
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Alleviation of cognitive impairment by gut microbiota lipopolysaccharide production-suppressing Lactobacillus plantarum and Bifidobacterium longum in mice†
Dong-Yun
Lee
,
Yoon-Jung
Shin
,
Jeon-Kyung
Kim
 ,
Hyo-Min
Jang
,
Min-Kyung
Joo
and
Dong-Hyun
Kim
*
,
Hyo-Min
Jang
,
Min-Kyung
Joo
and
Dong-Hyun
Kim
*
Neurobiota Research Center, College of Pharmacy, Kyung Hee University, 26, Kyungheedae-ro, Dongdaemun-gu, Seoul 02447, Korea. E-mail: dhkim@khu.ac.kr
First published on 5th October 2021
Abstract
Bacterial lipopolysaccharide (LPS) is a risk factor for the outbreak of Alzheimer's disease. Therefore, we isolated Lactobacillus plantarum NK151 and Bifidobacterium longum NK173 from a human fecal bacteria collection, which inhibited Escherichia coli LPS production, and examined their effects on the Escherichia coli K1- or LPS-induced cognitive impairment in mice. Oral gavage of NK151, NK173, or their (4![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1) mixture (NKm) significantly alleviated Escherichia coli K1-induced cognitive impairment-like behaviors in the Y-maze and novel object recognition tasks. Their treatments decreased IL-1β, IL-6, and TNF-α expression and NF-κB+/Iba1+ and LPS+/Iba1+ cell populations in the hippocampus, while the brain-derived neurotrophic factor (BDNF)+/neuronal nuclei (NeuN)+ cell population and BDNF to proBNDF expression increased. They suppressed LPS-induced cognition impairment-like behaviors and neuroinflammation marker levels in the hippocampus. Treatment with them reduced Escherichia coli K1- or LPS-induced LPS and apolipoprotein E levels in the blood and inflammatory marker levels in the colon. Furthermore, treatment with them modulated fecal Proteobacteria, Bacteroidetes, and Verrucomicrobia populations. Of these gut bacteria, Bacteroidaceae, Odoribacteraceae, Lactobacillaceae, Bifidobacteriaceae, Rikenellaceae, Helicobacteraceae, and Deferribacteraceae are correlated with cognitive function and blood and fecal LPS levels. These findings suggest that NK151 and NK173 may alleviate cognitive impairment with colitis by upregulating NF-κB-mediated BDNF expression through the suppression of fecal and blood bacterial LPS levels.
:1) mixture (NKm) significantly alleviated Escherichia coli K1-induced cognitive impairment-like behaviors in the Y-maze and novel object recognition tasks. Their treatments decreased IL-1β, IL-6, and TNF-α expression and NF-κB+/Iba1+ and LPS+/Iba1+ cell populations in the hippocampus, while the brain-derived neurotrophic factor (BDNF)+/neuronal nuclei (NeuN)+ cell population and BDNF to proBNDF expression increased. They suppressed LPS-induced cognition impairment-like behaviors and neuroinflammation marker levels in the hippocampus. Treatment with them reduced Escherichia coli K1- or LPS-induced LPS and apolipoprotein E levels in the blood and inflammatory marker levels in the colon. Furthermore, treatment with them modulated fecal Proteobacteria, Bacteroidetes, and Verrucomicrobia populations. Of these gut bacteria, Bacteroidaceae, Odoribacteraceae, Lactobacillaceae, Bifidobacteriaceae, Rikenellaceae, Helicobacteraceae, and Deferribacteraceae are correlated with cognitive function and blood and fecal LPS levels. These findings suggest that NK151 and NK173 may alleviate cognitive impairment with colitis by upregulating NF-κB-mediated BDNF expression through the suppression of fecal and blood bacterial LPS levels.
1. Introduction
Dementia is a neurodegenerative disorder characterized by a progressive decline in behavioral and cognitive functions.1,2 The most common form of dementia in aged adults is Alzheimer's disease (AD). The pathogenesis of AD includes neuroinflammation, neurotransmitter imbalance, amyloid-beta (Aβ) aggregation, and neurofibrillary tangle formation in the central nervous system.2–4 Aβ aggregation and neuroinflammation are promoted by gut dysbiosis and bacterial lipopolysaccharide (LPS) overproduction.5,6 Blood LPS levels in AD patients are significantly higher than in healthy control.7The gut microbiota communicates with the brain through the microbiota–gut–brain (MGB) axis.6,8,9 Exposure to stressors such as antibiotics and immobilization causes gut dysbiosis and bacterial LPS overproduction.10,11 Excessive, chronic exposure to LPS induces inflammatory responses including NF-κB activation and tumor necrosis factor (TNF)-α and interleukin (IL)-6 expression in the peripheral and systemic immune cells, inducing apolipoprotein E (apoE) in the blood and suppressing brain-derived neurotropic factor (BDNF) expression in the brain.12–14 Brain-derived neurotrophic factor (BDNF) regulates the neuronal synaptic plasticity in the peripheral and central nervous system.15,16 Moreover, blood BDNF is lower in AD patients than in healthy individuals.17,18 ApoE, a principal cholesterol carrier, directly binds LPS in the blood and delivers it to the liver, which degrades LPS.19,20 TNF-α expression suppressing Lactobacillus mucosae NK41 alleviates Escherichia coli K1-induced cognitive impairment in mice by regulating NF-κB-mediated BDNF expression.21Lactobacillus plantarum C29 improves cognitive impairment in 5XFAD and aged mice by modulating NF-κB-mediated BDNF expression.22,23Bifidobacterium longum NK46, which inhibits gut bacterial growth in vitro and NF-κB activation in BV-2 cells, suppresses cognitive decline in 5XFAD transgenic and aged mice by suppressing microbiota LPS-mediated NF-κB activation.24Lactobacillus brevis OW38 alleviates cognitive decline in aged mice by inhibiting gut microbiota LPS production and suppressing p16 expression.25 These findings suggest that the suppression of BDNF expression by LPS-mediated NF-κB activation can cause cognitive impairment and reducing the overproduction of LPS by the gut microbiota and blood LPS level may be beneficial for the therapy of cognitive impairment.
The aim of the present study was to elucidate whether gut microbiota LPS production-suppressing probiotics could alleviate gut dysbiosis-induced cognitive impairment. Therefore, we isolated Lactobacillus plantarum NK151 and Bifidobacterium longum NK173, which inhibited the LPS production of Escherichia coli K1 in vitro, from the human fecal bacteria collection, and examined their effects on the Escherichia coli K1- or LPS-induced cognitive impairment in mice.
2. Materials and methods
2.1. Materials
LPS (from Escherichia coli) and 4′,6-diamidino-2-phenylindole dilactate (DAPI) were purchased from Sigma (St Louis, MO). The De Man, Rogosa, and Sharpe (MRS) medium was purchased from BD (Franklin Lakes, NJ). The general anaerobic medium (GAM) was purchased from Nissui Pharmaceutical Co. (Tokyo, Japan). The antibody for NF-κB was purchased from Cell Signaling Technology (Danvers, MA). Antibodies for CD11c, Iba1, and BDNF were purchased from Abcam (Cambridge, UK). The antibody for NeuN was purchased from Millipore (Burlington, MA). Alexa Fluor 488 and Alexa Fluor 488 594 were purchased from Invitrogen (Carlsbad, CA). Enzyme-linked immunosorbent assay (ELISA) kits for IL-1β, IL-10, and TNF-α were purchased from Ebioscience (Atlanta, GA). A Limulus Amebocyte Lysate (LAL) assay kit was purchased from Cape Cod Inc. (East Falmouth, MA). A QIAamp Fast DNA stool mini kit was purchased from Qiagen (Hilden, Germany). EasyTaq DNA polymerase and 100 bp plus II DNA ladder were purchased from TransGen Biotech (Beijing, China). TB Green® Premix Ex Taq™ II was purchased from Takara Bio (Shiga, Japan).2.2. Isolation of Escherichia coli K1 LPS production-suppressing probiotics
Escherichia coli K1, which was isolated as a gut bacterium causing depression and memory impairment from human feces,21,26 and probiotics, which were selected from human fecal bacteria collection, were cultured in GAM broth at 37 °C for 24 h. Thereafter, Escherichia coli K1 (1 × 105 CFU mL−1) and probiotics (1 × 104 or 1 × 106 CFU mL−1) were simultaneously inoculated into GAM broth (10 mL) and anaerobically cultured at 37 °C for 24 h. These cultured broths were centrifuged at 15000g and at 4 °C for 20 min. The resulting supernatant was successively filtered through 0.45 μm and 0.22 μm filters. The LPS level was measured in the supernatant using a LAL assay kit.
2.3. Culture of Lactobacillus plantarum NK151 and Bifidobacterium longum NK173
Gut bacteria including NK151 and NK173 were cultured in general media for probiotics, such as De Man, Rogosa and Sharpe (MRS) broth, and centrifuged at 5000g and 4 °C for 20 min, and washed with saline. For the in vivo experiment, the cells were collected, and freeze-dried and suspended in 1% maltose.2.4. Animals
Specific pathogen-free C57BL/6 mice (male, 19–22 g, 6 week-old) were purchased from Koatech (Pyungtaek-Shi, Korea). All mice were housed in the plastic cage with the 5 cm-raised wire floor under standard conditions (temperature, 20–22 °C; humidity, 50 ± 10% humidity; and light/dark cycle, 12 h), fed standard laboratory chow and water ad libitum. Mice were acclimated for one week before being used in the experiment. Each group consisted of seven mice. All animal experiments were approved by The Committee for the Care and Use of Laboratory Animals in Kyung Hee University (IACUC No., KHUASP(SE)-20331) and performed according to The Kyung Hee University Guidelines for Laboratory Animals Care and Usage.2.5. Preparation of mice with cognitive impairment
Mice with cognitive impairment were prepared, as previously reported.21,26 First, the mice were randomly divided into five groups (NC, EC, NK151, NK173, and NKm). Escherichia coli K1 (EC, 1 × 109 CFU per mouse per day) were orally gavaged once a day for 5 days except for the NC group. In the NC group, 1% maltose (vehicle) was orally gavaged instead of Escherichia coli K1. Test agents (NC, vehicle; EC, vehicle; NK151, 1 × 109 CFU per mouse per day of Lactobacillus plantarum NK151; NK173, 1 × 109 CFU per mouse per day of Bifidobacterium longum NK173; NKm, 1 × 109 CFU per mouse per day of NK151 and NK174 [4:1] mixture) were orally gavaged once a day for 5 days from the day after the final gavage of Escherichia coli K1. Second, mice were randomly divided into five groups (NC, LPS, NK151, NK173, and NKm). LPS (10 μg kg−1 day−1) was intraperitoneally injected once a day for 5 days except for the NC group. In the NC group, 1% maltose (vehicle) was intraperitoneally injected instead of LPS. Test agents (NC, vehicle; LPS, vehicle; NK151, 1 × 109 CFU per mouse per day of Lactobacillus plantarum NK151; NK173, 1 × 109 CFU per mouse per day of Bifidobacterium longum NK173; NKm, 1 × 109 CFU per mouse per day of NK151 and NK173 [4:1] mixture) were orally gavaged once a day for 5 days from the day after the final injection of LPS.
For the assay of cognitive function, the Y-maze and novel object recognition (NOR) tasks were performed 20 h after the final gavage of test agents. The Y-maze was performed in a three-arm horizontal maze (40 cm long and 3 cm wide with 12 cm-high walls) according to the method of Jeong et al.27 A mouse was initially placed within one arm and the sequence and number of arm entries were manually recorded for 8 min. A spontaneous alternation was defined as entries into all three arms on consecutive choices. The ratio (%) of actual to possible alternations was calculated. The NOR task was performed in the apparatus consisting of a dark-open field box (45 × 45 × 50 cm) according to the method of Jeong et al.27 For the first trial, a mouse was placed in the box containing two identical objects and the frequency of touching each object was recorded for 10 min. In the second trial conducted 24 h after the first trial, a mouse was placed in the box containing one of the old objects used in the first trial and a new object. Novel object recognition was calculated as the ratio of the number of times touching the new object to the sum of the touching frequencies.
Mice were sacrificed 3 h after the final behavioral task by CO2 inhalation. Blood, colon, brain, and stool were removed and stored at −80 °C for further experiments.
2.6. Myeloperoxidase activity assay, ELISA, and immunoblotting
Myeloperoxidase activity was assayed according to the method of Jang et al.11 The assay of corticosterone, apoE, IL-1β, IL-10, and TNF-α expression levels were performed using commercial ELISA kits.21 p-p65, p65, BDNF, and β-actin expression levels were determined according to the method of Jang et al.112.7. Immunofluorescence staining
Immunofluorescence assay was performed, as reported previously.13,21 Mice were perfused transcardially with 4% paraformaldehyde. Their hippocampus and colon tissues were post-fixed with 4% paraformaldehyde, immersed in 30% sucrose solution, frozen, and sectioned. The sections were incubated with antibodies for NF-κB (1:100), CD11c (1:200), Iba1 (1:200), BDNF (1:200), and/or NeuN (1:200) overnight and then treated with Alexa Fluor 488 (1:200)- or Alexa Fluor 594 (1:200)-conjugated secondary antibody. The cell nucleus was stained with DAPI. The immunofluorescence-stained section was observed with a confocal microscope.
2.8. Microbiota 16S rRNA gene sequencing
Bacterial genomic DNA was isolated from fresh mouse stool using a QIAamp DNA stool mini kit, as reported previously,21 and amplified using barcoded primers, which targeted the bacterial 16S rRNA V4 gene region. Amplicon sequencing was performed using Illumina iSeq 100 (San Diego, CA). Sequenced reads were stored in the NCBI's short read archive (accession number, PRJNA726149).2.9. Whole genome analysis
The sequencing libraries were prepared according to the manufacturer's instructions of 20 kb Template Preparation using the BluePippin™ Size-Selection System using the PacBio DNA Template Prep Kit 1.0. NK151 genome sequence (5 contigs) was obtained by using the PacBio RSII platform and the complete NK173 genome sequence (1 contig) was obtained by using the PacBio Sequel platform.2.10. Statistical analysis
Data values are indicated as the means ± standard deviation (SD). The significance was analyzed by using one-way analysis of variance with Tukey's multiple comparison test (p < 0.05) and the correlation between the gut microbiota and cognitive function or the LPS level in the blood and feces was determined by Pearson's correlation coefficient, using Graph-Pad Prism 9 (GraphPad Software Inc., San Diego, CA).3. Results
3.1. NK151 and NK173 inhibited Escherichia coli K1 LPS production in vitro
First, to select gut bacterial LPS production-inhibitory probiotics, we incubated Escherichia coli K1 with gut bacterial strains isolated from human feces. Of the tested gut bacteria, NK151 and NK173 suppressed the secretion of LPS from Escherichia coli K1 most strongly (Fig. 1). NK151 and NK173 were identified as Lactobacillus (Lactiplantibacillus) plantarum and Bifidobacterium longum, respectively, on the basis of the results of gram staining, API 50 CHL kit assay, 16S rDNA, and whole genome sequences. The genome of NK151 was 3288981 bp with a GC content of 44.5% (Fig. 2 and ESI Table S1†). The total number of CDS was 3930. The number of tRNA genes was 70 and of rRNA genes was 16. The genome sequence of NK151 showed phylogenetic similarity to Lactobacillus (Lactiplantibacillus) plantarum NCTC13644 (99.0%), Lactobacillus (Lactiplantibacillus) argentoratensis DSM163565 (95.5%), and Lactobacillus (Lactiplantibacillus) paraplantarum DSM10667 (86.1%), using OrthoANI. The genome of NK173 was 2507434 bp with a GC content of 59.9% (Fig. 2). The total number of CDS was 2077. The number of tRNA and rRNA genes was 59 and 12, respectively. The genome sequence of NK151 showed phylogenetic similarity to those of Bifidobacterium longum JCM1217 (98.6%), Bifidobacterium longum DSM20211 (96.3%), and Bifidobacterium longum ATCC15697 (95.0%). NK151 and NK173 did not show hemolytic activity in the blood agar plate.
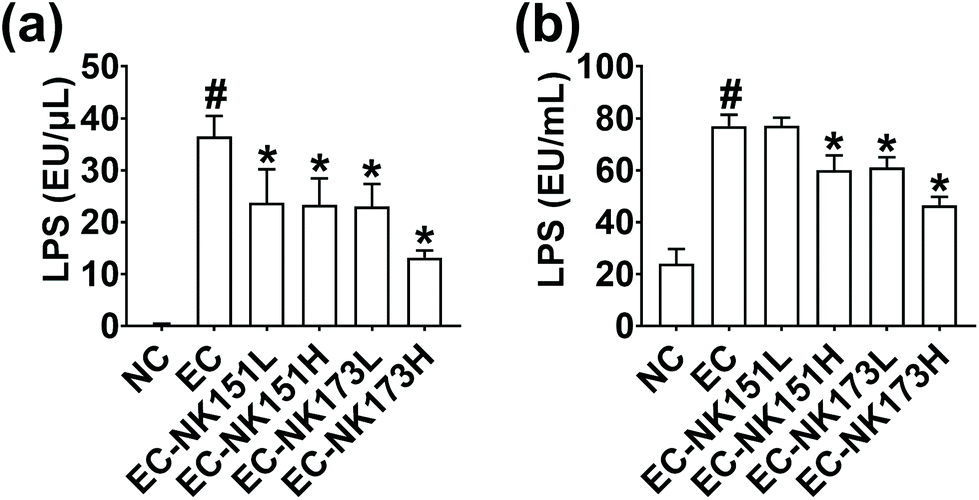 | ||
| Fig. 1 Effects of NK151 and NK173 on the LPS production in Escherichia coli K1. (a) Effects on the LPS production in the intact bacteria (precipitate). (b) Effects on the LPS production in the supernatant. Escherichia coli K1 (EC, 1 × 105 CFU) in the absence or presence of NK151 or NK173 (l, 1 × 104 CFU; h, 1 × 106 CFU) was anaerobically cultured at 37 °C for 24 h in general anaerobic medium. The LPS level was assayed in the supernatant of cultured media using an ELISA kit. Data represent the mean ± SD (n = 5 per group). #p < 0.05 vs. NC; * p < 0.05 vs. EC. | ||
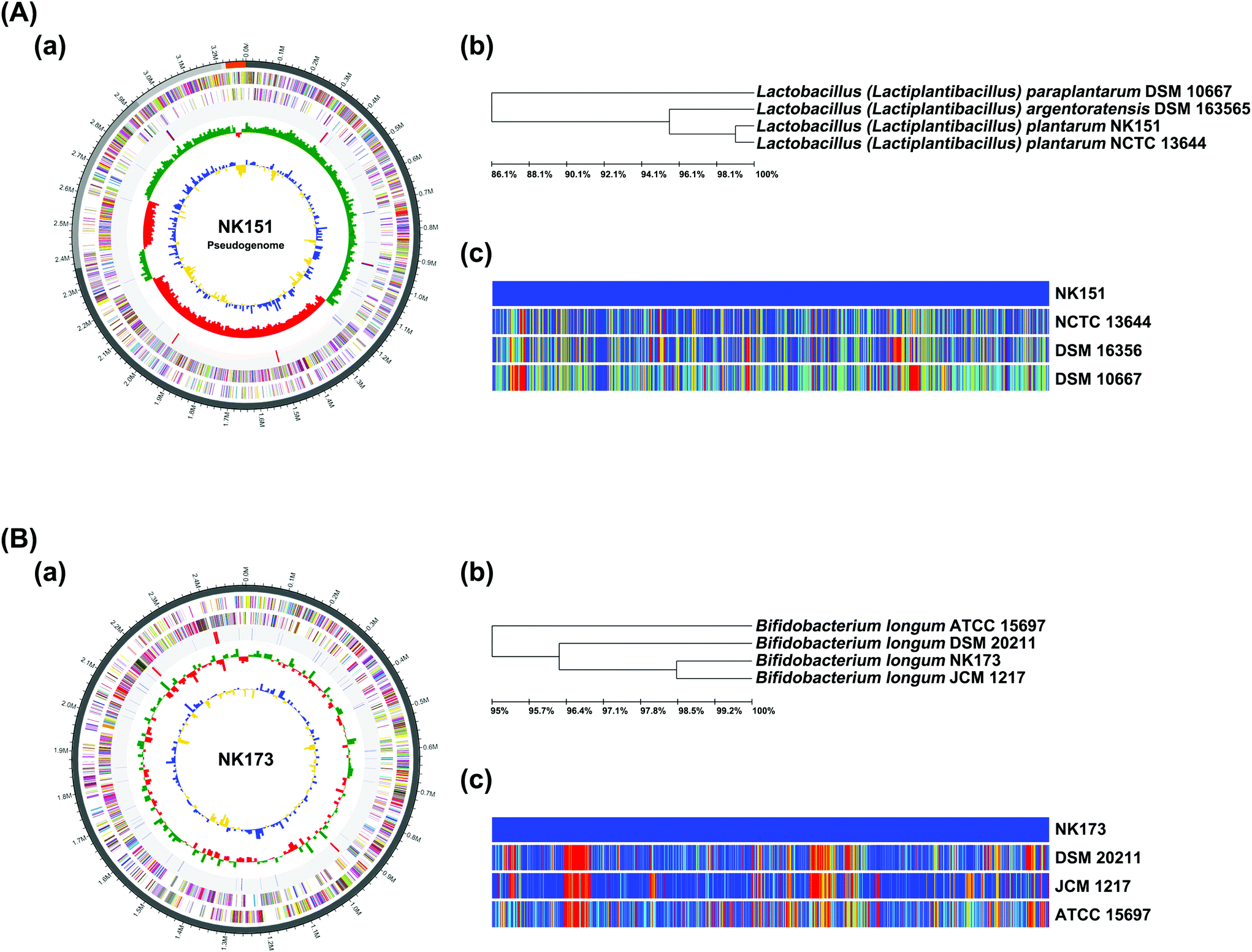 | ||
| Fig. 2 Taxonomic classification by genome-wide comparative analysis of NK151 (A) and NK173 (B). (a) The pseudochromosome drawn from 5 contigs. The outermost circle indicates contig. The inner circle is the color coded for the CDS information analyzed in the forward strand, and the inner circle is the CDS information analyzed in the reverse strand. The fourth circle from the outside represents tRNA (blue) and rRNA (red). The inner circle indicates GC skew metric information (green, higher than the average; red, lower than the average). The innermost circle is GC ratio metric information (blue, higher values than average; yellow, lower values) GC skew and GC ratio metrics are displayed at 10 kb intervals. (b) Neighbor-joining tree based on the OrthoANI distance matrix (analyzed using UPGMA dendrogram, Newick format). (c) The pairwise ortholog matrix table (generated and colored according to the similarity between matching sequences). | ||
3.2. NK151 and NK173 alleviated cognitive function, colitis, and gut dysbiosis in mice with Escherichia coli K1-induced cognitive impairment
In the preliminary study, we examined the effects of NK151 at doses of 1 × 108 and 1 × 109 CFU per mouse per day against cognitive impairment in mice orally gavaged with Escherichia coli K1. NK151 at a dose of 1 × 109 CFU per mouse per day significantly alleviated cognitive impairment in mice with Escherichia coli K1-induced cognitive impairment than at a dose of 1 × 108 CFU per mouse, while it did not affect the cognitive function in control mice (data not shown). Therefore, we examined the effects of NK151 and NK173 at a dose of 1 × 109 CFU per mouse per day on Escherichia coli K1-induced cognitive impairment in mice (Fig. 3). Oral gavage of Escherichia coli K1 significantly reduced the spontaneous alternation in the Y-maze task and exploration in the NOR task to 73.2% and 53.1% of the control group, respectively. However, oral administration of NK151, NK173, or NKm significantly alleviated Escherichia coli K1-suppressed spontaneous alternation to 93.4%, 94.1%, and 97.1% of the control group, respectively, and exploration to 77.4%, 102.4%, and 109.7%, respectively (Fig. 3b and c). Furthermore, their treatments reduced Escherichia coli K1-induced IL-1β and TNF-α expression and NF-κB activation in the hippocampus, while BDNF and IL-10 expression and BDNF to proBDNF expression ratio increased (Fig. 3d–g). They also reduced Escherichia coli K1-induced NF-κB+/Iba1+ and LPS+/Iba1+ cell populations, while the Escherichia coli K1-suppressed BDNF+/NeuN+ cell population increased (Fig. 3h). Furthermore, they significantly decreased LPS and apoE levels in the blood (Fig. 3i and j). Oral gavage of Escherichia coli K1 also caused colitis (Fig. 4). However, oral administration of NK151 and/or NK173 significantly alleviated the Escherichia coli K1-induced colon shortening, macroscopic score, myeloperoxidase activity, IL-1β and TNF-α expression, NF-κB activation, and NF-κB+/Iba1+ cell population, while IL-10 expression was increased. Furthermore, they decreased the Escherichia coli K1-induced LPS level in the feces. Of these, NKm alleviated Escherichia coli K1-induced neuroinflammation and colitis most potently. | ||
| Fig. 3 NK151 and NK173 alleviated Escherichia coli K1-induced cognitive impairment in mice. (a) Experimental procedure. (b) Effects on the spontaneous alternation in the Y-maze task. (c) Effects on the exploration (preference index) in the novel object recognition task. Effects on the BDNF and pro-BDNF expression, NF-KB activation, and their intensities (d), IL-1β (e), TNF-α (f), and IL-10 expression (g) and BDNF+/NeuN+, LPS+/Iba1+, and NF-κB+/Iba1+ cell populations (h) in the hippocampus. Effects on the LPS (i) and APOE levels (j) in the blood. Cytokines were assayed using ELISA kits. NK151, NK173, their mixture (NKm), or Escherichia coli K1 (EC, 1 × 109 CFU per mice per day) was orally gavaged. The control group (NC) was treated with the vehicle instead of test agents. Data values were described as mean ± SD (n = 7). #p < 0.05 vs. NC. *p < 0.05 vs. EC. | ||
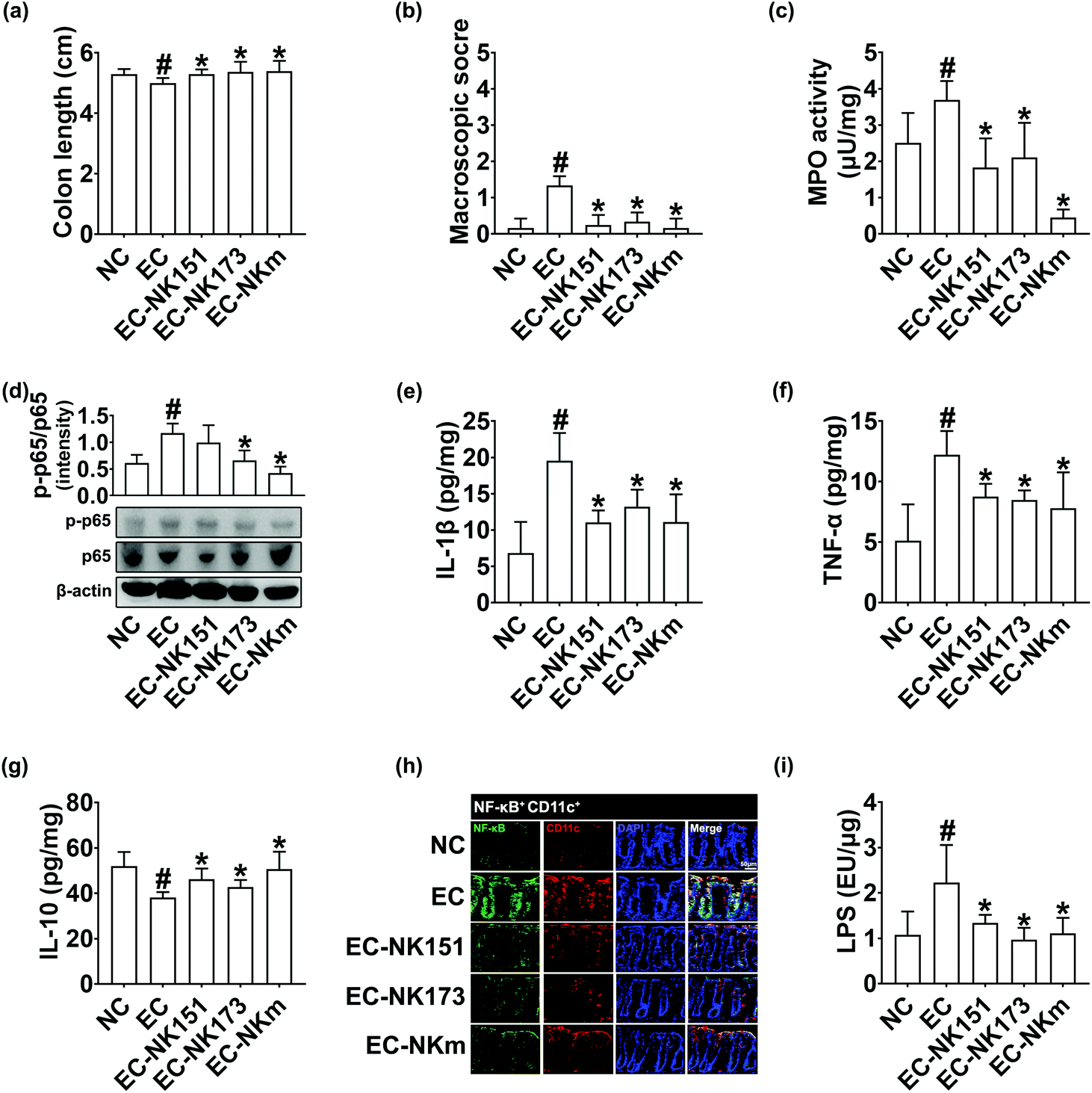 | ||
| Fig. 4 NK151 and NK173 alleviated colitis in mice with Escherichia coli K1-induced cognitive impairment. (a) Effects on the colon length. (b) Effects on the macroscopic score. (c) Effects on the myeloperoxidase activity in the colon. Effects on the NF-KB activation (d), IL-1β (e), TNF-α (f), and IL-10 expression (g) and NF-κB+/CD11c+ cell population (h) in the colon. (i) Effects on the LPS in the feces. Cytokines were assayed using ELISA kits. NK151, NK173, their mixture (NKm), or Escherichia coli K1 (EC, 1 × 109 CFU per mice per day) was orally gavaged. The control group (NC) was treated with the vehicle instead of test agents. Data values were described as mean ± SD (n = 7). #p < 0.05 vs. NC. *p < 0.05 vs. EC. | ||
Next, we investigated the effect of NKm on the fecal microbiota composition in mice with Escherichia coli K1-induced cognitive impairment. Oral gavage of Escherichia coli K1 shifted the α-(OTUs) and β-diversities (PCoA) (Fig. 5A and ESI Tables S2–S5†). However, treatment with NK151 and/or NK173 partially restored the α- and β-diversities of the gut microbiota. At the phylum level, oral gavage of Escherichia coli K1 decreased Firmicutes and Verrucomicrobia populations, while the Bacteroidetes, Proteobacteria, and Cyanobacteria populations increased. Treatment with Escherichia coli K1 also increased Rikenellaceae and AC160630_f populations at the family level; Alistipes, Helicobacter, PAC001068_g and PAC000186_g populations at the genus level; and EF097112_s, and PAC001064_s populations at the species level, while it decreased the Lactobacillaceae, Lachnospiraceae, Akkermansiaceae, Bifidobacteriaceae, and Dehalobacterium_f populations at the family level; Lactobacillus, LLKB_g, and PAC001092_g populations at the genus level; and the Lactobacillus reuteri group, PAC001093_s, and AB626939_s populations at the species level. However, treatment by oral administration of NK151 and/or NK173 suppressed Escherichia coli K1-induced Proteobacteria at the phylum level; Rikenellaceae, Christensenellaceae, and AC160630_f populations at the family level, Alistipes and Helicobacter populations in the genus level; and PAC001071_s population at the species level and increased Escherichia coli K1-suppressed Verrucomicrobia and Firmicutes populations at the phylum level; Lactobacillaceae and Bifidobacteriaceae populations at the family level, Lactobacillus, LLKB_g, and PAC001092_g populations at the genus level; and the Lactobacillus reuteri group population at the species level.
 | ||
| Fig. 5 Effects of NK151 and NK173 on the fecal microbiota composition in mice with Escherichia coli K1-induced cognitive impairment. (A) Effects on fecal microbiota composition: the (a) OTU richness, (b) principal coordinate analysis (PCoA) plot based on Bray–Curtis analysis, and (c) phylum level. (B) The relationship between the spontaneous alternation in the Y-maze task and the PC1 of the gut bacteria PCoA plot. (C) The relationship between the spontaneous alternation in the Y-maze task and gut bacteria. [(a) Rikenellaceae, (b) Lactobacillaceae, (c) Christensenellaceae, and (d) Odoribacteraceae]. (D) The relationship between the blood LPS level and gut bacteria [(a) Rikenellaceae, (b) Lactobacillaceae, (c) Christensenellaceae, and (d) Odoribacteraceae]. (E) The relationship between the blood LPS level and gut bacteria. (F) The relationship between the fecal LPS level and gut bacteria. NK151, NK173, their mixture (NKm), or Escherichia coli K1 (EC, 1 × 109 CFU per mice per day) was orally gavaged. The control group (NC) was treated with the vehicle instead of test agents. Data values were described as mean ± SD (n = 7). #p < 0.05 vs. NC. *p < 0.05 vs. EC. | ||
Next, we analyzed the correlation between gut microbiota populations and cognitive function or LPS level in the blood and feces (Fig. 4B–F and ESI Fig. S1, S2†). The gut microbiota composition (the PC1 of the PCoA plot) was positively correlated with cognitive function in the Y-maze task (R2 = 0.1723, p = 0.013). At the family level, Lactobacillaceae (R2 = 0.0256, p = 0.359 in the Y-maze task; R2 = 0.1459, p = 0.024 in NOR task) and PAC001057_f (Mollicutes; R2 = 0.0811, p = 0.097 in Y-maze task; R2 = 0.0209, p = 0.407 in the NOR task) populations were positively correlated with cognitive function, while Helicobacteraceae (R2 = 0.0479, p = 0.207 in the Y-maze task; R2 = 0.2585, p = 0.002 in the NOR task), Rikenellaceae (R2 = 0.2749, p = 0.001 in Y-maze task; R2 = 0.2372, p = 0.003 in the NOR task), Ruminococcaceae (R2 = 0.0160, p = 0.469 in Y-maze task; R2 = 0.1149, p = 0.047 in NOR task), Christensenellaceae (R2 = 0.0077, p = 0.618 in Y-maze task; R2 = 0.1188, p = 0.043 in NOR task), and Odoribacteraceae (R2 = 0.0237, p = 0.379 in the Y-maze task; R = 0.1640, p = 0.016 in the NOR task) populations were negatively correlated with it. At the genus level, Alistipes, Oscillibacter, LT706945_g, and Rikenella populations were negatively correlated with cognitive function, while the Lactobacillus, PAC001118_g, and Roseburia populations were positively correlated with it (Fig. 4C and D). The blood LPS levels were positively correlated with the Christensenellaceae (R2 = 0.1837, p = 0.010), Odoribacteraceae (R2 = 0.3532, p ≤ 0.001), and Rikenellaceae (R2 = 0.2572, p = 0.002) populations, while it was negatively correlated with the Lactobacillaceae population (R2 = 0.0250, p = 0.365) (Fig. 4E). The fecal LPS levels were positively correlated with the Christensenellaceae (R = 0.1181, p = 0.043), Odoribacteraceae (R2 = 0.1545, p = 0.020), and Rikenellaceae (R2 = 0.0359, p = 0.275) populations, while it was negatively correlated with the Lactobacillaceae (R2 = 0.0816, p = 0.096) population (Fig. 4F).
3.3. NK151 and NK173 alleviated LPS-induced memory impairment in mice
The gut microbiota communicates with the immune and neuronal systems through the stimulation of their byproducts such as neurotransmitters and endotoxins.8–10 Excessive exposure to the bacterial LPS causes cognitive impairment.10,11 Therefore, we examined the effects of NK151 and/or NK173 at a dose of 1 × 109 CFU per mouse on LPS-induced cognitive impairment in mice (Fig. 6). Intraperitoneal injection of LPS significantly reduced the spontaneous alternation in the Y-maze task and exploration in the NOR task to 74.8% and 61.3% of the control group, respectively. However, oral administration of NK151, NK173, or NKm significantly alleviated the LPS-suppressed spontaneous alternation to 94.2%, 92.1% and 97.2% of the control group, respectively, and exploration to 80.6%, 86.3%, and 89.5%, respectively (Fig. 6b and c). Oral administration of NK151 and/or NK173 reduced LPS-induced NF-κB activation and IL-1β and TNF-α expression in the hippocampus, while BDNF and IL-10 expression and BDNF to proBDNF expression ratio increased (Fig. 6d–g). They also reduced LPS-induced NF-κB+/Iba1+ and LPS+/Iba1+ cell populations, while the K1-suppressed BDNF+/NeuN+ cell population increased (Fig. 6h). Furthermore, they significantly decreased LPS and apoE levels in the blood (Fig. 6i and j). | ||
| Fig. 6 NK151 and NK173 alleviated LPS-induced cognitive impairment in mice. (a) Experimental procedure. (b) Effects on the spontaneous alternation in the Y-maze task. (c) Effects on the exploration (preference index) in the novel object recognition task. Effects on the BDNF, pro-BDNF, NF-κB activation, and their intensities (d), IL-1β (e), TNF-α (f), and IL-10 expression (g) and BDNF+/NeuN+, LPS+/Iba1+, and NF-κB+/Iba1+ cell populations (h) in the hippocampus. Effects on the LPS (i) and APOE levels (j) in the blood. Cytokines were assayed using ELISA kits. NK151, NK173, and their mixture (NKm) (1 × 109 CFU per mice per day) were orally gavaged 24 h after the final peritoneal injection of LPS (10 ng kg−1 day−1). The control group (NC) was treated with the vehicle instead of test agents. Data values were described as mean ± SD (n = 7). #p < 0.05 vs. NC. *p < 0.05 vs. LPS. | ||
Intraperitoneal injection of LPS also caused colitis: it caused colon shortening, induced myeloperoxidase activity, NF-κB activation, and TNF-α and IL-1β expression, and decreased IL-10 expression in the colon (Fig. 7). However, oral administration of NK151 and/or NK173 significantly suppressed LPS-induced colon shortening, myeloperoxidase activity, NF-κB activation, NF-κB+/CD11c+ cell population, and TNF-α and IL-1β expression, while LPS-suppressed IL-10 expression increased.
 | ||
| Fig. 7 NK151 and NK173 alleviated colitis in mice with LPS-induced cognitive impairment. (a) Effects on the colon length. (b) Effects on the macroscopic score. (c) Effects on the myeloperoxidase activity in the colon. Effects on the NF-κB activation (d), IL-1β (e), TNF-α (f), and IL-10 expression (g) and NF-κB+/CD11c+ cell population (h) in the colon. (i) Effects on the LPS in the feces. Cytokines were assayed using ELISA kits. NK151, NK151, NK173, and their mixture (NKm) (1 × 109 CFU per mice per day) were orally gavaged 24 h after the final peritoneal injection of LPS (10 ng kg−1 day−1). The control group (NC) was treated with the vehicle instead of test agents. Data values were described as mean ± SD (n = 7). #p < 0.05 vs. NC. *p < 0.05 vs. LPS. | ||
3.4. NK151 and NK173 modulated the gut microbiota composition in mice with LPS-induced cognitive impairment
We examined whether the attenuation of LPS-induced cognitive impairment by oral administration of NK151 and/or NK173 was associated with the gut microbiota composition. The intraperitoneal injection of LPS shifted the β-diversity (principal coordinate analysis [PCoA]) while the α-diversity was not affected (Fig. 8A and ESI Tables S6–S9†). However, treatment with NK151, NK173, and NKm partially shifted the gut microbiota β-diversity of LPS-treated mice to those of normal control mice, while the α-diversity (OTUs) weakly decreased. Furthermore, LPS treatment increased Firmicutes and Deferribacteres populations, while the Bacteroidetes population decreased. However, oral gavage of NK151, NK173, or NKm reduced Firmicutes, Deferribacteres, and Proteobacteria populations and increased Bacteroidetes population at the phylum level. Furthermore, they increased Muribaculaceae, Bacteroidaceae and Lactobacillaceae, Rikenellaceae, and Odoribacteraceae populations at the family level; PAC000198_g, Bacteroides, Lactobacillus, and PAC002400_g populations at the genus level; and the Lactobacillus murinus group and Bacteroides acidifaciens group populations at the species level. However, they decreased Lachnospiraceae, Prevotellaceae, and Helicobacteraceae populations at the family level, Prevotellaceae_uc, Helicobacter, LLKB_g, and PAC001092_g populations at the genus level, and Helicobacter rodentium group and PAC001072_s populations at the species level. | ||
| Fig. 8 Effects of NK151 and NK173 on the fecal microbiota composition in mice with LPS-induced cognitive impairment. (A) Effects on fecal microbiota composition: (a) the OTU richness, (b) principal coordinate analysis (PCoA) plot based on Bray–Curtis analysis, and (c) phylum level. (B) The relationship between the spontaneous alternation in the Y-maze task and the PC3 of the gut bacteria PCoA plot. (C) The relationship between the spontaneous alternation in the Y-maze task and gut bacteria [(a) Bacteroidaceae, (b) Odoribacteraceae, (c) Rikenellaceae, and (d) Deferribacteraceae]. (D) The relationship between the exploration in the novel object recognition task and gut bacteria [(a) Bacteroidaceae, (b) Odoribacteraceae, (c) Rikenellaceae, and (d) Deferribacteraceae]. (E) The relationship between the blood LPS level and gut bacteria Odoribacteraceae. NK151, NK173, and their mixture (NKm) (1 × 109 CFU per mice per day) were orally gavaged 24 h after the final peritoneal injection of LPS (10 ng kg−1 day−1). The control group (NC) was treated with the vehicle instead of test agents. Data values were described as mean ± SD (n = 7). #p < 0.05 vs. NC. *p < 0.05 vs. LPS. | ||
Next, we analyzed the correlation between the gut microbiota composition and LPS level in the blood or cognitive function (Fig. 8B–E and ESI Fig. S3, S4†). The gut microbiota composition (the PC3 of PCoA plot) was negatively correlated with cognitive function in the Y-maze task function (R2 = 2836, 0.001). At the family level, Bacteroidaceae (R2 = 0.1493, p = 0.024 in the Y-maze task; R2 = 0.1113, p = 0.053 in the NOR task), Odoribacteraceae (R = 0.0937, p = 0.079 in the Y-maze task; R2 = 0.0598, p = 0.164 in the NOR task), and Rikenellaceae (R = 0.0176, p = 0.455 in the Y-maze task; R2 = 0.1813, p = 0.012 in the NOR task) populations showed a positive correlation with the cognitive function, while Deferribacteraceae (R2 = 0.0447, p = 0.232 in the Y-maze task; R = 0.2267, p = 0.004 in the NOR task) population was negatively correlated with it (Fig. 7B and C). At the genus level, Alistipes, Bacteroides, Odoribacter, and KE159600_g populations were positively correlated with the cognitive function; while PAC000692_g (Lachnospiraceae), PAC001372_g, (Lachnospiraceae), and Sporobacter (Ruminococcaceae) populations were negatively correlated with it. The blood LPS levels were negatively correlated with the Odoribacteraceae (R2 = 0.1152, p = 0.049) and Odoribacter (R2 = 0.1137, p = 0.051) populations, while it was positively correlated with the PAC001118_g (Lachnospiraceae; R2 = 0.1155, p = 0.049), PAC001165_g (Lachnospiraceae; R2 = 0.1574, p = 0.020) populations (Fig. 7D).
4. Discussion
The bacterial LPS is a risk factor for the outbreak of sporadic AD.7 In the present study, oral gavage of Escherichia coli K1 or intraperitoneal injection of LPS increased the blood LPS level and hippocampal TNF-α and IL-6 expression and NF-κB+/Iba1+ and LPS+/Iba1+ cell populations and decreased the BDNF+/NeuN+ cell population in mice. They also caused cognitive impairment-like behaviors. Bacterial LPS levels are higher in AD patients than in healthy volunteers.7,28 Buttini et al. reported that a peritoneal injection of LPS activated microglia and strongly produced proinflammatory cytokines.29 Lee et al. reported that the intraperitoneal injection of LPS caused cognitive impairment and neuroinflammation in mice.24 Goel et al. reported that exposure to LPS impaired memory by modulating NF-κB-mediated BDNF expression.30 Kim et al. reported that oral gavage of Escherichia coli K1 impaired cognitive function in mice and increased the NF-κB+/Iba1+ cell population and TNF-α and IL-1β expression and suppressed the BDNF+/NeuN+ cell population and BDNF expression in the hippocampus.21 These suggest that LPS or Escherichia coli K1 can induce cognitive impairment by suppressing LPS-induced NF-κB activation-mediated BDNF expression. Wu et al. reported that chronic exposure to LPS induced AD-like phenotypes in mice.31 Lee et al. reported that blood and fecal LPS and hippocampal Aβ expression levels were higher in 5XFAD mice, which exhibited gut dysbiosis, than in healthy control mice.24 Zhan et al. reported that LPS induced NF-κB mediated inflammatory cytokines, which increase Aβ levels in the AD brains of rodents.7 These results suggest that the overexpression of gut bacterial LPS can induce neuroinflammation and Aβ formation, resulting in the occurrence of AD. We found that LPS or Escherichia coli K1 also caused colitis: they increased myeloperoxidase activity, TNF-α and IL-6 expression, and NF-κB+/Iba1+ cell population in the colon. Furthermore, they caused gut dysbiosis: they increased the Proteobacteria population and LPS level in the gut microbiota. These results support the previous suggestion that the gut microbiota bidirectionally communicates with the brain through the activation of the microbiota–gut–brain axis, linking the cognitive brain system with gut microbiota through the peripheral enteric nervous system.32,33We also found that oral administration of NK151 and/or NK173 suppressed LPS- or Escherichia coli K1-induced NF-κB+/Iba1+ and LPS+/Iba1+ cell population and TNF-α, IL-1β, and IL-6 expression in the hippocampus, while the BDNF+/NeuN+ cell population was increased. Furthermore, they significantly suppressed LPS levels in the blood. TNF-α expression is increased in the blood and cerebrospinal fluid of AD patients, compared with those in healthy volunteers.34 Lew et al. reported that Lactobacillus plantarum P8 reduced cognitive symptoms and the blood TNF-α level in stressed adults.35 Ni et al. reported that Bifidobacterium longum BL986 alleviated learning and memory ability and BDNF expression in aged mice.36 A TNF-α blocking agent reduced the risk for Alzheimer's disease (AD) in patients with IBD.37,38 These suggest that NK151 and/or NK173 can alleviate LPS- or Escherichia coli K1-induced cognitive impairment by increasing BDNF expression and BDNF to proBDNF expression ratio through the suppression of NF-κB activation. The combination of NK151 and NK173 synergistically or additively alleviated cognitive impairment. Moreover, although we did not analyse the Aβ expression levels, these probiotics may suppress Aβ expression in the brain by the suppression of LPS-inducible neuroinflammation and Aβ formation.
Oral gavage of Escherichia coli K1 increased blood LPS and apoE levels in mice. Intraperitoneal injection of LPS also increased the blood apoE levels in mice. Van Oosten et al. reported that the intravenous injection of LPS significantly increased the blood apoE level in mice.39 ApoE, which is primarily produced by the liver cells such as Kupffer cells and the brain cells such as astrocytes,14,40 binds LPS in the blood and most of the LPS is cleared by the liver. ApoE inhibits toll-like receptor (TLR)-3 and TLR-4-mediated inflammatory responses in macrophages.41–43 LPS and its producer Escherichia coli K1 cause systemic inflammation and cognitive impairment in mice.11 The concentration of apoE expression in blood is dependent on the expression of LPS-inducible proinflammatory cytokines TNF-α and IL-1β.39 These suggest that the translocation of bacterial LPS produced in the gut into the blood can induce the expression of apoE, which degrade LPS in the liver, and, if not excluded, excessive exposure to LPS may cause systemic inflammation and cognitive impairment.
Oral administration of NK151 and/or NK173 NK151 and/or NK173 suppressed the blood apoE level, which is dependent on the fecal and blood LPS levels in mice with LPS- or K1-induced cognitive impairment. They also suppressed gut bacterial LPS production in mice and reduced Escherichia coli K1 LPS production in vitro. These suggest that NK151 and/or NK173 may alleviate cognitive impairment by the suppression of systemic inflammation through the suppression of gut bacterial LPS production.
Oral gavage of Escherichia coli K1 or intraperitoneal injection of LPS caused colitis: it increased myeloperoxidase activity and proinflammatory cytokine expression in the colon. Moreover, it altered gut microbiota composition: LPS treatment shifted β-diversity, not α-diversity, while Escherichia coli K1 treatment affected α- and β-diversities. In particular, LPS treatment increased the Firmicutes population, while Escherichia coli K1 treatment suppressed it. However, Escherichia coli K1 treatment increased Proteobacteria and Bacteroidetes populations. They all increased the LPS level in the blood. Liu et al. reported that the γ-Proteobacteria population was higher in patients with AD than in healthy volunteers.44 Lee et al. reported that the Proteobacteria population and endotoxin levels were higher in 5XFAD and aged mice than young mice.23 These results suggest that cognitive impairment may be dependent on the Proteobacteria population and bacterial LPS levels, which may be a risk factor for the outbreak of cognitive impairment.
NK151 and NK173 significantly alleviated Escherichia coli K1- or LPS-induced gut microbiota alternation and cognitive impairment with colitis. They also suppressed LPS levels in the blood and feces and suppressed the Proteobacteria population in the gut microbiota. They can increase LPS-suppressed Bacteroidaceae, Odoribacteraceae, and Rikenellaceae populations and Escherichia coli K1-suppressed Verrucomicrobiota, Firmicutes, Lactobacillaceae and Bifidobacteriaceae populations. However, they decreased LPS-induced Lachnospiraceae, Prevotellaceae, Helicobacteraceae, and Deferribacteraceae populations and Escherichia coli K1-induced Proteobacteria, Rikenellaceae, Christensenellaceae, AC160630_f populations. Thus, the LPS-induced alteration of the gut microbiota composition was different from the Escherichia coli K1-induced one. However, their common features were that stressors LPS and Escherichia coli K1 increased the LPS level in the blood. In mice with Escherichia coli K1-induced cognitive impairment, gut bacteria, particularly Lactobacillaceae and PAC001057_f populations were positively correlated with the cognitive function, while Helicobacteraceae, Rikenellaceae, Christensenellaceae, and Odoribacteraceae populations were negatively correlated with it. Of these, Christensenellaceae and Odoribacteraceae were positively correlated with the blood and fecal LPS levels. However, in mice with LPS-induced cognitive impairment, gut bacteria, particularly Bacteroidaceae, Odoribacteraceae, and Rikenellaceae populations showed a positive correlation with the cognitive function, while Deferribacteraceae population was negatively correlated with it. Of these, Odoribacteraceae and Odoribacter were negatively correlated with the blood LPS levels. Overall, the gut microbiota composition was correlated with the cognitive function in mice with Escherichia coli K1-induced cognitive impairment more closely than in mice with LPS-induced cognitive impairment. These results suggest that NK151 and/or NK173 may alleviate cognitive impairment and systemic inflammation by suppressing the blood LPS and through the restoration of the gut microbiota. Moreover, the combination of NK151 and NK173 synergistically or additively alleviated cognitive impairment and systemic inflammation including neuroinflammation and colitis.
5. Conclusions
Oral gavage of NK151, NK173, and their (4:1) mixture (NKm) significantly alleviated Escherichia coli K1- or LPS-induced cognitive impairment-like behaviors and colitis. Treatment with them decreased inflammatory marker expression in the hippocampus and colon, while the BDNF+/NeuN+ cell population increased. They also reduced LPS and apolipoprotein E levels in the blood. Although the exact mechanism needs to be further elucidated, NK151 and/or NK173 can alleviate cognitive impairment with neuroinflammation by upregulating NF-κB-mediated BDNF expression and BDNF to proBDNF expression ratio through the suppression of fecal and blood bacterial LPS levels, and may thereby be beneficial for therapy of dementia.
Conflicts of interest
The authors have declared no conflict of interest.Acknowledgements
This research was funded by the Medical Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science and ICT (NRF-2017R1A5A2014768) and the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry (IPET) through High Value-added Food Technology Development Program funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA 318027-4).References
- M. A. DeTure and D. W. Dickson, The neuropathological diagnosis of Alzheimer's disease, Mol. Neurodegener., 2019, 14, 32 CrossRef PubMed.
- S. A. Gale, D. Acar and K. R. Daffner, Dementia, Am. J. Med., 2018, 131, 1161–1169 CrossRef PubMed.
- M. Dugu, J. Neugroschl, M. Sewell and D. Marin, Review of dementia, Mt. Sinai J. Med., 2003, 70, 45–53 Search PubMed.
- K. Blennow, M. J. de Leon and H. Zetterberg, Alzheimer's disease, Lancet, 2006, 368, 387–403 CrossRef CAS.
- F. Zhu, C. Li, F. Chu, X. Tian and J. Zhu, Target dysbiosis of gut microbes as a future therapeutic manipulation in Alzheimer's disease, Front. Aging Neurosci., 2020, 12, 544235 CrossRef CAS PubMed.
- K. Kowalski and A. Mulak, Brain-gut-microbiota axis in Alzheimer's disease, J. Neurogastroenterol. Motil., 2019, 25, 48–60 CrossRef PubMed.
- X. Zhan, B. Stamova and F. R. Sharp, Lipopolysaccharide Associates with Amyloid Plaques, Neurons and Oligodendrocytes in Alzheimer's Disease Brain: A Review, Front. Aging Neurosci., 2018, 10, 42 CrossRef PubMed.
- J. F. Cryan, K. J. O'Riordan, K. Sandhu, V. Peterson and T. G. Dinan, The gut microbiome in neurological disorders, Lancet Neurol., 2020, 19, 179–194 CrossRef CAS PubMed.
- J. R. Kelly, G. Clarke, J. F. Cryan and T. G. Dinan, Brain-gut-microbiota axis: challenges for translation in psychiatry, Ann. Epidemiol., 2016, 26, 366–372 CrossRef PubMed.
- H. M. Jang, H. J. Lee, S. E. Jang, M. J. Han and D. H. Kim, Evidence for interplay among antibacterial-induced gut microbiota disturbance, neuro-inflammation, and anxiety in mice, Mucosal Immunol., 2018, 11, 1386–1397 CrossRef CAS PubMed.
- H. M. Jang, K. E. Lee, H. J. Lee and D. H. Kim, Immobilization stress-induced Escherichia coli causes anxiety by inducing NF-κB activation through gut microbiota disturbance, Sci. Rep., 2018, 8, 13897 CrossRef PubMed.
- X. Zhao, F. Cao, Q. Liu, X. Li, G. Xu, G. Liu, Y. Zhang, X. Jang, S. Yi, E. Xu, K. Fan and M. Ma, Behavioral, inflammatory and neurochemical disturbances in LPS and UCMS-induced mouse models of depression, Behav. Brain Res., 2019, 364, 494–502 CrossRef CAS PubMed.
- K. E. Lee, J. K. Kim, S. K. Han, D. Y. Lee, H. J. Lee, S. V. Yim and D. H. Kim, The extracellular vesicle of gut microbial Paenalcaligenes hominis is a risk factor for vagus nerve-mediated cognitive impairment, Microbiome, 2020, 8, 107 CrossRef PubMed.
- G. C. Brown, The endotoxin hypothesis of neurodegeneration, J. Neuroinflammation, 2019, 16, 180 CrossRef PubMed.
- A. Cattaneo, N. Cattane, V. Begni, C. M. Pariante and M. A. Riva, The human BDNF gene: peripheral gene expression and protein levels as biomarkers for psychiatric disorders, Transl. Psychiatry, 2016, 6, e958 CrossRef CAS PubMed.
- A. E. Autry and L. M. Monteggia, Brain-derived neurotrophic factor and neuropsychiatric disorders, Pharmacol. Rev., 2012, 64, 238–258 CrossRef CAS PubMed.
- T. K. S. Ng, C. S. H. Ho, W. W. S. Tam, E. H. Kua and R. C. M. Ho, Decreased serum brain-derived neurotrophic factor (BDNF) levels in patients with Alzheimer's disease (AD): A systematic review and meta-analysis, Int. J. Mol. Sci., 2019, 20, 257 CrossRef PubMed.
- E. M. Borba, J. A. Duarte, G. Bristot, E. Scotton, A. L. Camozzato and M. L. F. Chavesa, Brain-derived neurotrophic factor serum levels and hippocampal volume in mild cognitive impairment and dementia due to Alzheimer disease, Dement. Geriatr. Cogn. Dis. Extra, 2016, 6, 559–567 CrossRef PubMed.
- Z. Yao, J. M. Mates, A. M. Cheplowitz, L. P. Hammer, A. Maiseyeu, G. S. Phillips, M. D. Wewers, M. V. S. Rajaram, J. M. Robinson, C. L. Anderson and L. P. Ganesan, Blood-borne lipopolysaccharide is rapidly eliminated by liver sinusoidal endothelial cells via high-density lipoprotein, J. Immunol., 2016, 197, 2390–2399 CrossRef CAS PubMed.
- M. Van Oosten, P. C. Rensen, E. S. Van Amersfoort, M. Van Eck, A. M. Van Dam, J. J. Breve, T. Vogel, A. Panet, T. J. Van Berkel and J. Kuiper, Apolipoprotein E protects against bacterial lipopolysaccharide-induced lethality, J. Biol. Chem., 2001, 276, 8820–8824 CrossRef CAS PubMed.
- J. K. Kim, K. E. Lee, S. A. Lee, H. M. Jang and D. H. Kim, Interplay between human gut bacteria Escherichia coli and Lactobacillus mucosae in the occurrence of neuropsychiatric disorders in mice, Front. Immunol., 2020, 11, 273 CrossRef CAS PubMed.
- J. J. Jeong, J. Y. Woo, K. A. Kim, M. J. Han and D. H. Kim, Lactobacillus pentosus var. plantarum C29 ameliorates age-dependent memory impairment in Fischer 344 rats, Lett. Appl. Microbiol., 2015, 60, 307–314 CrossRef CAS PubMed.
- H. J. Lee, Y. H. Hwang and D. H. Kim, Lactobacillus plantarum C29-fermented soybean (DW2009) alleviates memory impairment in 5XFAD transgenic mice by regulating microglia activation and gut microbiota composition, Mol. Nutr. Food Res., 2018, 62, e1800359 CrossRef PubMed.
- H. J. Lee, K. E. Lee, J. K. Kim and D. H. Kim, Suppression of gut dysbiosis by Bifidobacterium longum alleviates cognitive decline in 5XFAD transgenic and aged mice, Sci. Rep., 2019, 9, 11814 CrossRef PubMed.
- J. J. Jeong, K. A. Kim, Y. J. Hwang, M. J. Han and D. H. Kim, Anti-inflammaging effects of Lactobacillus brevis OW38 in aged mice, Benefic. Microbes, 2016, 7, 707–718 CrossRef CAS PubMed.
- S. W. Yun, J. K. Kim, K. E. Lee, Y. J. Oh, H. J. Choi, M. J. Han and D. H. Kim, A probiotic Lactobacillus gasseri alleviates Escherichia coli-induced cognitive impairment and depression in mice by regulating IL-1beta expression and gut microbiota, Nutrients, 2020, 12, 3441 CrossRef CAS PubMed.
- M. Y. Jeong, H. M. Jang and D. H. Kim, High-fat diet causes psychiatric disorders in mice by increasing Proteobacteria population, Neurosci. Lett., 2019, 698, 51–57 CrossRef CAS PubMed.
- J. M. Hill and W. J. Lukiw, Microbial-generated amyloids and Alzheimer's disease (AD), Front. Aging Neurosci., 2015, 7, 9 Search PubMed.
- M. Buttini, S. Limonta and H. W. Boddeke, Peripheral administration of lipopolysaccharide induces activation of microglial cells in rat brain, Neurochem. Int., 1996, 29, 25–35 CrossRef CAS PubMed.
- R. Goel, S. A. Bhat, K. Hanif, C. Nath and R. Shukla, Angiotensin II receptor blockers attenuate lipopolysaccharide-induced memory impairment by modulation of NF-κB-mediated BDNF/CREB expression and apoptosis in spontaneously hypertensive rats, Mol. Neurobiol., 2018, 55, 1725–1739 CrossRef CAS PubMed.
- Z. Wu, J. Ni, Y. Liu, J. L. Teeling, F. Takayama, A. Collcutt, P. Ibbett and H. Nakanishi, Cathepsin B plays a critical role in inducing Alzheimer's disease-like phenotypes following chronic systemic exposure to lipopolysaccharide from Porphyromonas gingivalis in mice, Brain, Behav., Immun., 2017, 65, 350–361 CrossRef CAS PubMed.
- B. B. Yoo and S. K. Mazmanian, The enteric network: Interactions between the immune and nervous systems of the gut, Immunity, 2017, 46, 910–926 CrossRef CAS PubMed.
- M. Carabotti, A. Scirocco, M. A. Maselli and C. Severia, The gut-brain axis: interactions between enteric microbiota, central and enteric nervous systems, Ann. Gastroenterol., 2015, 28, 203–209 Search PubMed.
- S. L. Montgomery and W. J. Bowers, Tumor necrosis factor-alpha and the roles it plays in homeostatic and degenerative processes within the central nervous system, J. Neuroimmune Pharmacol., 2012, 7, 42–59 CrossRef PubMed.
- L. C. Lew, Y. Y. Hor, N. A. A. Yusoff, S. B. Choi, M. S. B. Yusoff, N. S. Roslan, A. Ahmad, J. A. M. Mohammad, M. F. I. L. Abdullah, N. Zakaria, N. Wahid, Z. Sun, L. Y. Kwok, H. Zhang and M. T. Liong, Probiotic Lactobacillus plantarum P8 alleviated stress and anxiety while enhancing memory and cognition in stressed adults: A randomised, double-blind, placebo-controlled study, Clin. Nutr., 2019, 38, 2053–2064 CrossRef CAS PubMed.
- Y. Ni, X. Yang, L. Zheng, Z. Wang, L. Wu, J. Jiang, T. Yang, L. Ma and Z. Fu, Lactobacillus and Bifidobacterium improves physiological function and cognitive ability in aged mice by the regulation of gut microbiota, Mol. Nutr. Food Res., 2019, 63, e1900603 CrossRef PubMed.
- M. Zhou, R. Xu, D. C. Kaelber and M. E. Gurney, Tumor necrosis factor (TNF) blocking agents are associated with lower risk for Alzheimer's disease in patients with rheumatoid arthritis and psoriasis, PLoS One, 2020, 15, e0229819 CrossRef CAS PubMed.
- N. Torres-Acosta, J. H. O'Keefe, E. L. O'Keefe, R. Isaacson and G. Small, Therapeutic potential of TNF-alpha inhibition for Alzheimer's disease prevention, J. Alzheimer's Dis., 2020, 78, 619–626 CAS.
- M. Van Oosten, P. C. N. Rensen, E. S. Van Amersfoort, M. Van Eck, A. M. Van Dam, J. J. Breve, T. Vogel, A. Panet, T. Van Berkel and J. Kuiper, Apolipoprotein E protects against bacterial lipopolysaccharide-induced lethality, J. Biol. Chem., 2001, 276, 8820–8824 CrossRef CAS PubMed.
- K. Patra, A. Giannisis, A. K. Edlund, S. B. Sando, C. Lauridsen, G. Berge, G. R. Grøntvedt, G. Bråthen, L. R. White and H. M. Nielsen, Plasma apolipoprotein E monomer and dimer profile and relevance to Alzheimer's disease, J. Alzheimer's Dis., 2019, 71, 1217–1231 CAS.
- Y. Zhu, A. Kodvawala and D. Y. Hui, Apolipoprotein E inhibits toll-like receptor (TLR)-3- and TLR-4-mediated macrophage activation through distinct mechanisms, Biochem. J., 2010, 428, 47–54 CrossRef CAS PubMed.
- D. Baitsch, H. H. Bock, T. Engel, R. Telgmann, C. Müller-Tidow, G. Varga, M. Bot, J. Herz, H. Robenek, A. von Eckardstein and J. R. Nofer, Apolipoprotein E induces antiinflammatory phenotype in macrophages, Arterioscler Thromb Vasc Biol., 2011, 31, 1160–1168 CrossRef CAS PubMed.
- H. Zhang, L. M. Wu and J. Wu, Cross-talk between apolipoprotein E and cytokines, Mediators Inflammation, 2011, 2011, 949072 Search PubMed.
- P. Liu, L. Wu, G. Peng, Y. Han, R. Tang, J. Ge, L. Zhang, L. Jia, S. Yue, K. Zhou, L. Li, B. Luo and B. Wang, Altered microbiomes distinguish Alzheimer's disease from amnestic mild cognitive impairment and health in a Chinese cohort, Brain, Behav., Immun., 2019, 80, 633–643 CrossRef PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Table S1. Effects of NK151 and NK172 on the gut microbiota composition at the phylum level in mice with Escherichia coli K1-induced cognitive impairment. Table S2. Effects of NK151 and NK172 on the gut microbiota composition at the family level in mice with Escherichia coli K1-induced cognitive impairment. Table S3. Effects of NK151 and NK172 on the gut microbiota composition at the genus level in mice with Escherichia coli K1-induced cognitive impairment. Table S4. Effects of NK151 and NK172 on the gut microbiota composition at the species level in mice with Escherichia coli K1-induced cognitive impairment. Table S5. Effects of NK151 and NK172 on the gut microbiota composition at the phylum level in mice with LPS-induced cognitive impairment. Table S6. Effects of NK151 and NK172 on the gut microbiota composition at the family level in mice with LPS-induced cognitive impairment. Table S7. Effects of NK151 and NK172 on the gut microbiota composition at the genus level in mice with LPS-induced cognitive impairment. Table S8. Effects of NK151 and NK172 on the gut microbiota composition at the species level in mice with LPS-induced cognitive impairment. Fig. S1. Effects of NK151 and NK173 on the fecal microbiota composition in mice with Escherichia coli K1-induced cognitive impairment. Fig. S2. Effects of NK151 and NK173 on the fecal microbiota composition in mice with LPS-induced cognitive impairment. See DOI: 10.1039/d1fo02167b |
| This journal is © The Royal Society of Chemistry 2021 |