil). The amount of nirK and nirS genes (4.3 × 104 to 2.9 × 107 copies per g soil and 9.7 × 105 to 4.6 × 108 copies per g soil) in normal season was much less than that during dry season (2.7 × 106 to 7.3 × 107 copies per g soil and 1.8 × 106 to 8.0 × 108 copies per g soil). The number of nosZ genes in dry season (1.8 × 105 to 5.9 × 106 copies per g soil) had slight difference with that in normal season (6.4 × 105 to 4.0 × 106 copies per g soil).
The microorganisms related with nirS genes were dominant in both normal and dry water periods, and the microorganisms related with narG genes were 1 to 2 orders of magnitude lower than those related with other genes, which was corresponding to the low sediment nitrite in the Songhua River according to the analysis in 3.1.2 section. The number of narG genes in normal season was significantly higher than that in dry season, which might be due to the higher DO caused by water flow and natural aeration during normal season, previous study also found that oxygen is often considered as a significant factor controlling the nitrogen loss processes in sediment.61 Since nitrification is oxygen consumption process, it provides sufficient NO3−–N for the denitrification process in case of sufficient DO. In dry season, the sediment surface was covered by 1 meter-thick ice, which leads to the decrease of DO, and then results in the conversion of NO3−–N to N2O or N2 by denitrifying bacteria.64 The number of nirS genes in N5, D2 and S1 was one order of magnitude higher than that in other sampling sites during normal season.
3.3 Relationship between nitrogen transformation microorganisms and physical, chemical factors
 | ||
| Fig. 5 The RDA analysis ordination chart of the microbial environmental factors and Spearman correlation analysis during normal season ((a) RDA analysis, (b) Spearman analysis) and dry season ((c) RDA analysis, (d) Spearman analysis). ** is shown as significant correlation at the level of 0.01 (bilateral). * is shown as significant correlation at the level of 0.05 (bilateral). | ||
It can be seen from Fig. 5(a) that sediment MC and water DO were positively correlated with nitrite reductase coding genes nirS and nirK, and DO and nirK gene abundance were remarkably correlated (p < 0.05) in normal season. Sediment MC and water DO are the key influencing factors on the redox potential, and high redox potential facilitates nitrification while low redox reduction potential promotes denitrification. The alternation of nitrification and denitrification achieves the purification of nitrogenous sediment pollution. Denitrification rate increases sharply when the soil moisture exceeds 70–75% of the soil water-filled pores, and slight variation in water concentration will change the denitrification rate,66 which is consistent with the results of this study. As for DO, the water presented aerobic state at high DO levels (DO > 0.5 mg L−1), and nitrifying bacteria could carry out nitrification and convert most ammonia nitrogen in the water to nitrate nitrogen, which increased the concentration of nitrate nitrogen in the water, and the abundance of denitrifying genes was relatively high. The pH and narG gene abundance showed obvious negative correlation (p < 0.05), which might be due to the decrease of sediment nitrogen mineralization caused by pH reduction.67,68 pH affects the microbial activities, and the optimal pH for denitrification is 7.0–8.0.69 The pH of the sediments in the Songhua River was between 6 and 8 in normal season, which was consistent with the reported researches. The ammonification and nitrification of nitrogen are generally called nitrogen mineralization, which requires the participation of a variety of microorganisms. The produced ammonia will be consumed by microorganisms and plants, fixed by clay minerals, or converted to nitrate by autotrophic or heterotrophic nitrification.70,71
No significant correlation could be observed between the nitrogen conversion related microorganisms and water content, TP, pH and DO (Fig. 5(c)). Previous studies reported that DO was positively correlated with AOB amoA gene abundance and negatively correlated with AOA amoA gene abundance, which might be resulted from the far less water DO in dry season than that in normal season.72 AOA and AOB had strong affinity and high abundance in suitable low oxygen environment, while AOA could tolerate even lower DO than AOB. The related narG, nirS, nirK, and nosZ genes were also affected by the low DO concentration in dry season. As shown in Fig. 5(c), pH was negatively correlated with AOA and positively correlated with AOB in the nitrification process. The sediment pH of the Songhua River in dry season ranged from 6.4 to 7.7, which was relatively moderate in the environment, and the nitrification rate increased with the increase of pH within this range.73
4. Conclusion
Comprehensive study about the distribution of different nitrogen forms and the characteristics of relative microbial community was carried out about the surface sediments of the Songhua River in dry and normal seasons. TN (70.8–678.8 mg kg−1) in several sampling sites (678.8–759.7 mg kg−1 in normal season and 554.9–678.8582 mg kg−1 in dry season) exceeded the alert values 550 mg kg−1, TN, NH4+–N, NO3−–N, and TON in normal season were higher than those in dry season. TON was identified as the main form of nitrogen in the sediments of the Songhua River basin during both dry and normal seasons, and the highest concentration of which was 328.1 mg kg−1 in MSSR in dry period, and the second largest value was 324.0 mg kg−1 in NR during normal season. As for bacteria communities and functional genes structures in the sediments, AOB was dominant for the nitrification process in both dry and normal seasons. AOB genes abundance (7.2 × 105 to 3.3 × 108 copies per g soil) in dry season was higher than that (1.1 × 107 to 2.5 × 108 copies per g soilin normal season), while AOA amoA genes abundance had the reverse distribution (1.2 × 106 to 9.7 × 107 copies per g soil in normal season and 6.6 × 104 to 3.2 × 107 copies per g soil in dry season). The denitrification nirS genes were predominant in both dry and normal seasons, the abundance (1.8 × 106 to 8.0 × 108 copies per g soil) of which in dry season was higher than that (9.7 × 105 to 4.6 × 108 copies per g soil) in normal season. Physical and chemical factors such as pH, DO, MC, TP and different forms of nitrogen in the sediments had great influence on the profiles of functional genes, bacterial communities, and co-occurrence patterns between the functional genes and bacterial taxa. This study is beneficial to explain the influence of nitrogen forms on the nitrogen transformation microbial community of the sediment in Songhua River, which could afford the technical support for the water ecological restoration of the Songhua River Basin.Author contributions
Congyu Li: conceptualization, methodology, investigation, validation, writing – original draft. Zhen Zhong: conceptualization, methodology, validation, supervision, resources. Wenfu Wang: supervision. Haiyan Wang: investigation, validation, writing – review & editing. Weiyang Dong: resources. Guokai Yan: validation, conceptualization, resources. Zhaosheng Chu: supervision, resources. Huan Wang: supervision validation. Yang Chang: supervision, validation.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This study was supported by National Major Science and Technology Program for Water Pollution Control and Treatment (2015ZX07201-008, 2017ZX07401003-05). The authors also appreciate support from Research Center for Environmental Pollution Control Engineering at Chinese Research Academy of Environmental Sciences.References
- E. B. McAleer, C. E. Coxon, K. G. Richards, M. M. R. Jahangir, J. Grant and P. E. Mellander, Sci. Total Environ., 2017, 586, 372–389 CrossRef CAS.
- W. W. Wong, C. Greening, G. Shelley, R. Lappan, P. M. Leung, A. Kessler, B. Winfrey, S. C. Poh and P. Cook, Sci. Total Environ., 2021, 790, 147749 CrossRef CAS.
- D. Deng, Y. Pan, G. Liu, W. Liu and L. Ma, Sci. Total Environ., 2020, 737, 140253 CrossRef CAS PubMed.
- J. N. Galloway, A. R. Townsend, J. W. Erisman, M. Bekunda, Z. Cai, J. R. Freney, L. A. Martinelli, S. P. Seitzinger and M. A. Sutton, Science, 2008, 320, 889 CrossRef CAS.
- Y. C. Ren, S. L. Dong, F. Wang, Q. F. Gao, X. L. Tian and F. Liu, Aquacult. Res., 2010, 42, 14–21 CrossRef.
- S. H. Cui, Y. L. Shi, P. M. Groffman, W. H. Schlesinger and Y. G. Zhu, Proc. Natl. Acad. Sci. U. S. A., 2013, 110, 2052 CrossRef CAS.
- D. M. Orihel, H. M. Baulch, N. J. Casson, R. L. North, C. T. Parsons, D. C. M. Seckar and J. J. Venkiteswaran, Can. J. Fish. Aquat. Sci., 2017, 1–25 Search PubMed.
- V. Turan, Ecotoxicol. Environ. Saf., 2019, 183, 109594 CrossRef CAS PubMed.
- V. Turan, Chemosphere, 2020, 245, 125611 CrossRef CAS PubMed.
- V. Turan, Physiol. Plant., 2021, 1–12 Search PubMed.
- Z. Ni and S. Wang, Ecol. Eng., 2015, 79, 42–53 CrossRef.
- R. K. Lu, China Agric. Sci. Technol., 2000, 106–310 Search PubMed.
- A. L. Herczeg, A. K. Smith and J. C. Dighton, Appl. Geochem., 2001, 16, 73–84 CrossRef CAS.
- W. S. Gardner, L. Yang, J. B. Cotner, T. H. Johengen and P. J. Lavrentyev, J. Great Lakes Res., 2001, 27, 84–97 CrossRef CAS.
- D. Cavalli, G. Consolati, P. Marino and L. Bechini, Geoderma, 2015, 259–260, 116–125 CrossRef CAS.
- S. L. Huo, J. T. Zhang, B. D. Xi, F. Y. Zan, J. Su and H. Yu, Environ. Earth Sci., 2014, 71, 2167–2175 CrossRef CAS.
- S. M. Liu, B. D. Zhu, J. Zhang, Y. Wu, G. S. Liu, B. Deng, M. X. Zhao, G. Q. Liu, J. Z. Du, J. L. Ren and G. L. Zhang, Mar. Pollut. Bull., 2010, 60, 1591–1599 CrossRef CAS PubMed.
- Q. J. Guo, C. Y. Wang, R. F. Wei, G. X. Zhu, M. Cui and C. P. Okolic, Ecotoxicol. Environ. Saf., 2020, 195, 110436 CrossRef CAS.
- R. J. Bernard, B. Mortazavi and A. A. Kleinhuizen, Biogeochemistry, 2015, 125, 47–64 CrossRef CAS.
- A. K. Hardison, C. K. Algar, A. E. Giblin and J. J. Rich, Geochim. Cosmochim. Acta, 2015, 164, 146–160 CrossRef CAS.
- G. Kowalchuk and J. Stephen, Annu. Rev. Microbiol., 2001, 55, 485–529 CrossRef CAS PubMed.
- C. Okkyoung and H. Zhiqiang, Environ. Sci. Technol., 2008, 42(12), 4583–4588 CrossRef.
- C. A. Francis, K. J. Roberts, J. M. Beman, A. E. Santoro and B. B. Oakley, Proc. Natl. Acad. Sci. U. S. A., 2005, 102(41), 14683 CrossRef CAS PubMed.
- J. C. Venter, K. Remington, J. F. Heidelberg, A. L. Halpern, D. Rusch, J. A. Eisen, D. Wu, I. Paulsen, K. E. Nelson, W. Nelson, D. E. Fouts, S. Levy, A. H. Knap, M. W. Lomas, K. Nealson, O. White, J. Peterson, J. Hoffman, R. Parsons, H. Baden-Tillson, C. Pfannkoch, Y. H. Rogers and H. O. Smith, Science, 2004, 304(5667), 66 CrossRef CAS.
- M. Könneke, A. E. Bernhard, J. R. de la Torre, C. B. Walker, J. B. Waterbury and D. A. Stahl, Nature, 2005, 437(7058), 543–546 CrossRef.
- Y. Wang, G. B. Zhu, L. Ye, X. J. Feng, H. J. M. Op den Camp and C. Q. Yin, J. Environ. Sci., 2012, 24, 790–799 CrossRef CAS.
- X. Y. Wang, C. Wang, L. L. Bao and S. G. Xie, Ann. Microbiol., 2015, 65, 1411–1418 CrossRef CAS.
- X. Y. Wang, C. Wang, L. L. Bao and S. G. Xie, Appl. Microbiol. Biotechnol., 2014, 98, 1883–1892 CrossRef CAS.
- C. L. Zhang, Q. Ye, Z. Huang, W. Li, J. Chen, Z. Song, W. Zhao, C. Bagwell, W. P. Inskeep, C. Ross, L. Gao, J. Wiegel, C. S. Romanek, E. L. Shock and B. P. Hedlund, Appl. Environ. Microbiol., 2008, 74, 6417 CrossRef CAS PubMed.
- S.-J. Park, B.-J. Park and S.-K. Rhee, Extremophiles, 2008, 12, 605 CrossRef CAS PubMed.
- T. H. Erguder, N. Boon, L. Wittebolle, M. Marzorati and W. Verstraete, FEMS Microbiol. Rev., 2009, 33, 855–869 CrossRef CAS.
- M. Herrmann, A. Scheibe, S. Avrahami and K. Küsel, Appl. Environ. Microbiol., 2011, 77, 1896–1899 CrossRef CAS.
- R. D. Chen and T. M. LaPara, Process Biochem., 2008, 43, 33–41 CrossRef CAS.
- J. Ji, Y. Peng, X. Li, Q. Zhang and X. Liu, Water Res., 2020, 175, 115690 CrossRef CAS PubMed.
- Y. Nakamura, H. Satoh, T. Kindaichi and S. Okabe, Environ. Sci. Technol., 2006, 40, 1532–1539 CrossRef CAS.
- E. W. Boyer, R. W. Howarth, J. N. Galloway, F. J. Dentener, P. A. Green and C. J. Vörösmarty, Global Biogeochem. Cycles, 2006, 20, GB1S91 CrossRef.
- L. Philippot and S. Hallin, Curr. Opin. Microbiol., 2005, 8, 234–239 CrossRef CAS.
- A. Priemé, G. Braker and J. Tiedje, Appl. Environ. Microbiol., 2002, 68, 1893–1900 CrossRef.
- A. Mosier and C. Francis, Environ. Microbiol. Rep., 2010, 2(5), 667–676 CrossRef CAS PubMed.
- I. J. Washbourne, C. Crenshaw and M. Baker, Aquat. Microb. Ecol., 2011, 65, 55–64 CrossRef.
- Y. Yang, H. Wang, Y. Chang, G. Yan, Z. Chu, Z. Zhao, L. Li, Z. Li and T. Wu, Mar. Pollut. Bull., 2020, 152, 110923 CrossRef CAS PubMed.
- T. Li, Z. Zhou, Q. Fu, D. Liu, M. Li, R. Hou, W. Pei and L. Li, J. Water Clim. Change, 2019, 11, 839–864 CrossRef.
- V. Ruban, J. F. López-Sánchez, P. Pardo, G. Rauret, H. Muntau and P. Quevauviller, J. Environ. Monit., 2001, 3, 121–125 RSC.
- X. Jiang, Handbook of Sediment Quality Survey and Assessment, Science Press, 2012, (in Chinese) Search PubMed.
- G. D. Ji, W. Zhi and Y. F. Tan, Ecol. Eng., 2012, 44, 269–277 CrossRef.
- A. Vilar-Sanz, S. Puig, A. García-Lledó, R. Trias, M. Balaguer, J. Colprim and L. Bañeras, PLoS One, 2013, 8, e63460 CrossRef CAS PubMed.
- I. Throbäck, K. Enwall, A. Jarvis and S. Hallin, FEMS Microbiol. Ecol., 2004, 49, 401–417 CrossRef.
- E. V. Etten, Austral Ecol., 2005, 30, 486–487 CrossRef.
- W. Jiang, Y. Xing, X. Wang, X. Liu and Z. Cui, Sustainability, 2020, 12, 1–12 Search PubMed.
- L. Yuan, X. Chen, J. Jia, H. Chen, Y. Shi, J. Ma, C. Liang, Y. Liu, H. Xie, H. He, X. Zhang, X. Peng and C. Lu, Agric., Ecosyst. Environ., 2021, 312, 107360 CrossRef CAS.
- S. Cieniawski, D. Macdonald and C. Ingersoll, An Ecosystem-Based Framework for Assessing and Managing Contaminated Sediments, A Guidance Manual to Support the Assessment of Contaminated Sediments in Freshwater Ecosystems, 2020, vol. 1 Search PubMed.
- H. V. Leland, W. E. Dean, A. Mudroch and J. M. Azcue, J. North Am. Benthol. Soc., 1995, 16, 296 CrossRef.
- S. Y. Lu, Y. Yuan, X. C. Jin, W. Jiao, Y. J. Wu, D. Y. Ren, Y. H. Zhou and L. Chen, Environ. Sci., 2012, 33, 1497–1502 Search PubMed (in Chinese)..
- J. Zeng, L. Y. Yang, H. W. Du, L. Xiao, L. J. Jiang, J. Wu and X. L. Wang, World J. Microbiol. Biotechnol., 2009, 25, 763–772 CrossRef CAS.
- Y. Y. Fan, B. B. Li, Z. C. Yang, Y. Y. Cheng, D. F. Liu and H. Q. Yu, Environ. Pollut., 2019, 250, 233–240 CrossRef CAS.
- D. H. Li, M. Kim, W. J. Shim, U. H. Yim, J.-R. Oh and Y.-J. Kwon, Chemosphere, 2004, 56, 1–6 CrossRef CAS.
- M. Varol and B. Şen, Catena, 2012, 92, 1–10 CrossRef CAS.
- E. D. Ongley, Z. Xiaolan and Y. Tao, Environ. Pollut., 2010, 158, 1159–1168 CrossRef CAS.
- C. L. Ettinger, S. L. Williams, J. M. Abbott, J. J. Stachowicz and J. A. Eisen, PeerJ, 2017, 5, e3674 CrossRef.
- Y. Li, C.-X. Li, W. Lin, S.-S. Wang, W.-X. Zhang, Y.-M. Jiang, Y. Zhang, H. Zhang and Y.-H. Han, Environ. Technol. Innovation, 2021, 23, 101731 CrossRef CAS.
- J. Wu, Y. Hong, X. Liu and Y. Hu, Environ. Pollut., 2021, 286, 117267 CrossRef CAS.
- Y. W. Yang, X. L. Gao and P. M. Li, Environ. Sci., 2012, 33(10), 3449–3456 CAS.
- A. L. W. Kemp and A. Mudrochova, Limnol. Oceanogr., 1972, 17, 855–867 CrossRef CAS.
- X. Q. Li, C. Q. Lu, Y. F. Zhang, H. D. Zhao, J. Y. Wang, H. B. Liu and K. D. Yin, Mar. Pollut. Bull., 2020, 151, 110814 CrossRef CAS.
- H. W. Chen, H. C. Lin, Y. H. Chuang, C. T. Sun, W. Y. Chen and C. Y. Kao, Ocean Coast. Manage., 2019, 169, 37–49 CrossRef.
- F. P. Vinther, Biol. Fertil. Soils, 1992, 14, 43–48 CrossRef CAS.
- A. M. Blackmer and J. M. Bremner, Soil Biol. Biochem., 1978, 10, 187–191 CrossRef CAS.
- D. J. Mladenoff, Ecology, 1987, 68, 1171–1180 CrossRef.
- M. Šimek, L. Jišova and D. W. Hopkins, Soil Biol. Biochem., 2002, 34, 1227–1234 CrossRef.
- M. Akcura, V. Turan, K. Kokten and M. Kaplan, Ind. Crops Prod., 2019, 128, 140–146 CrossRef CAS.
- V. Turan, P. Schröder, S. Bilen, H. Insam and M. Fernández-Delgado Juárez, Sci. Rep., 2019, 9, 15178 CrossRef.
- S. Tomoko, Microbes Environ., 2012, 1, 61–66 Search PubMed.
- Y. Y. Li, S. J. Chapman, G. W. Nicol and H. Y. Yao, Soil Biol. Biochem., 2018, 116, 290–301 CrossRef CAS.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d1ra04903h |
| This journal is © The Royal Society of Chemistry 2021 |