
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceNanocellulose-based mechanically stable immobilization matrix for enhanced ethylene production: a framework for photosynthetic solid-state cell factories†
V.
Rissanen‡
a,
S.
Vajravel‡
b,
S.
Kosourov
b,
S.
Arola
a,
E.
Kontturi
 c,
Y.
Allahverdiyeva
*b and
T.
Tammelin
*a
c,
Y.
Allahverdiyeva
*b and
T.
Tammelin
*a
aVTT Technical Research Centre of Finland Ltd, VTT, PO Box 1000, FI-02044 Espoo, Finland. E-mail: tekla.tammelin@vtt
bMolecular Plant Biology, Department of Life Technologies, University of Turku, Turku, FI-20014, Finland. E-mail: allahve@utu.fi
cDepartment of Bioproducts and Biosystems, Aalto University, Espoo FI-00076, Finland
First published on 20th April 2021
Abstract
Cell immobilization is a promising approach to create efficient photosynthetic cell factories for sustainable chemical production. Here, we demonstrate a novel photosynthetic solid-state cell factory design for sustainable biocatalytic ethylene production. We entrapped cyanobacteria within never-dried hydrogel films of TEMPO-oxidized cellulose nanofibers (TCNF) cross-linked with polyvinyl alcohol (PVA) to create a self-standing matrix architecture. The matrix is operational in the challenging submerged conditions and outperforms existing alginate-based solutions in terms of wet strength, long-term cell fitness, and stability. Based on rheological investigations, the critical strength of wet TCNF matrices is three times higher than in the existing immobilization matrices of alginate cross-linked with Ca2+. This is due to the rigid nature of the colloidal nanofiber network and the strong cross-linking with PVA, as opposed to polymeric alginate with reversible ionic Ca2+ bonds. The porous and hygroscopic nanofiber network also shields the cyanobacterial cells from environmental stress, maintaining photosynthetic activity during partial drying of films, and when submerged in the nutrient medium for long-term cultivation. Finally, TCNF matrices allow the ethylene-producing Synechocystis sp. PCC 6803 cells to operate in submerged conditions under high inorganic carbon loads (200 mM NaHCO3), where Ca2+-alginate matrices fail. The latter show severe cell leakage due to matrix disintegration already within 20 min of NaHCO3 supplementation. In contrast, TCNF-based matrices prevent cell leakage to the medium and restrict culture growth, leading to improved ethylene production yields. Furthermore, the operational capacity of the self-standing TNCF cell factory can be maintained long-term by periodically refreshing the nutrient medium. All in all, the results showcase the versatility and potential of cell immobilization with the never-dried colloidal TCNF matrix, paving the way towards novel biotechnological pathways using solid-state cell factories designed for efficient and sustainable production of e.g., monomers and fuels.
Introduction
Photosynthetic cell factories (PCFs) are versatile platforms promoting a circular bioeconomy, where photosynthetically active microbial cells of cyanobacteria and green algae are used as biocatalysts for sustainable production of targeted biochemicals and biofuels. These photosynthetic microbes use solar energy, water and trace mineral nutrients to create chemical energy by splitting water to O2 and assimilating atmospheric CO2 into biomass and energy-rich organic compounds from monomers to complex bioactive compounds.1,2 One such compound is ethylene, a major chemical building block and an attractive fuel source that is currently produced from fossil sources using energy-intensive steam-cracking, which generates significant amounts of greenhouse gases and toxic co-products.3,4To improve the production efficiency of traditional PCFs based on suspension culturing, many of its physiological and technical drawbacks can be overcome by immobilizing the photosynthetic cells, i.e. distributed within a thin layer of the solid or gel-like carrier matrix.5–8 This transition can improve light-to-product conversion efficiency by enabling more uniform irradiation of photosynthetic cells and restricting cell division, while simultaneously reducing water and energy consumption.9,10 Cell immobilization has been shown to increase the yield of ethylene production by 2-fold when compared to suspended cells, as well as the light-to-ethylene conversion efficiency by 3.5 times4 under non-submerged conditions. Using alginate cross-linked with divalent cations (e.g.Ca2+ and Ba2+) has been the conventional state-of-the-art solution for immobilizing green algae and cyanobacteria, despite its limitations in mechanical properties, such as wet strength and porosity.11–13 Alginate-based matrices are especially poorly suited for many environments with high ionic concentrations, such as wastewaters,6,14 as their ionic bonds are reversible depending on the concentration and species of the surrounding ions.6,15 Similarly, biocatalytic ethylene production with cyanobacteria presents especially challenging conditions, as Vajravel et al. (2020) reported that the optimal production requires the addition of 200 mM sodium bicarbonate (NaHCO3) as an inorganic carbon supplement, which leads to precipitation of Ca2+ ions as insoluble CaCO3, disrupting the Ca-cross-linking of alginate. Thus, a liquid-conveying intermediate was used to support the alginate matrix.4
Here, we present a novel solid-state cell factory matrix design exemplified by efficient biocatalytic ethylene production with entrapped cyanobacteria. The self-standing matrix architecture, which utilises never-dried hydrogel thin films from TEMPO-oxidized cellulose nanofibers (TCNF),16 is able to operate in challenging conditions of submerged cultivation. The synergistic advantages of this design are two-fold: firstly, it provides direct transportation of water, nutrient, gases and light for the cells. Secondly, the high mechanical stability of the nanoscale fibril network enables the cell factory to operate without any supporting structures, thus outperforming the proof-of-concept matrices we have presented earlier.13 The immobilization framework architecture is achieved by cross-linking the never-dried TCNF hydrogel with both chemical and ionic means using polyvinyl alcohol (PVA)17 and Ca2+,18 respectively. To evidence the improved mechanical performance, we employ thorough rheological investigations to reveal the viscoelastic behaviour of the wet hydrogel matrices. Previously, wet strength measurements have been only reported for dried and rewetted TCNF-PVA films,17 with several times higher values than for similar Ca2+-alginate films.15,19 Furthermore, we observe that submerged TCNF-based matrices remain stable during long-term ethylene production with the NaHCO3 supplemented conditions that lead to the disintegration of Ca2+-cross-linked alginate matrices.
In this paper, we showcase a novel solid-state cell factory matrix design based on never-dried colloidal TCNF network, surpassing the existing polymeric alternatives with respect to wet strength, long-term cell fitness and ethylene production yield. Nanocellulose-based matrix displays superior mechanical stability over conventionally utilised alginate systems. TCNF network fully prevented the cell leakage and matrix disintegration in challenging ethylene photoproduction conditions. These discoveries have the potential to revolutionize the cell factory concept by providing pathways to overcome the bottlenecks related to excessive water consumption, low volumetric productivity and inefficient light utilization.
Experimental
Materials
![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 g mol−1, DP 4300) used as a cross-linking additive to improve the wet strength of TCNF gels was purchased from Sigma-Aldrich and dissolved to a 5 wt% solution in Milli-Q water at 95 °C.
000 g mol−1, DP 4300) used as a cross-linking additive to improve the wet strength of TCNF gels was purchased from Sigma-Aldrich and dissolved to a 5 wt% solution in Milli-Q water at 95 °C.
Methods
 | ||
| Fig. 1 Concept for solid-state cell factories. The schematic representation of the experimental approach for immobilizing ethylene-producing mutant cells within TCNF- and alginate -based matrices. The appearance, AFM phase topography images, cross-linking mechanism and chemical structure of (A) colloidal TCNF network cross-linked with polyvinyl alcohol (PVA), and (B) dissolved alginate polymer chains with Ca2+-crosslinking are shown. (C) A representation of solid-state cell factory with the chemical formula for photosynthetic ethylene production by the immobilized mutant cells. The Z ranges for AFM images are 10 nm for TCNF and 2 nm for alginate, respectively. | ||
| Wet matrix | Cross-link | Water removed | Sample handling | |
|---|---|---|---|---|
| Experimental design #1 | 1% TCNF | Ca2+, PVA | 50% | Cut in circles (d = 5 cm) |
| 2% Alginate | Ca2+ | 0% | ||
| Experimental design #2 | 1% TCNF | Ca2+, PVA | 10–100% | Cut in ∼5 × 5 cm pieces |
| 1% Alginate | Ca2+ | 10–100% | ||
| Experimental design #3 | 1% TCNF | Ca2+, PVA | 50% | Cut in 3 × 1 cm strips |
| 1% Alginate | Ca2+ | 50% |
000 rpm for 1 minute. Air bubbles were removed via centrifugation at 3000g for 5 minutes, and the hydrogels were spread onto a Teflon film using a handheld coating applicator with a 2 mm gap at high speed to achieve sufficient shear forces for even spreading. Cross-linking with PVA was induced by evaporating water in a controlled temperature of 23 °C and relative humidity (RH) of 50%, until 50 wt% of water was evaporated from the samples, resulting in a final TCNF concentration of 1 wt%. Gelation of TCNF with calcium ions was induced by spraying of 50 mM CaCl2 onto the samples until they were fully wetted and waiting for 15 minutes before dipping the samples in a bath of 50 mM CaCl2 for additional 15 minutes to ensure full gelation. The hydrogel films were cut into circular pieces (d = 5 cm) and allowed to swell in a Petri dish filled with BG11 medium at low light for 24 hours prior to measurements. Control samples with BG11 medium without cells were prepared in the same way.
 | ||
| Fig. 2 Visual appearance of the never-dried hydrogel films containing immobilized cyanobacterial cells. (A) Self-standing PVA-Ca2+-TCNF, and (B) Ca2+-alginate. | ||
For preparation of Ca2+-alginate assemblies, WT cells in a suspension of BG11 medium were entrapped within alginate hydrogel similarly to preparation of Ca2+-TCNF-PVA, except 2 wt% alginate was used and no PVA was added. The formulation of cells and alginate was spread onto a Teflon film using a pipette until a thickness of 1 mm was achieved. The water evaporation used for PVA-Ca2+-TCNF was not applied, resulting in a final concentration of 1 wt%. Control samples with BG11 medium without cells were prepared in the same way.
δ) represents the ratio of loss to storage modulus (G′′/G′), and critical stress signifies the onset at which irreversible plastic deformation occurs. The stress sweep tests were performed between 0.05 and 200 Pa under a constant frequency of 0.1 Hz, and the frequency sweep tests (between 0.05 and 100 Hz at a constant stress of 0.1 Pa. All samples were allowed to relax for 2 min under the measuring head prior to measurements.
Results and discussion
Wet strength of immobilization matrices with rheology measurements
Small deformation oscillatory stress sweeps and frequency sweeps (ESI†) were performed on PVA-Ca2+-TCNF and Ca2+-alginate hydrogel films prepared as described for Experimental Design #1, both with and without WT cells. The elastic modulus (G′), viscous modulus (G′′) and tan δ values (G′′ divided by G′) of typical stress sweep measurement for each sample are presented in Fig. 3. As the figure shows, the average G′ values in the linear viscoelastic region for both PVA-Ca2+-TCNF and Ca2+-alginate samples are approximately an order of magnitude higher than G′′, indicating the systems are predominantly elastic (solid-like). However, they reach a crossover point where G′′ surpasses G′ after the onset of non-linear behaviour at critical stress, or yield stress, where the system becomes predominantly viscous. Similarly, frequency sweep measurements (Fig. S1†) show a linear behaviour for all samples under time-dependent stress until the crossover point at around 50 Hz. After this, the system is no longer able to reliably measure the samples and the values begin to change beyond the sensitivity limits of the device. For all samples, the cross-over is seen at similar frequencies, indicating that all gels are similarly stable under oscillatory frequency, and unaffected by time-dependent small deformation stress.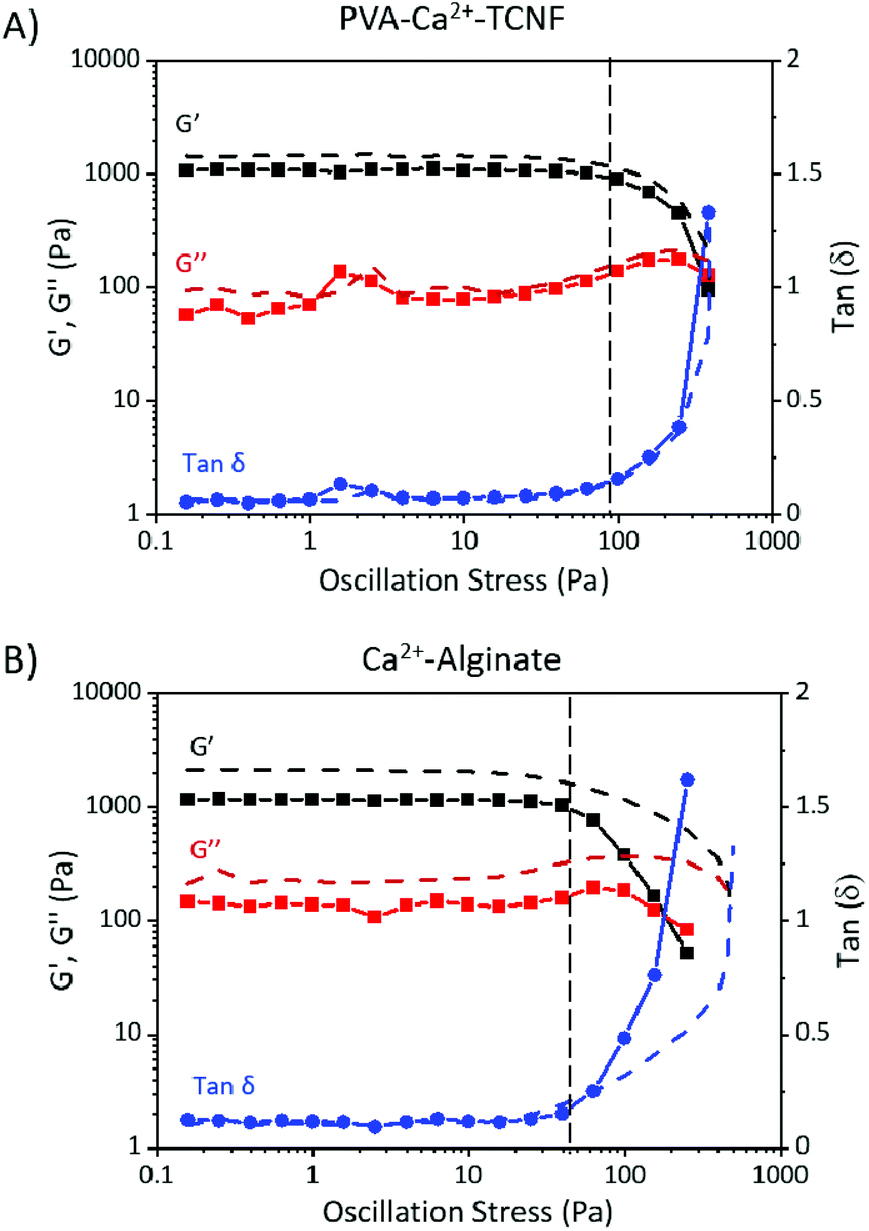 | ||
| Fig. 3 Small deformation oscillatory rheology of matrix hydrogel films. Typical stress sweeps of (A) PVA-Ca2+-TCNF and (B) Ca2+-alginate hydrogel films with (solid lines) and without (dotted lines) Synechocystis sp. PCC6803 wild-type (WT) cells, showing elastic moduli (G′, black square), viscous moduli (G′′, red square) and tan (δ) (blue circle). The dashed vertical line marks the onset of critical stress, where the material begins to flow. | ||
Fig. 4A shows a comparison of the tanδ values between typical stress sweeps of PVA-Ca2+-TCNF and Ca2+-alginate hydrogel films with and without WT cells. In the linear region, the tanδ values are ∼0.07 Pa for PVA-Ca2+-TCNF and ∼0.12 Pa for Ca2+-alginate. Moreover, critical stress occurs significantly later for PVA-Ca2+-TCNF than for Ca2+-alginate. As shown in Fig. 4B, the average G′ and G′′ values in the linear viscoelastic region for PVA-Ca2+-TCNF with WT cells are ∼1000 Pa and ∼75 Pa, respectively, and the average critical stress is ∼70 Pa. For the same system without cells, G′ increases to ∼1300 Pa while G′′ remains relatively unchanged, and critical stress increases to ∼75 Pa. For Ca2+-alginate with WT cells, the G′ and G′′ values are ∼1200 Pa and 160 Pa, respectively, and critical stress is around 20 Pa. Without cells, G′ and G′′ are significantly higher, with values of 1600–2200 Pa, and ∼220 Pa, respectively. Similarly, the critical stress increases to ∼40 Pa.
 | ||
| Fig. 4 Comparison of rheological properties of Synechocystis sp. PCC6803 s wild-type (WT) cells immobilized within PVA-Ca2+-TCNF and Ca2+-alginate hydrogel films. (A) Tan δ values during stress sweep. (B) Average values for elastic moduli (G′, black square), viscous moduli (G′′, red circle) and the critical stress (blue diamond) values of PVA-Ca2+-TCNF and Ca2+-alginate hydrogel films with and without the cells. | ||
Mechanical wet-strength testing of cross-linked TCNF and alginate gels and films is often carried out via either large-deformation tensile strength tests,17,24,25 rheological measurements,26,27 or a combination of both.28–30 However, tensile tests are predominantly performed with once-dried films, and therefore do not suit the never-dried hydrogel films demonstrated here. Nonetheless, there is literature for wet tensile strength measurements using TCNF-PVA and Ca2+-alginate in different conditions. Hakalahti et al.17 reported tensile strength of approximately 30 MPa for dried TCNF-PVA -films that had been immersed in water for 24 hours, whereas Sikareepaisan et al.15 and Pereira et al.19 reported values ranging from 3 to 9 MPa for Ca2+-alginate films immersed in water for 30 seconds before the measurement. Here, we use small-deformation rheological measurements to reveal the viscoelastic materials properties and internal interactions of self-standing never-dried PVA-Ca2+-TCNF and Ca2+-alginate hydrogel films in the wet state.
The relatively similar G′, G′′ and critical stress values of PVA-Ca2+-TCNF with and without WT cells (Fig. 3 and 4B) demonstrate that their viscoelastic properties are mostly independent of the cells, at least within the time scale of this experiment (∼24 hours). These results indicate that the cells are not strongly interacting with TCNF but passively entrapped within the fibrillar network matrix. Moreover, the percolation of the rigid network is not largely affected by the presence of the cells. Conversely, the larger difference in these values in Ca2+-alginate with and without the cells suggests that the inclusion of cells influences significantly the interactions of the alginate polymer matrix. The cells might also disrupt the Ca2+ -bonding between alginate polymer chains by bonding competitively with the Ca2+-ions. Moreover, the larger variance in rheology results for Ca2+-alginate compared to PVA-Ca2+-TCNF is potentially caused by the less specific nature of the ionic Ca2+ bonding between carboxylates in alginate compared to the bridging between the hydroxyl/carboxyl groups of TCNF and PVA chains. Similarly, the higher elastic moduli of Ca2+-alginate hydrogels compared to PVA-Ca2+-TCNF may be due to interactions between Ca2+-alginate and the cells. However, as Fig. 4A shows, the tanδ values of Ca2+-alginate are higher compared to PVA-Ca2+-TCNF, indicating they are more viscous (liquid-like) of the two systems. Similarly, TCNF-based samples have significantly higher critical stress than Ca2+-alginate, and thus they tolerate a higher shear stress before the structure yields. These results are likely due to the more rigid structure of colloidal nanocellulose fibril network compared to more flexible polymeric alginate, and the higher strength of the cross-links between TCNF and PVA chains compared to the ionic bonds between alginate polymer chains and Ca2+.
As described by Hakalahti et al.,17,31 the wet strength of dried and rewetted TCNF films cross-linked with PVA is due to covalent cross-linking occurring between the carboxyl groups present in TCNF, and the hydroxyl groups of PVA via an esterification reaction. Our results suggest indirectly that the partial removal of water in the PVA-Ca2+-TCNF hydrogel films can induce a similar reaction to some degree, and a direct verification warrants further investigations.
Effect of matrix material and water content on long-term fitness of immobilized cells
To create water-stable PVA-Ca2+-TCNF and Ca2+-alginate hydrogel films, samples with immobilized WT cells were prepared by mixing and controlled evaporation of water (drying) as described in Experimental Design #2. Ten different samples were prepared for both materials, where 0–90% of the total water content was gradually reduced by evaporation at 10% intervals. The samples were placed into Petri dishes filled with 40 ml of BG-11 medium, and their photosynthetic activity (Y(II)) was monitored regularly for 9 weeks. Furthermore, a visual assessment was used to estimate the mechanical stability of the submerged hydrogel films.The photosynthetic activity (Y(II)) and the visual appearance of the hydrogel films with immobilized WT cells are presented in Fig. 5. As shown in the highlighted area of the figure, the Y(II) of the cells in all samples decreases after the evaporation of water, which indicates that the drying causes stress to the cells. The Y(II) values also decrease more for the Ca2+-alginate, except for samples with 20–30% water (Fig. 5D). However, the Y(II) values begin to quickly increase afterwards as the cells recover, reaching similar values as before the immobilization. Samples with 30–20% water had some unrecovered areas near the edges of the hydrogel films, but samples with 10–0% water did not recover from the drying (Y(II) = 0, Fig. S2†) and are excluded from the remaining discussion of the results. After 9 weeks, the photosynthetic activity remained higher for the cells entrapped within PVA-Ca2+-TCNF (Y(II) ∼0.37) than within Ca2+-alginate (Y(II) ∼0.31–0.35), suggesting that the polymeric alginate network does not protect the cells from environmental stress as well as the colloidal TCNF network.
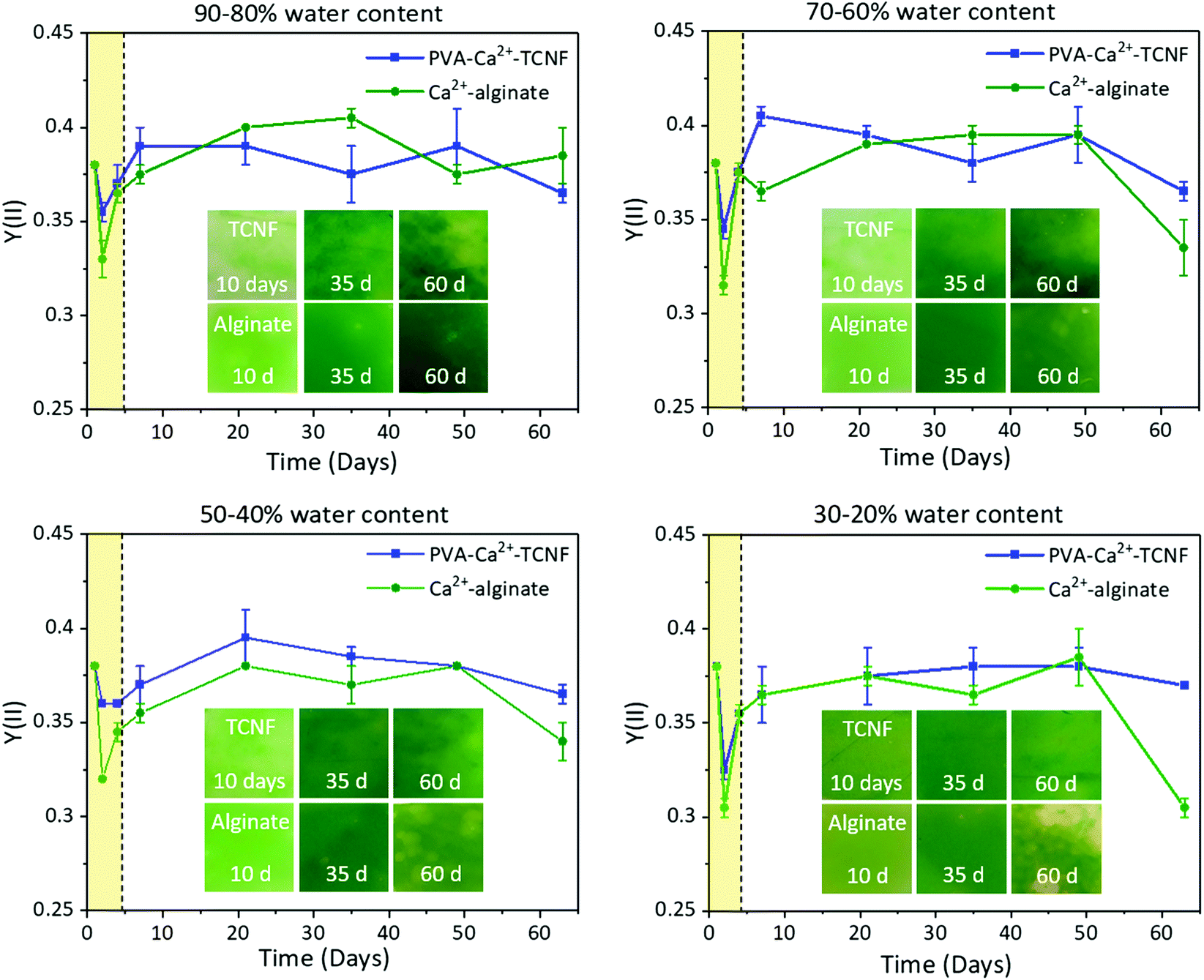 | ||
| Fig. 5 The long-term performance of Synechocystis sp. PCC6803 wild-type (WT) cells immobilized within PVA-Ca2+-TCNF and Ca2+-alginate. The samples were dried to 20–90% of original water content, at 10% intervals. Photosynthetic activity Y(II) and photographs are shown as a function of incubation time in a Petri dish filled with 40 ml of BG11 medium. The area highlighted with yellow colour and dotted line denote the period of drying, causing stress to the cells shown as a drop in Y(II), and the subsequent recovery after rewetting. | ||
Based on the visual appearance, PVA-Ca2+-TCNF hydrogel films with 70–20% water remained stable in submerged conditions throughout the experiment, with no remarkable differences in stability between them. In contrast, the PVA-Ca2+-TCNF with 80–90% remaining water appeared less stable in submerged conditions, with small pieces detaching from the gels (Fig. S3†). These results suggest that ∼30–60% of water should be removed to form a cross-link strong enough between TCNF and PVA and to improve the mechanical stability of the hydrogel films for cultivation in submerged conditions without causing the irreversible stress to cells. We have previously demonstrated a different method to prepare PVA-crosslinked TCNF films with immobilized cyanobacteria via drying the formulation at 70% RH for 22 hours, resulting in a structure that is stable on a water conveying support but not in submerged conditions.13 For Ca2+-alginate, all samples appeared to remain stable in submerged conditions throughout the experiment. However, the samples with 80–90% remaining water had higher Y(II) values than the ones with less water during the experiment, and especially after 8 weeks. Moreover, the samples with 20–30% of remaining water began to visibly bleach after 7 weeks, as shown in the photographs in Fig. 5. This indicates that drying of the Ca2+-alginate hydrogels creates stress to the cells and brings no additional benefits to the mechanical performance.
Ethylene production by immobilized cyanobacterial efe cells
Finally, we examined the effect of different immobilization matrices on genetically engineered ethylene-producing mutant cells under submerged conditions. The cells were immobilized within PVA-Ca2+-TCNF and Ca2+-alginate matrices as described in Experimental design #3, using a high optical cell density (∼2 OD750). The matrices with entrapped cells were placed in tightly sealed vials containing BG11 medium (pH 7.5). The medium was supplemented with 200 mM NaHCO3 as a Ci source. According to previous experiments by Vajravel et al., where NaHCO3 concentrations from 20 to 600 mM were tested, 200 mM was shown to be the optimal concentration for both the ethylene productivity and viability of the immobilized mutant cells in sealed vials.4 It is also worth noting that using CO2 as a replacement Ci source is unfeasible in this setup, as satisfying amount of Ci equivalent to 200 mM NaHCO3 would require saturating the cultivation media with 14% CO2, causing significant medium acidification. CO2 could alternatively be supplied to the system “on-demand” in accordance with its consumption by the culture. However, this system would require precise microprocessor-controlled monitoring of CO2 content in the bioreactor with feed-back regulation of CO2 supply, and as such is a topic for future efforts.However, within 20 minutes of the NaHCO3 addition, the submerged matrices of Ca2+-alginate were already mostly disintegrated, releasing cells into the medium, as seen in Fig. 6B and E. This is caused by the disruption of the coordination bonds between Ca2+ and alginate polymer chains, as Ca2+ ions are precipitated from the medium and alginate gels as insoluble CaCO3. As a result, a higher number of cells (∼11 μg of Chl) were in the medium than in the Ca2+-alginate matrix (∼4.8 μg of Chl) (Fig. 6B). An additional matrix formulation with a higher (2 wt%) alginate content was also tested, resulting in similar disintegration (Fig. S4D†). In contrast, the PVA-Ca2+-TCNF matrices remained stable throughout the experiment, and no Chl was detected in the medium (Fig. 6A). Additionally, Ca2+-alginate matrices with no external support are completely disintegrated within 60 minutes in these conditions, while self-standing PVA-Ca2+-TCNF matrices remain stable and applicable (Fig. S5†). According to these results, alginate-based matrices are not suitable for chemicals production process in the submerged system with NaHCO3 supplement, but TCNF-PVA-based matrices perform well under these conditions. Notwithstanding, high photosynthetic activity (Y(II) = 0.4) was detected for all samples (Fig. 6C). Thus, we continued with the production assay using the samples with a mixture of suspended and immobilized cells in the disintegrated Ca2+-alginate matrix (hereby referred as “destroyed Ca2+-alginate”) as a control.
 | ||
| Fig. 6 Ethylene producing capacity of the Synechocystis sp. PCC6803 efe cells entrapped within PVA-Ca2+-TCNF and Ca2+-alginate matrices under submerged production conditions. A, (B) Chl content and visual appearance of the films at 0–20 min, as well as Chl content at 7 and 8 days. (C) Photosynthetic activity. (D) Ethylene production yield calculated based on initial total Chl content. Ethylene production was initiated with an addition of IPTG (1 mM) and NaHCO3 (200 mM) in sealed vials containing 3 ml of BG11 medium (pH 7.5). (E) Visual appearance of the entrapped cells in self-standing PVA-Ca2+-TCNF hydrogel film matrices, as well as in destroyed Ca2+-alginate matrices. Each experimental point represents at least 3 independent measurements from 3 vials. The error bar represents the mean of all replicates (±SD). | ||
As shown in Fig. 6D, all samples were able to produce ethylene for more than 7 days. The cells entrapped in PVA-Ca2+-TCNF produced ∼180 μmol of C2H4 (mg Chl)−1, whereas the cells in Ca2+-alginate produced only ∼130 μmol C2H4 (mg Chl)−1. After 7 days of cultivation, the photosynthetic activity (Y(II)) of cells in the PVA-Ca2+-TCNF was ∼0.32, whereas the photosynthetic activity of cells in destroyed Ca2+-alginate was ∼0.2. The CO2 uptake of the cells entrapped in PVA-Ca2+-TCNF was also 1.4-times higher compared to the cells in destroyed Ca2+-alginate (Fig. S4A†). These results indicate that cell immobilization within the PVA-Ca2+-TCNF matrix improves their photosynthetic performance and long-term production phase. Furthermore, the final total Chl content of cells in the PVA-Ca2+-TCNF matrix increased ca. two-fold during 7 days (Fig. 6A), whereas the total Chl content of the cells in vials with destroyed Ca2+-alginate was ∼2.5-times higher (Fig. 6B). This difference indicates that the growth of the cells entrapped in PVA-Ca2+-TCNF matrices is restricted as compared to the partially suspended cells in destroyed Ca2+-alginate. The growth restriction might allow the cell metabolism to shift more towards ethylene production.
At the end of the stationary phase of ethylene production (at 8 days), the photosynthetic activity of all cells had substantially decreased in all samples due to their prolonged exposure to a closed system (Fig. 6C). However, a bleaching effect was observed only in the cells in destroyed Ca2+-alginate, which is also reflected in their decreased Chl content at 8 days (Fig. 6B).
For the long-term operational capacity of the system, the cells must maintain high levels of photosynthetic activity after periodic refreshment of medium nutrients and Ci source. In our multi-cycle photosynthetic activity monitoring experiment in closed conditions and in the presence of 200 mM NaHCO3, WT cells entrapped within PVA-Ca2-TCNF retained high photosynthetic activity after two refreshment cycles. In contrast, the operational capacity of the destroyed Ca2+-alginate was lost after the first refreshment cycle (Fig. S6†). Overall, our results indicate that the cells entrapped in PVA-Ca2+-TCNF matrices can be subjected to a long-term production phase due to high mechanical properties of the colloidal cross-linked fibril matrix, which protects the cells and enables good fitness even in submerged conditions with NaHCO3 supplement.
Conclusions
We fabricated a novel immobilization matrix design for photosynthetic cell factories with superior performance in mechanical strength, long-term cell fitness, matrix stability and ethylene productivity in challenging submerged conditions, when compared with the existing alginate-based alternatives.The matrix utilises never-dried TCNF cross-linked with PVA and Ca2+ (PVA-Ca2+-TCNF). After partial drying, PVA-Ca2+-TCNF forms self-standing hydrogel films with higher wet-strength than in Ca2+-alginate films. This is due to the colloidal nature of TCNF and their ability to form strong cross-links with PVA in contrast to the reversible ionic bonding between polymeric alginate and Ca2+. Observed via rheology, the critical stress of wet PVA-Ca2+-TCNF is over three times higher than in Ca2+-alginate. Moreover, cyanobacterial cells immobilised in PVA-Ca2+-TCNF maintain optimal photosynthetic activity during partial drying and demonstrate faster cell recovery from stress caused by drying. Importantly, improved photosynthetic activity of immobilised cyanobacterial cells is also observed in PVA-Ca2+-TCNF with low initial moisture content when submerged in cultivation medium due to the mechanical stability and hygroscopicity of the TCNF network. These results may also suggest that the TCNF matrix has more favourable porosity compared to alginate and investigating this in more detail is a topic for future efforts. Finally, PVA-Ca2+-TCNF matrices remained stable under submerged cultivation for ethylene production in the presence of NaHCO3 supplement with no cell leakage, while the Ca2+-alginate matrices disintegrated within 20 minutes, causing most of the cells to leak into the medium. Thus, the ethylene-producing Synechocystis sp. PCC 6803 cells immobilized within PVA-Ca2+-TCNF demonstrated improved photosynthetic activity with restricted growth and higher ethylene productivity throughout the experiment than the cells in the destroyed Ca2+-alginate. These results highlight the versatility and potential of this immobilization matrix design for long-term chemicals production.
Conflicts of interest
There are no conflicts to declare.Acknowledgements
This research was financially supported by the NordForsk NCoE program “NordAqua” (project 82845), the Academy of Finland (AlgaLEAF, project no. 322752, 322754 and 322755). This project has received funding from the European Union's Horizon 2020 research and innovation programme under grant agreement No 899576. The work is a part of the Academy of Finland's Flagship Programme under Projects No. 318890 and 318891 (Competence Center for Materials Bioeconomy, FinnCERES). Dr Suvi Arola was supported by the Academy of Finland postdoctoral fellow grant no 311608 and 326262. We are thankful to Dr Valentina Guccini for help in AFM imaging.References
- A. Hitchcock, C. N. Hunter and D. P. Canniffe, Microbiol. Biotechnol., 2020, 13, 363–367 CrossRef PubMed.
- R. Miao, H. Xie, X. Liu, P. Lindberg and P. Lindblad, Curr. Opin. Chem. Biol., 2020, 59, 69–76 CrossRef CAS PubMed.
- J. Ungerer, L. Tao, M. Davis, M. Ghirardi, P. C. Maness and J. Yu, Energy Environ. Sci., 2012, 5, 8998–9006 RSC.
- S. Vajravel, S. Sirin, S. Kosourov and Y. Allahverdiyeva, Green Chem., 2020, 22, 6404–6414 RSC.
- M. C. Flickinger, J. L. Schottel, D. R. Bond, A. Aksan and L. E. Scriven, Biotechnol. Prog., 2007, 23, 2–17 CrossRef CAS PubMed.
- L. E. de-Bashan and Y. Bashan, Bioresour. Technol., 2010, 101, 1611–1627 CrossRef CAS PubMed.
- A. Léonard, P. Dandoy, E. Danloy, G. Leroux, C. F. Meunier, J. C. Rooke and B. L. Su, Chem. Soc. Rev., 2011, 40, 860–885 RSC.
- S. Vasilieva, E. Lobakova and A. Solovchenko, in Environmental Biotechnology Vol. 3, Springer, Cham, 2021, pp. 193–220. Search PubMed.
- S. Kosourov, G. Murukesan, M. Seibert and Y. Allahverdiyeva, Algal Res., 2017, 28, 253–263 CrossRef.
- S. N. Kosourov, M. He, Y. Allahverdiyeva and M. Seibert, Microalgal hydrogen production: achievements and perspectives, 2018, 16, 355–383 CAS.
- S. N. Kosourov and M. Seibert, Biotechnol. Bioeng., 2009, 102, 50–58 CrossRef CAS PubMed.
- H. Leino, S. N. Kosourov, L. Saari, K. Sivonen, A. A. Tsygankov, E. M. Aro and Y. Allahverdiyeva, Int. J. Hydrogen Energy, 2012, 37, 151–161 CrossRef CAS.
- M. Jämsä, S. Kosourov, V. Rissanen, M. Hakalahti, J. Pere, J. A. Ketoja, T. Tammelin and Y. Allahverdiyeva, J. Mater. Chem. A, 2018, 6, 5825–5835 RSC.
- N. Mallick and L. C. Rai, World J. Microbiol. Biotechnol., 1994, 10, 439–443 CrossRef CAS PubMed.
- P. Sikareepaisan, U. Ruktanonchai and P. Supaphol, Carbohydr. Polym., 2011, 83, 1457–1469 CrossRef CAS.
- T. Saito, Y. Nishiyama, J. L. Putaux, M. Vignon and A. Isogai, Biomacromolecules, 2006, 7, 1687–1691 CrossRef CAS PubMed.
- M. Hakalahti, A. Salminen, J. Seppälä, T. Tammelin and T. Hänninen, Carbohydr. Polym., 2015, 126, 78–82 CrossRef CAS PubMed.
- H. Dong, J. F. Snyder, K. S. Williams and J. W. Andzelm, Biomacromolecules, 2013, 14, 3338–3345 CrossRef CAS PubMed.
- R. Pereira, A. Carvalho, D. C. Vaz, M. H. Gil, A. Mendes and P. Bártolo, Int. J. Biol. Macromol., 2013, 52, 221–230 CrossRef CAS PubMed.
- K. Thiel, E. Mulaku, H. Dandapani, C. Nagy, E. M. Aro and P. Kallio, Microb. Cell Fact., 2018, 17, 34 CrossRef PubMed.
- R. Rippka, J. Deruelles and J. B. Waterbury, Microbiology, 1979, 111, 1–61 Search PubMed.
- M. Hakalahti, M. Faustini, C. Boissière, E. Kontturi and T. Tammelin, Biomacromolecules, 2017, 18, 2951–2958 CrossRef CAS PubMed.
- H. K. Lichtenthaler, Methods Enzymol., 1987, 148, 350–382 CAS.
- J. L. Drury, R. G. Dennis and D. J. Mooney, Biomaterials, 2004, 25, 3187–3199 CrossRef CAS PubMed.
- J. W. Rhim, LWT – Food Sci. Technol., 2004, 37, 323–330 CrossRef CAS.
- B. T. Stokke, K. I. Draget, O. Smidsrød, Y. Yuguchi, H. Urakawa and K. Kajiwara, Macromolecules, 2000, 33, 1853–1863 CrossRef CAS.
- H. Park, S. W. Kang, B. S. Kim, D. J. Mooney and K. Y. Lee, Macromol. Biosci., 2009, 9, 895–901 CrossRef CAS PubMed.
- S. E. C. Whitney, M. G. E. Gothard, J. T. Mitchell and M. J. Gidley, Plant Physiol., 1999, 121, 657–663 CrossRef CAS PubMed.
- B. A. Harper, S. Barbut, L. T. Lim and M. F. Marcone, J. Food Sci., 2014, 79, E562–E567 CrossRef CAS PubMed.
- A. Walther, F. Lossada, T. Benselfelt, K. Kriechbaum, L. Berglund, O. Ikkala, T. Saito, L. Wågberg and L. Bergström, Biomacromolecules, 2020, 21, 2536–2540 CrossRef CAS PubMed.
- M. Hakalahti, A. Mautner, L. S. Johansson, T. Hänninen, H. Setälä, E. Kontturi, A. Bismarck and T. Tammelin, ACS Appl. Mater. Interfaces, 2016, 8, 2923–2927 CrossRef CAS PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d1gc00502b |
| ‡ These authors contributed equally. |
| This journal is © The Royal Society of Chemistry 2021 |