
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceProphylactic treatment of curcumin in a rat model of depression by attenuating hippocampal synaptic loss†
Cuiqin
Fan
a,
Ye
Li
a,
Tian
Lan
a,
Wenjing
Wang
a,
Xueqin
Mao
b and
Shu Yan
Yu
 *ac
*ac
aDepartment of Physiology, School of Basic Medical Sciences, Cheeloo College of Medicine, Shandong University, Jinan, Shandong 250012, China. E-mail: shuyanyu@sdu.edu.cn; Tel: +86-0531-88383902; Fax: +86-0531-88382502
bDepartment of Psychology, Qilu Hospital of Shandong University, 107 Wenhuaxilu Road, Jinan, Shandong Province 250012, PR China
cShandong Provincial Key Laboratory of Mental Disorders, School of Basic Medical sciences, Cheeloo College of Medicine, Shandong University, Jinan, Shandong 250012, China
First published on 6th October 2021
Abstract
Curcumin is a polyphenol substance considered to be effective in the treatment of a number of neurodegenerative diseases. However, details regarding the exact mechanisms for the protective effects of curcumin in neuropsychiatric disorders, like depression, remain unknown. In the pathogenesis of major depressive disorder (MDD) it appears that dysregulation of oxidative stress and immune systems, particularly within the hippocampal region, may play a critical role. Here, we show that pre-treatment with curcumin (40 mg kg−1) alleviates depression-like behaviors in a LPS-induced rat model of depression, effects which were accompanied with suppression of oxidative stress and inflammation and an inhibition of neuronal apoptosis in the hippocampal CA1 region, and results from ultramicrostructure electrophysiological experiments revealed that the curcumin pre-treatment significantly prevented excessive synaptic loss and enhanced synaptic functioning in this LPS-induced rat model. In addition, curcumin attenuated the increases in levels of miR-146a-5p and decreases in the expression of p-ERK signaling that would normally occur within CA1 regions of these depressed rats. Taken together, these results demonstrated that curcumin exerts neuroprotective and antidepressant activities by suppressing oxidative stress, neural inflammation and their related effects upon synaptic dysregulation. One of the mechanisms for these beneficial effects of curcumin appears to involve the miR-146a-5p/ERK signaling pathway within the hippocampal CA1 region. These findings not only elucidated some of the mechanisms underlying the neuroprotective/antidepressant effects of curcumin, but also suggested a role of curcumin as a potential therapeutic strategy for depression.
Introduction
Major depressive disorder (MDD), a high-risk neuropsychiatric condition well known for its slow progression, variety of symptoms and unpredictable course/prognosis, greatly restricts psychosocial functioning and severely lowers personal quality of life.1 Although there has been some progress in the understanding of the pathophysiology of MDD and its treatment, a substantial proportion of patients (up to 27%) do not respond adequately, or at all, to the antidepressants currently available.2,3 Moreover, significant, often intolerable, side-effects can be associated with these drugs.4 Accordingly, a better understanding of the underlying pathogenesis of depression is urgently needed for the development of more effective treatments.1,5Recent evidence has accrued which suggests that depression is related to changes in neuronal plasticity and reductions in size and functioning of the frontal cortex and hippocampal area.6 A major cause of damaging alterations in brain structure and function involves oxidative stress (OS). With increases in brain OS, there is a subsequent activation of pro-inflammatory signaling pathways that then promote the pathological processes of depression.6–9 In this way, an excess of OS along with aberrations in inflammatory pathways represent two important components of the pathophysiology of depression.10 In patients with MDD, there is a significant decrease in superoxide dismutase (SOD) levels,11 while treatment with antidepressants tends to restore these decreased levels of SODs, in particular within the prefrontal cortex of these patients.12 The antioxidative enzymes, SOD are responsible for the spontaneous dismutation of superoxide radicals and generation of H2O2, which is detoxified by CAT and GSH peroxidase (GPX).13 Similarly, our findings that reactive oxygen species (ROS) are significantly elevated in a rat model of depression and are involved in neuronal injury, provided further evidence for the involvement of OS in depression.14,15 With regards to the role of inflammation in the etiology and deterioration of depression, there is evidence of microglial activation and neuroinflammation within post-mortem brains of depressed patients.16 These findings suggest that drugs which specifically target the oxidative and immune systems may prove an effective strategy for the treatment of depression. However, the exact mechanisms relating to oxidative stress and inflammation with depression have yet to be examined in detail.
Curcumin is the main natural polyphenol found in the rhizome of curcuma longa (turmeric) and in other curcuma spp.17 Beneficial effects of curcumin treatment have been widely reported for a variety of neurological diseases and cancer due to its formidable antioxidant and anti–inflammatory activities.18 As a result, a considerable number of studies have been directed toward understanding the mechanisms of action of curcumin and its activities against a variety of pathologies.17 And, in fact, there is a growing interest on the use of curcumin to prevent or delay the occurrence of neurodegenerative diseases, such as Alzheimer's disease (AD).18–20 Previous work in our laboratory has shown that one potential mechanisms underpinning the putative antidepressant effects of curcumin is the inhibition of expression of pro-inflammatory IL-1-beta in the mPFC of a CUMS (chronic unpredictable mild stress)-rat model for aspects of depression.21,22 However, alternative mechanisms such as the effects of curcumin on oxidative stress and the inflammatory response are as yet unexplored. Accordingly, in this study, we carried out a series of experiments to determine the extent to which these mechanisms contribute to the antidepressant-like effects of curcumin. In light of the findings indicating antioxidant and anti–inflammatory activities of curcumin as related to the pathogenesis of neurological diseases, we hypothesized that these antioxidant and anti–inflammatory effects may also enable curcumin to function as an agent capable of diminishing the display of depression-like behaviors. We found that in the LPS-induced rat model of depression pre-treatment with curcumin attenuated depression-like behaviors, effects which were associated with a suppression of oxidative stress and inflammation. Such processes can then alter synaptic structure and function via the miR-146a-5p/ERK signaling pathway.
Materials and methods
Animals
Male Wister rats (160–180 g body weight) were purchased from the Shandong University Experimental Animal Centre. All experiments were performed according to the National Institutes of Health Guide for the Care and Use of Laboratory animals (NIH Publications No. 8023, revised 1978) and were approved by the Shandong University Animal Care and Use Committee. The rats were maintained three/cage under a 12 h light–dark cycle with free access to food and water.Drugs
Curcumin (Sigma, USA) was dissolved in 0.1% dimethyl sulfoxide (DMSO, Sigma, USA) at a concentration of 10 mg ml−1. Rats received daily intraperitoneal injections of curcumin (40 mg kg−1) for 1 week at 60 min prior to LPS injection.23 The AAV9-U6-miR-146a-5p-inhibitor-CAG-eGFP virus (AAV-miR-146a-5p-inhibitor), AAV9-CMV-eGFP-miR-146a-5p virus (AAV-miR-146a-5p) and scrambled control were purchased from GENE (Shanghai, China) and were injected within the hippocampal CA1 region while rats were position in a stereotaxic apparatus.Rat model of depression and drug administration
Wistar male rats received daily intraperitoneal injections of LPS (0.5 mg kg−1, sigma, L-2880) dissolved in sterile 0.9% saline for one week to induce depression-like behaviors as described previously.24,25 This dosage was used to stimulate a subclinical infection without inducing obvious inflammation and other apparent impairments in animals. Curcumin (40 mg kg−1, Sigma, C-1386) was prepared at a concentration of 10 mg ml−1.26 Curcumin or LPS was injected between 10:00 and 11:00 a.m. daily for 7 days. In all experiments, Curcumin was administered intraperitoneally (i. p.) 60 min prior to LPS injection. The control group was untreated. Behavioral tests were performed on the day following their final LPS injection. The hippocampal tissue was dissected 24 h after the behavioural test for molecular experiment or electrophysiology experiments.Behavioral tests
All behavioral tests were conducted during the dark circadian period (19:00–24:00 for rats).Sucrose preference test (SPT)
The SPT was conducted as described previously with slightly modifications.24,27,28 In brief, rats were exposed to two bottles containing water for 2 days, followed by two bottles containing 1% sucrose for 2 days. After 24 h of food and water deprivation, rats were permitted free access to the two bottles with one containing water and the other 1% sucrose. The test duration was 2 h and was conducted in the dark, with bottle positions exchanged after 1 h. A sucrose preference was defined as the ratio of the consumption of sucrose solution to the total consumption of both the water and sucrose solution during the 2 h test period.Forced swim test (FST)
The FST was also conducted as described previously.24,29,30 Rats were individually placed in a cylinder of water (height: 80 cm, diameter: 30 cm, 25 °C) for an initial 15 min training session. At 24 h later the rat was again placed in the cylinder for a videotaped 5 min test session. An immobility score was recorded from each rat for this test period with immobility defined as floating or remaining motionless with only minimal motion expended to maintain their head above the water.Open field test (OPT)
The OPT was performed after all other behavioral tests were completed. Rats were placed in the center of an arena (100 × 100 × 40 cm) and permitted to move freely throughout the arena for 5 min. During this 5 min period, the movement of rat was tracked by a video camera and subsequently analyzed.Oxidative stress measurements
The activity of anti-oxidative enzymes in hippocampal CA1 tissue homogenates was estimated with use of the superoxide dismutase (SOD) activity (No. A001-3), catalase (CAT) activity (A007-1), total antioxidant capacity (T-AOC) activity (A015-2) and the lactic dehydrogenase (LDH) (A020-2) assay kits. The content of malondialdehyde (MDA) and nitric oxide (NO) were detected using the MDA (No. A003-1) and NO (No. A013-2) assay kits. All assay kits were obtained from Jiancheng Inc. (Nanjing, China) and all experimental protocols were according to the manufacturers’ guidelines.Exosome isolation and identification
Supernatants of microglial cultures and serum of rats were collected and exosomes were isolated with use of differential centrifugation.31 Briefly, cells and debris were cleared from sample with centrifugation at 300g for 10 min, then pelleted from the supernatant by centrifugation at 10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000g for 30 min. Exosomes were subsequently isolated from the supernatant following centrifugation at 100000g for 1 h and resuspended within 1 ml of filtered PBS with sizes being determined using a ZETASIZER Nano series-Nano-ZS (Malvern, England). The procedure for electron microscopy analysis of exosomes was according to that described previously with slight modifications.32 In brief, freshly purified exosomes were deposited onto a paraffin membrane and covered with a 150-mesh copper mesh for 45 min. They were then washed with PBS, before the copper mesh was fixed in 3% glutaraldehyde for 10 minutes and negatively stained with 2% uranyl acetate. Images of exosomes were obtained using a JEM-1011 electron microscope (JEOL, Japan) at an accelerating voltage of 80 kV.
000g for 30 min. Exosomes were subsequently isolated from the supernatant following centrifugation at 100000g for 1 h and resuspended within 1 ml of filtered PBS with sizes being determined using a ZETASIZER Nano series-Nano-ZS (Malvern, England). The procedure for electron microscopy analysis of exosomes was according to that described previously with slight modifications.32 In brief, freshly purified exosomes were deposited onto a paraffin membrane and covered with a 150-mesh copper mesh for 45 min. They were then washed with PBS, before the copper mesh was fixed in 3% glutaraldehyde for 10 minutes and negatively stained with 2% uranyl acetate. Images of exosomes were obtained using a JEM-1011 electron microscope (JEOL, Japan) at an accelerating voltage of 80 kV.
Transmission electron microscopy (TEM)
Samples of CA1 tissues (1 × 1 × 1 mm) were placed in 2.5% glutaraldehyde overnight at 4 °C before being fixed with 1% osmium tetroxide for 1 h. Tissues were then subjected to a graded ethanol dehydration series, followed by immersion in propylene oxide overnight and embedded with resin. After being cut into ultrathin sections (70 nm), tissues were stained with 4% uranyl acetate for 20 min and 0.5% lead citrate for 5 min. Micrographs were captured using transmission electron microscopy (Philips Tecnai 20 U-Twin, Holland) at an accelerating voltage of 120 kV. The number of synapses per unit volume and total number of presynaptic vesicles were calculated as described previously.33,34Electrophysiology
Hippocampal slice preparations and electrophysiological recordings were performed according to procedures described previously.35 For detailed experiential procedures, see ESI.†Virus injections
Rats were deeply anesthetized with 4% pentobarbital and positioned in a stereotactic frame (Stoelting, USA). The AAV virus (2.5 μl, ∼1012 infection units per ml) was bilaterally injected into the hippocampus CA1 region (coordinates from the bregma, −3.48 mm; medial/lateral, ±1.8 mm; dorsal/ventral, −2.55 mm) using an electric microinjection pump (Stoelting, USA) at a rate of 100 nl min−1. After the injection was completed, the micro syringe remained at the site for an additional 10 min and was then slowly withdrawn.Primary culture of microglia
Primary cultures of microglia were prepared from newborn P1 rat pups as described previously.36 In brief, brains from the animals (P0–P1 rat) were extracted and placed in an ice-cold DMEM medium with 4.5 g L−1 glucose. Meninges were carefully removed using forceps. The brain tissue was then trypsinized with 0.25% trypsin and DNase for 5–10 min at room temperature to obtain the cell suspension through a 70 μm cell strainer. The cells were seeded into poly-L-lysine-coated flasks with DMEM/F12 medium supplemented with 10% FBS. After culture for 10–14 days, microglial cells were isolated from mixed glial cultures by gently shaking the flasks for 30 min on an orbital shaker at 37 °C. Microglia were reseeded onto 24-well plates and used for experiments.Additional assays/procedures
Golgi Staining, Immunofluorescence, western blotting, and Quantitative real-time polymerase chain reaction (PCR) were performed as described in the ESI.†Statistical analysis
The required sample sizes were estimated on the basis of our past experience performing similar experiments. Animals were randomly assigned to treatment groups. Behavioural analysis was performed blinded to experimental conditions. All data were presented as the means ± SEMs. A two-tailed Student's t-test was used for pairwise comparisons between two groups and a one-way ANOVA followed by Bonferroni's post-hoc test was used for comparisons involving multiple groups. All statistical analyses were performed using the GraphPad Prism software 8 program with a 95% confidence interval. A P < 0.05 was required for results to be considered statistically significant.Results
Curcumin attenuates depression-like behaviors and oxidative stress in LPS rats
A one-week treatment with lipopolysaccharide (LPS) was sufficient to produce depression-like behaviors in male Wistar rats as indicated by their performance in the SPT, FST and OPT (Fig. 1A). As shown in Fig. 1B and C, a significantly reduced intake of sucrose in SPT and markedly increased immobility times in the FST were observed in LPS compared to control rats. Responses which reflect anhedonia and behavioral despair. Moreover, no statistically significant differences were obtained between LPS and control rats with regard to locomotor activity in the OPT (Fig. 1D). Interestingly, pre-treatment with curcumin significantly increased sucrose consumption and decreased immobility times within LPS rats, suggesting that this treatment relieved the core symptom of depression in these depressed rats. | ||
| Fig. 1 Curcumin alleviates the core symptoms of depression and oxidative stress in hippocampal CA1 regions of LPS rats. (A) Experimental design: schematic figure of the treatment protocol. (B) Pretreatment with curcumin (40 mg kg−1) prevented the decreases in sucrose consumption in LPS rats in the SPT. (C) Pretreatment with curcumin reversed the increases in immobility times in LPS rats in the FST. (D) No statistically significant differences in locomotion were obtained among the groups in the OPT. (E–G) Activities of the antioxidant enzymes, CAT (E), SOD (F) and T-AOC (G) were analyzed within the hippocampal CA1 region. (H–J) Quantification of MDA (H), LDH (I) and NO (J) within the hippocampal CA1 region. (K) Western blot analysis showing levels of Nrf2 and HO-1 in the hippocampal CA1 region. (L and M) Representative images of 8-OHdG (L) and Mito-SOX (M) staining within the hippocampal CA1 region as indices for Mitochondria and DNA Oxidative Damage analysis. Scale bar: 20 μm (K) scale bar: 50 μm (L). N = 18 per group (B–D). N = 6 per group (E–M). The experiments were repeated at least three independently. *P < 0.05, **P < 0.01, ***P < 0.001, ns. No significance. One-way ANOVA with Bonferroni post-hoc analysis. | ||
The imbalance between OS and antioxidant defenses represent a major pathogenic basis for depression. To assess whether the antidepressant activity of curcumin may be due to alterations in oxidative stress levels, we first examined the levels of oxidative stress in these LPS rats. We found that activities of the antioxidant enzymes, CAT, SOD and T-AOC were all significantly decreased within the CA1 region of LPS rats (Fig. 1E–G), while contents of MDA, LDH and NO were all significantly increased (Fig. 1H–J). In contrast, the decreased expressions in the essential antioxidative nuclear transcription factor, Nrf2 and its product HO-1 (Fig. 1K), were observed in the hippocampal CA1 area of LPS rats, suggesting that oxidative stress does contribute to the pathological processes of depression. It has been reported that ROS attack nuclear and mitochondrial DNA, resulting in oxidization of nucleosides and mutagenic DNA lesions.37 Here, we show that mitochondrial superoxide levels, as measured using MitoSox (Fig. 1L), and 8-OHdG (Fig. 1M) were increased in the hippocampal CA1 region of LPS rats, results which indicate that oxidative damage to DNA by ROS was increased as a result of this LPS treatment. In contrast, pretreatment of curcumin suppressed all of the above oxidative stress changes induced by LPS exposure in rats, was associated with a significant downregulation of mitochondria and DNA oxidative damage in the hippocampal CA1 region of LPS rats.
Curcumin suppresses neuro-inflammatory responses in LPS rats
OS is intimately associated with inflammation, and dysfunction of mitochondria.38 Considering the key roles played by microglia in neuroinflammatory processes, we considered it important to examine microglial morphology and characteristics in these LPS rats. The microglia (CD11b and CD45) were significantly increased within hippocampal CA1 regions of LPS rats (Fig. 2A) and activated microglia within this site showed expanded cell bodies and shortened projections (Fig. 2B) Moreover, mRNA expression levels of some critical pro-inflammatory cytokines, such as interleukin-1β (IL-1β), interferon gamma (IFN-γ) and tumor necrosis factor-a (TNF-a), were all markedly upregulated in hippocampal CA1 regions of LPS rats as compared with controls (Fig. 2C). We also found that cyclo-oxygenase (COX)-2, whose expression is induced by ROS, was substantially increased in LPS rats (Fig. 2D). Notably, all of these responses resulting from LPS exposure were effectively reversed in LPS rats receiving a pre-treatment of curcumin. Together these results suggest that one of the mechanisms for the antidepressant effects of curcumin appears to involve a reduction in the increase of inflammatory responses associated with depression. | ||
| Fig. 2 Curcumin decreases neuro-inflammatory responses in LPS rats. (A) Western blot analysis of CD45 and CD11b protein expressions in the hippocampal CA1 region. (B) Representative images of morphological characteristics of Iba1+ microglia within the hippocampal CA1 region. Scale bar: 70 μm (white) and 40 μm (yellow). (C) RT-PCR results for the expressions of IL1β, TNF-α, and IFN-γ in the hippocampal CA1 region. (D) Western blot analysis showing change of COX-2 in the hippocampal CA1 region. N = 6 per group. The experiments were repeated at least three independently. *P < 0.05, **P < 0.01, ***P < 0.001, one-way ANOVA with Bonferroni post-hoc analysis. | ||
Curcumin decreases neuronal apoptosis within the hippocampal CA1 region of LPS rats
As excessive production of proinflammatory cytokines may cause neuronal death and cognitive changes, we next investigated whether the antidepressant activity of curcumin affects neuronal apoptosis within hippocampal CA1 regions. LPS-exposed rats displayed a notable increase in neuronal apoptosis, as indicated by increased densities of cleaved caspase 3 positive (cleaved caspase 3+/NeuN+) cells (Fig. 3A), and a greater number of apoptotic cells with nuclear chromatin margination, aggregation, and condensation as revealed from Hoechst-33258 staining (Fig. 3B). In addition, transcriptional levels of the apoptosis-related proteins, Bax and caspase 3, were significantly increased, within the CA1 region of LPS rats (Fig. 3C). Western blotting showed that hippocampal CA1 PARP, a protein senses in DNA strand breaks which contributes to their repair, was increased in LPS rats, and the apoptosis-related proteins BAK were increased, but Bcl-2 decreased, in LPS rats (Fig. 3D). With curcumin pre-treatment, the number of apoptotic cells and expressions of pro-apoptotic factors were significantly decreased in LPS rats. These findings demonstrate that curcumin suppresses neuronal apoptosis in LPS rats, which may then reveal another contributing process for its capacity to alleviate depression-like behaviors in LPS rats. | ||
| Fig. 3 Curcumin suppresses/reduces neuronal apoptosis within the hippocampal CA1 region of LPS rats. (A) Representative images of density of cleaved caspase 3+/NeuN+ within the hippocampal CA1 region. Scale bar: 50 μm. (B) Representative images of morphological changes/characteristics of nuclei by Hoechst-33258 staining in the hippocampal CA1 region. Scale bar: 5 μm. (C) RT-PCR results for mRNA expression levels of Bcl-2, and caspase-3 in the hippocampal CA1 region. (D) Western blot showing protein levels of PARP, Bcl-2, and BAK in the hippocampal CA1 region. N = 6 per group. The experiments were repeated at least three independently. *P < 0.05, **P < 0.01, one-way ANOVA with Bonferroni post-hoc analysis. | ||
Curcumin rescues synaptic loss and enhances synaptic function in the hippocampal CA1 region of LPS rats
OS in the brain modulates neuronal and extra neuronal signaling, producing neuronal inflammation, cell death and eventually synaptic loss. In this study, we found LPS rats had significantly fewer total synapses than controls (Fig. 4A) and exhibited a decrease in the number of synapses. Moreover, CA1 dendritic spine densities were strikingly reduced in LPS rats compared with control rats (Fig. 4B). Additional analyses revealed that expressions of synaptic proteins, including synaptophysin (Syn), postsynaptic density protein 95 (PSD-95), synaptotagmin1 (Syt1) and postsynaptic neuroligin1 (Nlg1) were all reduced in these LPS rats (Fig. 4C). These deficiencies in dendritic spines and synapses within CA1 neurons resulting from LPS exposure were rescued by curcumin pre-treatment. Next, whole-cell patch-clamp recordings were performed in acutely dissected hippocampal slices to investigated synaptic transmission efficiency within hippocampal CA1 neurons of LPS rats (Fig. S1†). In these LPS rats, we found that mEPSCs frequencies and amplitudes were largely decreased as compared with that of control rats (Fig. 4D), indicating a decrease in synaptic strength. Similarly, LPS rats also showed significantly reduced frequencies and amplitudes of spontaneous excitatory post-synaptic currents (sEPSCs) (Fig. 4E), as well as reduction in spontaneous tonic firing (Fig. 4F) in CA1 neurons and the number of docked vesicles located near presynaptic terminals was reduced in CA1 regions of LPS rats (Fig. 4G). Curcumin pre-treatment essentially reversed all of these deficits with increases in the frequencies and amplitudes of mEPSCs, sEPSCs, spontaneous tonic firing and number of docked vesicles being observed in the curcumin treated LPS rats. Altogether, these findings imply that curcumin may decrease the excessive synaptic loss and effectively normalize synaptic function in hippocampal CA1 regions to prevent the display of depression-like symptoms in LPS rats.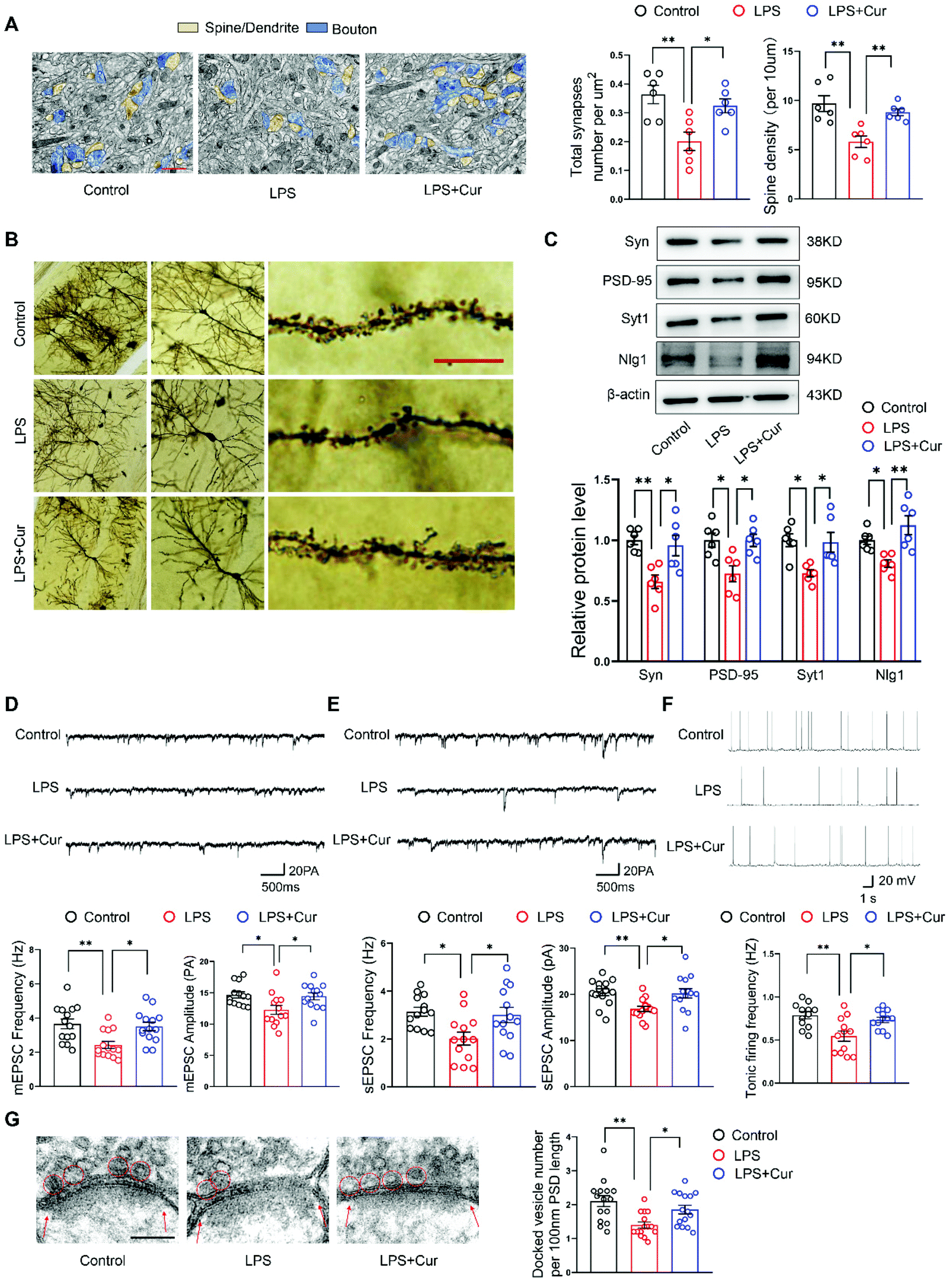 | ||
| Fig. 4 Curcumin increases synapse density and synaptic activity in the hippocampal CA1 region of LPS rats. (A) Representative electron micrographic images of CA1 neurons with yellow arrowheads indicating spine synapses. Scale bar: 1 μm. (B) Representative Golgi staining images of CA1 neurons with red arrowheads indicating dendritic spines. Scale bar: 10 μm. (C) Western blot analysis showing deregulation of neuroplasticity-related mediators Syn, PSD-95, Syt1 and Nlg1 protein in the hippocampal CA1 region of LPS rats. (D) Representative traces of mEPSCs from CA1 neurons and summary histogram showing mean frequencies and amplitudes of mEPSCs. (E) Example sEPSCs traces from CA1 neurons and summary histogram showing mean frequencies and amplitudes of sEPSCs. (F) Sample traces and frequencies of spontaneous tonic firing of CA1 neurons. (G) Representative TEM images of docked vesicles (in red circles). Red arrows show postsynaptic density (PSD). Scale bar: 100 nm. N = 6 per group (A–C). N = 13 neurons from 6 rats per group (D). N = 14 neurons from 6 rats per group (E). N = 12 neurons from 6 rats per group (F). N = 15 axon terminals from 6 rats per group (G). The experiments were repeated at least three independently. *P < 0.05, **P < 0.01, one-way ANOVA with Bonferroni post-hoc analysis. | ||
Curcumin attenuates miR-146a-5p overexpression and ERK activation in LPS rats
Finally, we wanted to evaluate some of the molecular mechanisms of curcumin in suppressing depression-like phenotypes. It had been reported that exosomes carrying miRNA were capable of exerting an acute impact upon neuroplasticity in the central nervous system.31 Accordingly, exosomes from rat serum, were assessed with electron microscopy (Fig. S2†), nanoparticle tracking analysis (Fig. 5A) and western blot analysis (Fig. 5B). We observed that miR-146a-5p was markedly increased in exosomes from the serum (Fig. 5C) and hippocampal CA1 regions (Fig. 5D) of LPS rats. These LPS-induced increases in miR-146a-5p at both sites were reversed by curcumin pre-treatment. Interestingly, miR-146a-5p, a microglia-specific miRNA, is not present in hippocampal neurons,31 Q-PCR analysis confirmed that an upregulation of miR-146a-5p was present in exosomes released from cultured inflammatory microglia (Fig. 5E and Fig. S3†). Further, decreases in the phosphorylation of ERK within CA1 regions of LPS rats were also significantly increased by curcumin pre-treatment, as determined with western blot (Fig. 5F).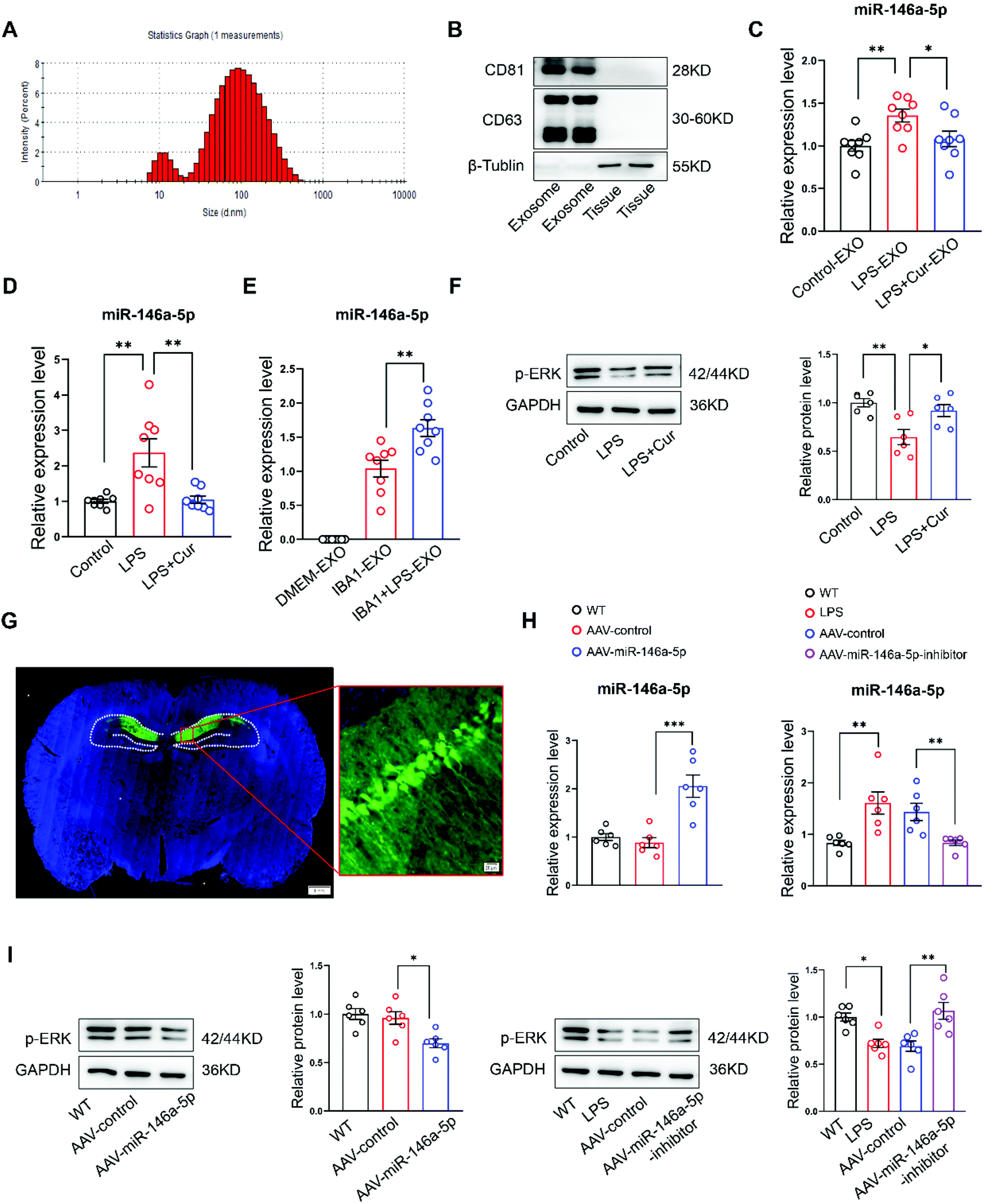 | ||
| Fig. 5 Curcumin inhibits LPS-induced miR-146a-5p overexpression and Erk activation. (A) Representative nanoparticle tracking analysis of exosomes. (B) Western blot to observe the expression of CD63 and CD81 in exosomes. (C) Representative q-PCR analysis of miR-146a-5p in exosomes released from rat serum. (D) Quantitative PCR showing expression of miR-146a-5p in the hippocampal CA1 region. (E) Relative quantification of miR-146a-5p in exosomes produced by microglia. (F) Western blot analysis of hippocampal CA1 regions showing downregulation of ERK phosphorylation levels in LPS rats. (G) Representative fluorescence image of the virus-infected slice. Scale bar: 1 mm. Scale bar: 20 μm. (H) Q-PCR analyzes the expression of miR-146a-5p after injection of AAV-miR-146a-5p-inhibitor or AAV-miR-146a-5p to the hippocampus CA1 of rats. (I) Western blot was used to analyze ERK phosphorylation levels within the hippocampal CA1 region of LPS rats with AAV-miR-146a-5p-sponge or control rats with AAV-miR-146a-5p injections. N = 6 per group. The experiments were repeated at least three independently. *P < 0.05, **P < 0.01, ***P < 0.001, one-way ANOVA with Bonferro. | ||
To follow up on these findings regarding miR-146a-5p, we examined the effects of injecting, AAV-miR-146a-5p-sponge or AAV-miR-146a-5p into the hippocampal CA1 region of LPS and control rats (Fig. 5G and Fig. S4†). These miR-146a-5p levels within CA1 regions were then analyzed with use of Q-PCR (Fig. 5H). The knock-down of miR-146a-5p within CA1 regions of LPS rats significantly increased phosphorylation levels of ERK, while phosphorylation levels of ERK were markedly reduced in control rats infected by AAV-miR-146a-5p (Fig. 5I). These data suggest that curcumin can exert its antidepressant effects, in part, through its capacity to initially suppress miR-146a-5p overexpression and ERK activation.
Discussion
Depression is regarded as a heterogeneous disease with unclear pathological mechanisms and inconsistent responses to treatment.39 To a large extent, the lack of a comprehensive understanding of this diseases’ mechanisms hinders the development of efficacious therapeutic approaches.36 Here, we showed that suppression of oxidative stress and neuroinflammation within hippocampal CA1 regions, mediates the antidepressant actions of curcumin in a rat model of depression. In these LPS injected rats, depression-like behaviors, including behavioral despair and anhedonia, are present. Pre-treatment with curcumin reversed these depression-like behaviors in LPS rats as well as decreased the accumulation of oxidative stress products and inflammatory factors in the CA1 region, which we believe to be key mechanisms responsible for inducing these robust antidepressant effects. In addition, CA1 neurons showed significant increases in synaptic activity and function in LPS rats receiving curcumin. Accordingly, our results suggest that curcumin quickly elevates mood by blocking oxidative stress and neuroinflammation activity of CA1 neurons thereby inhibiting excessive synaptic loss. These findings not only provided new insights into some of the pathological mechanisms involved with depression, but also indicated the potential for the development of curcumin as a new rapid-acting antidepressant.As a systemic administration of the cytokine inducer lipopolysaccharide (LPS) can produce clinical symptoms of depression in humans and rodents,25 this agent was selected as a means to generate a model of depression and thus provide an opportunity for assessing some of the pathological mechanisms involved with this condition. In specific, LPS treatment in rats has been shown to significantly reduce the percent of sucrose consumption in the SPT and increase immobility times in the FST, two notable indices of behavioral despair and anhedonia, which are core symptoms of depression. When these LPS rats received a pre-treatment of curcumin these depression-like behaviors were significantly alleviated. These findings are in accord with previous reports showing that curcumin possess considerable antidepressant activity as demonstrated in animal models of depression,40 but little is known about the possible mechanisms of these beneficial effects in the treatment of depression. The identification of new/possible processes and molecular targets of curcumin responsible for this antidepressant activity has the potential of generating novel insights that could guide future research in the treatment of depression. Therefore, the goal of this report was to examine some of the underlying mechanisms through which curcumin may exert these therapeutic effects in depression.
Reactive oxygen species (ROS) represent an important component of neurogenesis and neuronal activity and are essential for the growth and development of neurons. However, an imbalance between the production of ROS and antioxidant capacity, can contribute to a risk for depression.37 Notably, alterations in antioxidant capacity of aged mice seem to be highly localized to the hippocampus.41 In our study, we found that antioxidative enzyme activities were decreased in the hippocampal CA1 region of LPS rats but increased in LPS rats receiving curcumin treatment. In addition, increases in oxidative stress products and the antioxidative factor, Nrf2, were also reversed by curcumin in LPS rats. Augmented levels of OS damage neuronal DNA and induce inflammation and dysfunction of mitochondria,42 with increased levels of Mito-SOX and 8-OHdG, indicating amplified ROS activity as accompanying mitochondrial oxidative stress and DNA damage. Curcumin was effective in mitigating this oxidative DNA damage, suggesting that this curcumin-dependent reduction of oxidative stress plays a key role in protecting LPS-induced depression-like behaviors. Increased oxidative stress triggers multiple proinflammatory mediators, such as IL-1β, IFN-γ and TNF-a, leading to increases in the activity of proinflammatory pathways and other apoptotic mediators, such as Caspase-3, ultimately resulting in neuronal death and synaptic loss.37 Here, we found that LPS exposure induces neuronal inflammation and apoptosis in the hippocampal CA1 region of rats and exerts a profound impact on the loss of dendritic spines in CA1 neurons. However, treatment with curcumin inhibits glial cell activation, reduces inflammatory cytokines and attenuates neuronal apoptosis and synaptic loss in this LPS rat model of depression. Dendritic spine loss is accompanied with decreases in the density and strength of excitatory synapses, as indicated by reductions in mEPSC frequencies and amplitudes.31 Again, curcumin markedly enhanced these frequencies and amplitudes of mEPSCs within CA1 neurons in LPS rats, implicating that this treatment produced increases in presynaptic release and sensitivity or number of postsynaptic membrane transmitters. Our current results, together with previous findings from our laboratory,21,22 suggest that these mechanisms, which are affected by curcumin, reveal some of the basis for the neuroprotective and antidepressant-like effects of curcumin, as observed in these LPS rats.
An additional factor that can play a significant role in depression is that of microglia. Microglia are specialized immune cells in the brain, that perform central roles in immune surveillance and maintenance of brain homeostasis.43 Under stress or pathological conditions, microglia secrete inflammatory mediators that disrupt neuronal function and, in this way, promote the occurrence and development of depression. In fact, active microglia were present within the CA1 region of the LPS rats of our study. However, the possible mechanisms through which reactive microglia may produce synaptic defects are not fully understood. It has been reported that microglia can secrete exosomes carrying miRNA which are capable of exerting an acute impact upon adult neuroplasticity.31,44 In this study, we found that a specific microglial miRNA, miR-146a-5p, was upregulated in exosomes and CA1 regions of LPS rats. An overexpression or knock-down of miR-146a-5p in hippocampal CA1 regions markedly downregulated and upregulated the phosphorylation of ERK, which could then regulate the expression of key synaptic proteins.31 Interestingly, we found that the increased levels of miR-146a-5p and decreased phosphorylation of ERK as observed in LPS rats were reversed by curcumin treatment. When collating the findings of this report, our results reveal a neuroprotective mechanism underlying the anti-depressive effects of curcumin. Specifically, we report that curcumin mitigates synaptic losses resulting from brain oxidative stress and inflammation and these antidepressant effects of curcumin correlate with the release of microglia-enriched miRNAs and exosomes.
Results from previous studies have revealed that curcumin can interact and modulate a variety of molecular targets such as transcription factors, inflammatory cytokines and kinases.17,18,45 This array of effects suggests that the pharmacological basis for the antidepressant effects of curcumin may be diverse and complicated. It should be pointed out that the present study mainly investigated the preventive activity of pretreatment of curcumin on the LPS-induced behavioral and neurobiological changes in male rats. However, depression is a complex psychiatric disease involving potential interactions among gene-environment and gene-gender factors. Whether this mechanism was equally applicable to female rats have not been fully identified on account of the female rats usually experience the effects of estrous cycle. This study failed to consider gender factors, which could also influence the study results.
In conclusion, the findings presented here demonstrated the specific neuroprotective mechanism underlying the anti-depressant effect of curcumin. Specifically, through its capacity to reduce inflammation and oxidative stress, curcumin prevents synaptic loss along with modulating microglial responses, effects which culminate in diminishing depression-like behaviors in a rat model of depression. These results not only provide new insights into some of the mechanisms of curcumin, but also suggest the potential for its use as a therapeutic agent against depression.
Author contributions
Cuiqin Fan roles: Data curation, visualization, conceptualization, formal analysis; Ye Li roles: formal analysis, validation, software; Tian Lan and Wenjing Wang roles: methodology; Xueqin Mao roles: investigation, resources; ShuYan Yu roles: conceptualization, funding acquisition, project administration, resources, supervision, writing – original draft, writing – review & editing.Conflicts of interest
The authors declare that they have no conflict of interest.Acknowledgements
The authors declare no competing interests. This study was supported by grants to Shu Yan Yu from the National Natural Science Foundation of China (NSFC81873796; 82071513), the Natural Science Foundation of Shandong Province of China (ZR2020ZD25) and the Fundamental Research Funds of Shandong University (SDCA20191021, 6010119069). We thank the staff of the electronic microscope laboratory of the experimental center in the basic medical school for technical assistance.References
- G. S. Malhi and J. J. Mann, Depression, Lancet, 2018, 392, 2299–2312 CrossRef.
- J. Angst, A. Gamma, W. Rossler, V. Ajdacic and D. N. Klein, Long-term depression versus episodic major depression: Results from the prospective Zurich study of a community sample, J. Affect. Disord., 2009, 115, 112–121 CrossRef PubMed.
- L. Boschloo, R. A. Schoevers, A. T. F. Beekman, J. H. Smit, A. M. van Hemert and B. W. J. H. Penninx, The Four-Year Course of Major Depressive Disorder: The Role of Staging and Risk Factor Determination, Psychother. Psychosom., 2014, 83, 279–288 CrossRef PubMed.
- C. Moret, M. Isaac and M. Briley, Problems associated with long-term treatment with selective serotonin reuptake inhibitors, J. Psychopharmacol., 2009, 23, 967–974 CrossRef CAS PubMed.
- D. S. Bassett, K. E. Cullen, S. B. Eickhoff, M. J. Farah, Y. Goda, P. Haggard, H. L. Hu, Y. L. Hurd, S. A. Josselyn, B. S. Khakh, J. A. Knoblich, P. Poirazi, R. A. Poldrack, M. Prinz, P. R. Roelfsema, T. L. Spires-Jones, M. Sur and H. R. Ueda, Reflections on the past two decades of neuroscience, Nat. Rev. Neurosci., 2020, 21, 524–534 CrossRef PubMed.
- E. L. Belleau, M. T. Treadway and D. A. Pizzagalli, The Impact of Stress and Major Depressive Disorder on Hippocampal and Medial Prefrontal Cortex Morphology, Biol. Psychiatry, 2019, 85, 443–453 CrossRef PubMed.
- T. M. Michel, S. Frangou, D. Thiemeyer, S. Camara, J. Jecel, K. Nara, A. Brunklaus, R. Zoechling and P. Riederer, Evidence for oxidative stress in the frontal cortex in patients with recurrent depressive disorder - a postmortem study, Psychiatry Res., 2007, 151, 145–150 CrossRef CAS PubMed.
- T. M. Michel, S. Camara, T. Tatschner, S. Frangou, A. J. Sheldrick, P. Riederer and E. Grunblatt, Increased xanthine oxidase in the thalamus and putamen in depression, World J. Biol. Psychiatry, 2010, 11, 314–320 CrossRef PubMed.
- N. Bakunina, C. M. Pariante and P. A. Zunszain, Immune mechanisms linked to depression via oxidative stress and neuroprogression, Immunology, 2015, 144, 365–373 CrossRef CAS PubMed.
- M. Maes, P. Galecki, Y. S. Chang and M. Berk, A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro)degenerative processes in that illness, Prog. Neuropsychopharmacol. Biol. Psychiatry, 2011, 35, 676–692 CrossRef CAS PubMed.
- A. Bajpai, A. K. Verma, M. Srivastava and R. Srivastava, Oxidative stress and major depression, J. Clin. Diagn. Res., 2014, 8, CC04–CC07 Search PubMed.
- G. A. Behr, J. C. F. Moreira and B. N. Frey, Preclinical and Clinical Evidence of Antioxidant Effects of Antidepressant Agents: Implications for the Pathophysiology of Major Depressive Disorder, Oxid. Med. Cell. Longevity, 2012, 2012, 609421 Search PubMed.
- Y. Wang, R. Branicky, A. Noe and S. Hekimi, Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling, J. Cell Biol., 2018, 217, 1915–1928 CrossRef CAS PubMed.
- Y. Li, L. Y. Wang, P. Wang, C. Q. Fan, P. Zhang, J. Shen and S. Y. Yu, Ginsenoside-Rg1 Rescues Stress-Induced Depression-Like Behaviors via Suppression of Oxidative Stress and Neural Inflammation in Rats, Oxid. Med. Cell. Longevity, 2020, 2020, 2325391 Search PubMed.
- C. Q. Fan, Y. F. Long, L. Y. Wang, X. H. Liu, Z. C. Liu, T. Lan, Y. Li and S. Y. Yu, N-Acetylcysteine Rescues Hippocampal Oxidative Stress-Induced Neuronal Injury via Suppression of p38/JNK Signaling in Depressed Rats, Front. Cell. Neurosci., 2020, 14, 554613 CrossRef CAS PubMed.
- E. Setiawan, A. A. Wilson, R. Mizrahi, P. M. Rusjan, L. Miler, G. Rajkowska, I. Suridjan, J. L. Kennedy, V. Rekkas, S. Houle and J. H. Meyer, Role of Translocator Protein Density, a Marker of Neuroinflammation, in the Brain During Major Depressive Episodes, JAMA Psychiatry, 2015, 72, 268–275 CrossRef PubMed.
- M. Pulido-Moran, J. Moreno-Fernandez, C. Ramirez-Tortosa and M. C. Ramirez-Tortosa, Curcumin and Health, Molecules, 2016, 21(3), 264 CrossRef PubMed.
- A. Bhat, A. M. Mahalakshmi, B. Ray, S. Tuladhar, T. A. Hediyal, E. Manthiannem, J. Padamati, R. Chandra, S. B. Chidambaram and M. K. Sakharkar, Benefits of curcumin in brain disorders, Biofactors, 2019, 45, 666–689 CrossRef CAS PubMed.
- A. N. Begum, M. R. Jones, G. P. Lim, T. Morihara, P. Kim, D. D. Heath, C. L. Rock, M. A. Pruitt, F. Yang, B. Hudspeth, S. Hu, K. F. Faull, B. Teter, G. M. Cole and S. A. Frautschy, Curcumin structure-function, bioavailability, and efficacy in models of neuroinflammation and Alzheimer's disease, J. Pharmacol. Exp. Ther., 2008, 326, 196–208 CrossRef CAS PubMed.
- V. Sorrenti, G. Contarini, S. Sut, S. Dall'Acqua, F. Confortin, A. Pagetta, P. Giusti and M. Zusso, Curcumin Prevents Acute Neuroinflammation and Long-Term Memory Impairment Induced by Systemic Lipopolysaccharide in Mice, Front. Pharmacol., 2018, 9, 183 CrossRef PubMed.
- C. Q. Fan, Q. Q. Song, P. Wang, M. Yang, B. Liu and S. Y. Yu, Curcumin Protects Against Chronic Stress-induced Dysregulation of Neuroplasticity and Depression-like Behaviors via Suppressing IL-1 beta Pathway in Rats, Neuroscience, 2018, 392, 92–106 CrossRef CAS PubMed.
- C. Fan, Q. Song, P. Wang, Y. Li, M. Yang and S. Y. Yu, Neuroprotective Effects of Curcumin on IL-1beta-Induced Neuronal Apoptosis and Depression-Like Behaviors Caused by Chronic Stress in Rats, Front. Cell. Neurosci., 2018, 12, 516 CrossRef CAS PubMed.
- W. K. Bhutani, M. Bishnoi and S. K. Kulkarni, Anti-depressant like effect of curcumin and its combination with piperine in unpredictable chronic stress-induced behavioral, biochemical and neurochemical changes, Pharmacol. Biochem. Behav., 2009, 92, 39–43 CrossRef PubMed.
- Y. H. Cui, Y. Yang, Z. Y. Ni, Y. Y. Dong, G. H. Cai, A. Foncelle, S. S. Ma, K. N. Sang, S. Y. Tang, Y. Z. Li, Y. Shen, H. Berry, S. X. Wu and H. L. Hu, Astroglial Kir4.1 in the lateral habenula drives neuronal bursts in depression, Nature, 2018, 554(7692), 323–327 CrossRef CAS PubMed.
- M. Adzic, J. Djordjevic, M. Mitic, Z. Brkic, I. Lukic and M. Radojcic, The contribution of hypothalamic neuroendocrine, neuroplastic and neuroinflammatory processes to lipopolysaccharide-induced depressive-like behaviour in female and male rats: Involvement of glucocorticoid receptor and C/EBP-beta, Behav. Brain Res., 2015, 291, 130–139 CrossRef CAS PubMed.
- L. Zhang, T. Xu, S. Wang, L. Yu, D. Liu, R. Zhan and S. Y. Yu, NMDA GluN2B receptors involved in the antidepressant effects of curcumin in the forced swim test, Prog. Neuropsychopharmacol. Biol. Psychiatry, 2013, 40, 12–17 CrossRef CAS PubMed.
- R. D. Porsolt, G. Brossard, C. Hautbois and S. Roux, Rodent models of depression: forced swimming and tail suspension behavioral despair tests in rats and mice, Curr. Protoc. Neurosci., 2001, Chapter 8, Unit 8.10A, DOI:10.1002/0471142301.ns0810as14.
- X. Y. Lu, B. Ross, M. Sanchez-Alavez, E. P. Zorrilla and T. Bartfai, Phenotypic analysis of GalR2 knockout mice in anxiety- and depression-related behavioral tests, Neuropeptides, 2008, 42, 387–397 CrossRef CAS PubMed.
- K. Li, T. Zhou, L. J. Liao, Z. F. Yang, C. Wong, F. Henn, R. Malinow, J. R. Yates and H. L. Hu, beta CaMKII in Lateral Habenula Mediates Core Symptoms of Depression, Science, 2013, 341, 1016–1020 CrossRef CAS PubMed.
- C. H. Duman, L. Schlesinger, M. Kodama, D. S. Russell and R. S. Duman, A role for MAP kinase signaling in behavioral models of depression and antidepressant treatment, Biol. Psychiatry, 2007, 61, 661–670 CrossRef CAS PubMed.
- I. Prada, M. Gabrielli, E. Turola, A. Iorio, G. D'Arrigo, R. Parolisi, M. De Luca, M. Pacifici, M. Bastoni, M. Lombardi, G. Legname, D. Cojoc, A. Buffo, R. Furlan, F. Peruzzi and C. Verderio, Glia-to-neuron transfer of miRNAs via extracellular vesicles: a new mechanism underlying inflammation-induced synaptic alterations, Acta Neuropathol., 2018, 135, 529–550 CrossRef CAS PubMed.
- F. Marques-Garcia and M. Isidoro-Garcia, Protocols for Exosome Isolation and RNA Profiling, Methods Mol. Biol., 2016, 1434, 153–167 CrossRef CAS PubMed.
- J. H. Lee, J. Y. Kim, S. Noh, H. Lee, S. Y. Lee, J. Y. Mun, H. Park and W. S. Chung, Astrocytes phagocytose adult hippocampal synapses for circuit homeostasis, Nature, 2021, 590(7847), 612–617 CrossRef CAS PubMed.
- B. Minnich, H. Leeb, E. W. N. Bernroider and A. Lametschwandtner, Three-dimensional morphometry in scanning electron microscopy: a technique for accurate dimensional and angular measurements of microstructures using stereopaired digitized images and digital image analysis, J. Microsc., 1999, 195, 23–33 CrossRef CAS PubMed.
- N. Li, L. Cui, G. Song, L. Guo, H. Gu, H. Cao, G. D. Li and Y. Zhou, Adolescent Isolation Interacts With DISC1 Point Mutation to Impair Adult Social Memory and Synaptic Functions in the Hippocampus, Front. Cell. Neurosci., 2018, 12, 238 CrossRef PubMed.
- T. T. Tamashiro, C. L. Dalgard and K. R. Byrnes, Primary Microglia Isolation from Mixed Glial Cell Cultures of Neonatal Rat Brain Tissue, J. Vis. Exp., 2012,(66), e3814 Search PubMed.
- S. Bhatt, A. N. Nagappa and C. R. Patil, Role of oxidative stress in depression, Drug Discov. Today, 2020, 25, 1270–1276 CrossRef CAS PubMed.
- P. Jenner, Oxidative stress in Parkinson's disease, Ann. Neurol., 2003, 53, S26–S36 CrossRef CAS PubMed.
- R. H. Belmaker and G. Agam, Major depressive disorder, N. Engl. J. Med., 2008, 358, 55–68 CrossRef CAS PubMed.
- L. Zhang, T. Y. Xu, S. Wang, L. Q. Yu, D. X. Liu, R. Z. Zhan and S. Y. Yu, Curcumin produces antidepressant effects via activating MAPK/ERK-dependent brain-derived neurotrophic factor expression in the amygdala of mice, Behav. Brain Res., 2012, 235, 67–72 CrossRef CAS PubMed.
- K. A. Stebbings, H. W. Choi, A. Ravindra and D. A. Llano, The impact of aging, hearing loss, and body weight on mouse hippocampal redox state, measured in brain slices using fluorescence imaging, Neurobiol. Aging, 2016, 42, 101–109 CrossRef CAS PubMed.
- E. Tonnies and E. Trushina, Oxidative Stress, Synaptic Dysfunction and Alzheimer's Disease, J. Alzheimer's Dis., 2017, 57, 1105–1121 Search PubMed.
- J. Q. Zhang, P. J. Rong, L. J. Zhang, H. He, T. Zhou, Y. H. Fan, L. Mo, Q. Y. Zhao, Y. Han, S. Y. Li, Y. F. Wang, W. Yan, H. F. Chen and Z. L. You, IL4-driven microglia modulate stress resilience through BDNF-dependent neurogenesis, Sci. Adv., 2021, 7(12), eabb9888 CAS.
- E. McNeill and D. Van Vactor, MicroRNAs Shape the Neuronal Landscape, Neuron, 2012, 75, 363–379 CrossRef CAS PubMed.
- B. Kocaadam and N. Sanlier, Curcumin, an active component of turmeric (Curcuma longa), and its effects on health, Crit. Rev. Food Sci. Nutr., 2017, 57, 2889–2895 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d1fo02676c |
| This journal is © The Royal Society of Chemistry 2021 |