
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Transient in situ measurement of kombucha biofilm growth and mechanical properties
Pascal
Bertsch
 a,
Danai
Etter
ab and
Peter
Fischer
*a
a,
Danai
Etter
ab and
Peter
Fischer
*a
aInstitute of Food Nutrition and Health, ETH Zurich, 8092 Zurich, Switzerland. E-mail: peter.fischer@hest.ethz.ch
bInstitute for Food Safety and Hygiene, Vetsuisse Faculty, University of Zürich, 8057 Zurich, Switzerland
First published on 29th March 2021
Abstract
Kombucha is a traditional beverage obtained by the fermentation of sugared tea by a symbiotic culture of bacteria and yeast which has recently re-emerged as a popular lifestyle product with potential health benefits. The characteristic feature of kombucha is the formation of a cellulosic biofilm due to the excretion of bacterial cellulose with high purity and crystallinity. Despite the growing industrial and technological interest in kombucha, current characterization techniques rely on the periodic sampling of tea broth or biofilm and ex situ analysis of its biochemical or microbial composition. Here, we use interfacial shear rheology (ISR) for the transient in situ determination of kombucha biofilm growth directly at the interface. ISR revealed that kombucha biofilm formation is a two step process with clearly distinguishable growth phases. The first phase can be attributed to the initial adsorption of bacteria at the air–water interface and shows great variability, probably due to varying bacteria content and composition. The second phase is initiated by bacterial cellulose excretion and shows astonishing reproducibility regarding onset and final mechanical properties. Hence, ISR qualifies as a new in situ characterization technique for kombucha biofilm growth and bacterial cellulose production.
1. Introduction
Kombucha is a mildly alcoholic and effervescent beverage obtained by the fermentation of sugared tea by a symbiotic culture of bacteria and yeast (SCOBY). The yeasts metabolize sucrose to ethanol and CO2. Acetic acid bacteria further metabolize ethanol to acetic acid and excrete bacterial cellulose, forming the characteristic floating cellulose biofilm of kombucha shown in Fig. 1. Originally a preservation method, kombucha has re-emerged since the 2000s as a popular lifestyle product with potential health benefits which is produced industrially and homebrewed globally.1,2 | ||
| Fig. 1 Kombucha after four weeks of fermentation with characteristic cellulosic biofilm. | ||
The yeast and bacteria composition of kombucha can differ significantly depending on the starter culture and growth conditions. The most commonly reported yeasts include Saccharomyces, Schizosaccharomyces, Brettanomyces, and Zygosaccharomyces.3–6 The yeasts cleave sucrose into glucose and fructose, and subsequently metabolize it to ethanol and CO2. The yeast ecology changes during the fermentation from osmotolerant species in early stages to acid tolerant species in later stages.4 The majority of bacteria in kombucha are acetic acid bacteria, i.e. Acetobacter, Gluconobacter, and Komagataeibacter, which metabolize glucose or ethanol to acetic acid. The most prominent bacterium is Acetobacter xylinum, which was reclassified as Komagataeibacter xylinus in 2012.1,5–7K. xylinus is best known for its ability to produce bacterial cellulose. The bacteria excrete 2–4 nm thick cellulose nanofibrils from their membrane which assemble into larger macrofibrils, and ultimately form the 3D cellulosic biofilm typical for kombucha.8–11 Bacterial cellulose is valued in material sciences due to its high purity and degree of crystallinity, and kombucha biofilms are thus increasingly considered as renewable raw material for bioadsorbents, packaging materials, or biomedical applications.12
The biochemical composition of kombucha continuously changes during fermentation. The sucrose concentration decreases due to ethanol fermentation, which is in turn the educt for acetic acid fermentation.1,5,13 With ongoing fermentation time, the concentration of amino acids, vitamins, minerals, and organic acids with potential health benefits increases.14–17 Due to tea polyphenols and vitamin C production kombucha exhibits pronounced antioxidant activity.17,18 Furthermore, probiotic effects have been reported.19 There is a nearly endless list of proclaimed health benefits associated with kombucha consumption, however, there is very limited empirical evidence for health benefits in humans.20 Hence, although kombucha certainly contains valuable nutrients and potential bioactives, most of the alleged health benefits remain to be confirmed in clinical studies.21,22
Current kombucha characterization techniques mostly rely on the periodic sampling of the broth or biofilm and analysis of their biochemical (sucrose, ethanol, organic acids, pH) or microbial composition. Here, we demonstrate that interfacial shear rheology (ISR) allows for the transient in situ measurement of kombucha biofilm growth, revealing new aspects of kombucha biofilm dynamics and mechanical properties.
2. Experimental
2.1. Kombucha preparation
Black tea was prepared by infusing 6 g L−1 Lipton Yellow Label in boiling deionized water for 7 minutes and dissolving 100 g L−1 sucrose (BioXtra, ≥99.5%, Sigma). As a starting culture 10 vol% kombucha broth that has been equally prepared and fermented for 10 days was added to the sugared tea. Hence, the initial sucrose concentration was in the range of 90 g L−1, which has been reported to induce maximum cellulose production.10 The initial SCOBY was obtained from a home-brewing setup.2.2. Interfacial shear rheology (ISR)
An Anton Paar MCR 502 was equipped with a bicone custom-made from polyamide PA12 by selective laser sintering with a radius of 34.15 mm and cone angle of 13.2°. In contrast to common biconical disk geometries, it had a considerable edge thickness of 3 mm to account for the 3D structure of kombucha biofilms. 150 mL of inoculated black tea were filled in a glass cup with 40 mm radius. The surface area/depth ratio was 94 mm, which has been reported to affect cellulose production as acetic acid bacteria are aerobic and accumulate at the interface.10 The bicone was positioned with the upper edge at the air/tea interface and the cup was capped with a cover housing a wet sponge. The ISR setup is shown in Fig. 2. | ||
| Fig. 2 Image of the used ISR setup with close-up of the custom made bicone at the air/tea interface and attached biofilm after 9 days of kombucha biofilm growth. | ||
The formation of kombucha biofilms was measured for 7 days by transient ISR time sweeps, i.e. sinusoidal oscillation of the bicone at constant frequency of 1 rad s−1 and strain amplitude of 0.5%. In separate experiments, frequency sweeps of the formed kombucha biofilms were performed every 24 h for 7 days by ranging the oscillation frequency from 0.1–100 rad s−1 at constant strain amplitude of 0.5%. Amplitude sweeps were performed to confirm that the measurements were performed in the linear viscoelastic regime (≤1% at day 7). The obtained raw data was corrected with respect to the subphase (kombucha broth) viscosity,23 which was determined before and after ISR experiments with a double gap geometry DG 26.7 and remained constant at 0.01 Pa s. The room temperature was controlled to 22 °C.
3. Results and discussion
Kombucha fermentation has been well characterized regarding microbial1,3–6 or biochemical1,14–17 dynamics. However, most analyses rely on periodic sampling and ex situ analysis. Particularly the cellulosic biofilm is limited to harvesting and ex situ determination of its weight, height, structure, and composition.9–11 ISR on the other hand allows the transient in situ determination of kombucha biofilm growth kinetics and mechanical properties. Fig. 3A shows the first 48 hours of four representative biofilm growth curves of equally prepared kombucha. The interfacial shear storage G′i and loss moduli G′′i increase over time and reach a plateau value which denotes the first step of biofilm formation. The formation kinetics and interfacial viscoelasticity of early kombucha biofilms are in good agreement with those reported for pure K. xylinus24 and other bacteria biofilms investigated by ISR.25–33 This initial increase is generally attributed to the adsorption of bacteria and surface active substances in the growth medium at the interface. In case of kombucha, the presence of proteins in the inoculum can be presumed,16 which indeed adsorb at similar time-scales and network strength at low concentrations.34 It is apparent from Fig. 3A that the initial kombucha biofilm formation showed significant variability within experiments, with plateau values ranging within one decade from 10−3 to 10−2 Pa m and time scales ranging from 3 to 24 hours to attain the plateau. Also, both the interfacial storage G′i or loss modulus G′′i may dominate in this phase. It is likely that the bacteria and protein concentration in the inoculum varied within experiments, leading to variations in initial biofilm formation. Another potential explanation is the competition between different bacteria at the interface. It is known that different bacteria show specific patterns in ISR depending on bacteria hydrophobicity, metabolic activity, and potential release of biosurfactants.26–28,35 In presence of multiple bacteria species the growth pattern depends on which bacteria dominates the interface, and accordingly, the initial density ratio of bacteria.31,32 We therefore assume that the variability in initial kombucha biofilm formation derives from variations in bacteria density and composition in the inoculum.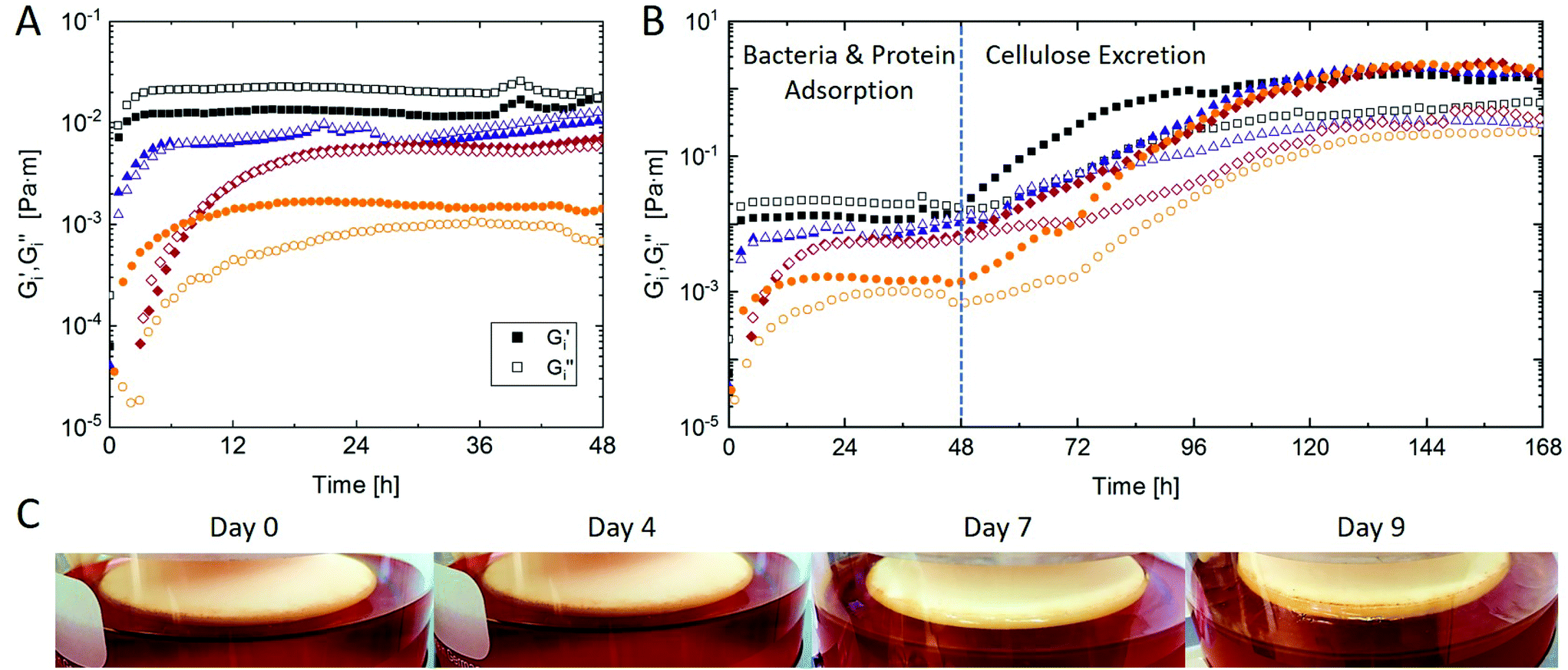 | ||
| Fig. 3 Interfacial shear storage G′i and loss G′′i moduli of four representative kombucha biofilms as a function of time for (A) the initial 48 h and (B) 7 days of biofilm growth. The dotted line indicates the onset of cellulose excretion by K. xylinus. Determined by ISR at constant frequency 1 rad s−1 and amplitude 0.5% at 22 °C. (C) Images of kombucha biofilms over time in the ISR setup. All samples were prepared equally with 100 g L−1 sucrose and inoculated with 10 vol% starter culture. | ||
Fig. 3B shows the kombucha biofilm growth curves for 7 days of fermentation. After the plateau following initial biofilm formation there is a secondary increase in interfacial shear moduli which can be attributed to bacterial cellulose excretion, primarily by K. xylinus. It has been reported that up to 90% of bacteria in kombucha biofilms are acetic acid bacteria, and ≈50% are Komagataeibacter.5 Besides cellulose, some acetic acid bacteria may further excrete fructan.36 The onset of cellulose excretion was found after 48 hours under the used conditions, in agreement with previous findings.10 Hence, kombucha biofilm formation is a two-step process characterized by initial bacteria and protein adsorption followed by cellulose excretion after ≈2 days. The onset of cellulose production was astonishingly reproducible within different measurements and independent of the initial biofilm formation which showed comparably high variations as discussed above. Also, G′i values after 7 days were very similar at 2 Pa m and G′′i values varied in a narrow range of 0.2–0.6 Pa m. The good reproducibility of the onset and mechanical properties could make ISR particularly useful to determine the dynamics and quantity of bacterial cellulose production in kombucha biofilms as a function of e.g. sucrose content, bacteria composition, or temperature.
The obtained ISR values are in good agreement with viscoelasticity reported for pure K. xylinus biofilms,24 and about 1–2 orders of magnitude higher than reported for other biofilms.25–31,33 Such secondary increases in interfacial moduli are well known from bacteria that secrete exopolysaccharides or proteins once in the stationary phase.24,25,29,33 A similar two step evolution of ISR moduli was also reported for fluid interfaces with secondarily adsorbed polysaccharides.37 In a previous study investigating pure K. xylinus biofilms, Rühs et al.24 found the onset of cellulose production already after 24 hours. This may indicate a delay in cellulose production in kombucha due to interfacial competition with other bacteria. However, Rühs et al.24 used an optimized culture medium containing mannitol which is the most efficient carbon source for K. xylinus cellulose production.9,38 Further, significant differences may occur with K. xylinus strains.39 The kombucha biofilm is only apparent to the naked eye after 6–7 days of fermentation, as visualized in Fig. 3C. Hence, ISR allows the determination of kombucha biofilm properties before it is visible and may be physically harvested.
The increase in interfacial viscoelasticity of kombucha biofilms over 7 days is further visualized by frequency sweeps in Fig. 4A, demonstrating the evolution from weakly viscoelastic biofilms in early stages to gel-like biofilms with frequency independent moduli after cellulose excretion. The kombucha biofilm continues to grow into the characteristic 3D cellulose film, which however cannot be assessed by ISR due to an increasing brittleness, i.e. decreasing linear viscoelastic regime, as visualized by amplitude sweeps in Fig. 4B. For the analysis of kombucha biofilms beyond 7 days of fermentation, biofilm harvesting and ex situ bulk rheology or mechanical testing may be more suitable, as previously employed for other cellulosic biofilms.40–42
 | ||
| Fig. 4 (A) Interfacial frequency and (B) amplitude sweeps of kombucha biofilms at different time points. Determined by ISR at constant amplitude 0.5% and frequency 1 rad s−1, respectively, at 22 °C. | ||
4. Conclusions
The use of ISR allows the in situ determination of kombucha biofilm growth and mechanical properties, and has revealed new aspects of kombucha growth dynamics. Kombucha biofilm formation was found to be a two step process. Initially, the adsorption of bacteria and surface active proteins induces an increase in interfacial rheological moduli which attain a first plateau. Biofilm growth kinetics and plateau values exhibit significant variability in this first phase, probably due to variations in bacteria density and composition within experiments. In the second phase, kombucha biofilm mechanical properties further increase due to the onset of bacterial cellulose excretion by K. xylinus. Both the onset of the secondary phase and final biofilm mechanical properties were acutely reproducible and independent of the first phase. Hence, ISR provides transient in situ information on kombucha biofilm dynamics and mechanical properties, and is particularly useful to investigate early stages of biofilm formation or determine the onset, kinetics, and amount of bacterial cellulose production.Conflicts of interest
There are no conflicts to declare.Acknowledgements
The authors thank Stefan Gstöhl for design of the bicone and Leandro Portenier for providing the original kombucha SCOBY.References
- M. Sievers, C. Lanini, A. Weber, U. Schuler-Schmid and M. Teuber, Microbiology and fermentation balance in a Kombucha beverage obtained from a tea fungus fermentation, Syst. Appl. Microbiol., 1995, 18, 590–594 CrossRef.
- R. Jayabalan, R. V. Malbaša, E. S. Lončar, J. S. Vitas and M. Sathishkumar, A review on kombucha tea-microbiology, composition, fermentation, beneficial effects, toxicity, and tea fungus, Compr. Rev. Food Sci. Food Saf., 2014, 13, 538–550 CrossRef PubMed.
- C. H. Liu, W. H. Hsu, F. L. Lee and C. C. Liao, The isolation and identification of microbes from a fermented tea beverage, Haipao, and their interactions during Haipao fermentation, Food Microbiol., 1996, 13, 407–415 CrossRef.
- A. L. Teoh, G. Heard and J. Cox, Yeast ecology of Kombucha fermentation, Int. J. Food Microbiol., 2004, 95, 119–126 CrossRef CAS PubMed.
- S. Chakravorty, S. Bhattacharya, A. Chatzinotas, W. Chakraborty, D. Bhattacharya and R. Gachhui, Kombucha tea fermentation: Microbial and biochemical dynamics, Int. J. Food Microbiol., 2016, 220, 63–72 CrossRef CAS PubMed.
- S. A. Villarreal-Soto, S. Beaufort, J. Bouajila, J. P. Souchard and P. Taillandier, Understanding Kombucha Tea Fermentation: A Review, J. Food Sci., 2018, 83, 580–588 CrossRef CAS PubMed.
- Y. Yamada, P. Yukphan, H. T. L. Vu, Y. Muramatsu, D. Ochaikul and Y. Nakagawa, Subdivision of the genus Gluconacetobacter Yamada, Hoshino and Ishikawa 1998: The proposal of Komagatabacter gen. nov., for strains accommodated to the Gluconacetobacter xylinus group in the α-Proteobacteria, Ann. Microbiol., 2012, 62, 849–859 CrossRef CAS.
- P. Ross, R. Mayer and M. Benziman, Cellulose biosynthesis and function in bacteria, Microbiol. Rev., 1991, 55, 35–58 CrossRef CAS PubMed.
- V. T. Nguyen, B. Flanagan, M. J. Gidley and G. A. Dykes, Characterization of cellulose production by a Gluconacetobacter xylinus strain from Kombucha, Curr. Microbiol., 2008, 57, 449–453 CrossRef CAS PubMed.
- W. N. Goh, A. Rosma, B. Kaur, A. Fazilah, A. A. Karim and R. Bhat, Fermentation of black tea broth (kombucha): I. Effects of sucrose concentration and fermentation time on the yield of microbial cellulose, Int. Food Res. J., 2012, 19, 109–117 CAS.
- W. N. Goh, A. Rosma, B. Kaur, A. Fazilah, A. A. Karim and R. Bhat, Microstructure and physical properties of microbial cellulose produced during fermentation of black tea broth (Kombucha). II., Int. Food Res. J., 2012, 19, 153–158 CAS.
- D. Laavanya, S. Shirkole and P. Balasubramanian, Current challenges, applications and future perspectives of SCOBY cellulose of Kombucha fermentation, J. Cleaner Prod., 2021, 126454 CrossRef CAS.
- K. Neffe-Skocińska, B. Sionek, I. Ścibisz and D. Kołożyn-Krajewska, Acid contents and the effect of fermentation condition of Kombucha tea beverages on physicochemical, microbiological and sensory properties, CyTA–J. Food, 2017, 15, 601–607 CrossRef.
- B. Bauer-Petrovska and L. Petrushevska-Tozi, Mineral and water soluble vitamin content in the Kombucha drink, Int. J. Food Sci. Technol., 2000, 35, 201–205 CrossRef CAS.
- R. Jayabalan, S. Marimuthu and K. Swaminathan, Changes in content of organic acids and tea polyphenols during kombucha tea fermentation, Food Chem., 2007, 102, 392–398 CrossRef CAS.
- R. Jayabalan, K. Malini, M. Sathishkumar, K. Swaminathan and S. E. Yun, Biochemical characteristics of tea fungus produced during kombucha fermentation, Food Sci. Biotechnol., 2010, 19, 843–847 CrossRef CAS.
- R. V. Malbaša, E. S. Lončar, J. S. Vitas and J. M. Čanadanović-Brunet, Influence of starter cultures on the antioxidant activity of kombucha beverage, Food Chem., 2011, 127, 1727–1731 CrossRef.
- C. Fu, F. Yan, Z. Cao, F. Xie and J. Lin, Antioxidant activities of kombucha prepared from three different substrates and changes in content of probiotics during storage, Food Sci. Biotechnol., 2014, 34, 123–126 Search PubMed.
- M. Afsharmanesh and B. Sadaghi, Effects of dietary alternatives (probiotic, green tea powder, and Kombucha tea) as antimicrobial growth promoters on growth, ileal nutrient digestibility, blood parameters, and immune response of broiler chickens, Comp. Clin. Pathol., 2014, 23, 717–724 CrossRef CAS.
- U. Hiremath, M. Vaidehi and B. Mushtari, Effect of fermented tea on the blood sugar levels of NIDDM subjects, Indian Pract., 2002, 55, 423–425 Search PubMed.
- J. M. Kapp and W. Sumner, Kombucha: a systematic review of the empirical evidence of human health benefit, Ann. Epidemiol., 2019, 30, 66–70 CrossRef PubMed.
- D. Morales, Biological activities of kombucha beverages: The need of clinical evidence, Trends Food Sci. Technol., 2020, 105, 323–333 CrossRef CAS.
- P. Erni, P. Fischer, E. J. Windhab, V. Kusnezov, H. Stettin and J. Läuger, Stress- and strain-controlled measurements of interfacial shear viscosity and viscoelasticity at liquid/liquid and gas/liquid interfaces, Rev. Sci. Instrum., 2003, 74, 4916–4924 CrossRef CAS.
- P. A. Rühs, F. Storz, Y. A. López Gómez, M. Haug and P. Fischer, 3D bacterial cellulose biofilms formed by foam templating, npj Biofilms Microbiomes, 2018, 4, 21 CrossRef PubMed.
- C. Wu, J. Y. Lim, G. G. Fuller and L. Cegelski, Quantitative analysis of amyloid-integrated biofilms formed by uropathogenic escherichia coli at the air-liquid interface, Biophys. J., 2012, 103, 464–471 CrossRef CAS PubMed.
- P. A. Rühs, L. Böni, G. G. Fuller, R. F. Inglis and P. Fischer, In situ quantification of the interfacial rheological response of bacterial biofilms to environmental stimuli, PLoS One, 2013, 8(11), e78524 CrossRef PubMed.
- P. A. Rühs, L. Böcker, R. F. Inglis and P. Fischer, Studying bacterial hydrophobicity and biofilm formation at liquid-liquid interfaces through interfacial rheology and pendant drop tensiometry, Colloids Surf., B, 2014, 117, 174–184 CrossRef PubMed.
- T. De Wouters, C. Jans, T. Niederberger, P. Fischer and P. A. Rühs, Adhesion potential of intestinal microbes predicted by physico-chemical characterization methods, PLoS One, 2015, 10, 1–17 CrossRef PubMed.
- E. C. Hollenbeck, J. C. Fong, J. Y. Lim, F. H. Yildiz, G. G. Fuller and L. Cegelski, Molecular determinants of mechanical properties of V. cholerae biofilms at the air-liquid interface, Biophys. J., 2014, 107, 2245–2252 CrossRef CAS PubMed.
- L. Qi and G. F. Christopher, Role of flagella, type IV pili, biosurfactants, and extracellular polymeric substance polysaccharides on the formation of pellicles by Pseudomonas aeruginosa, Langmuir, 2019, 35, 5294–5304 CrossRef CAS PubMed.
- C. Abriat, N. Virgilio, M. C. Heuzey and F. Daigle, Microbiological and real-time mechanical analysis of bacillus licheniformis and pseudomonas fluorescens dual-species biofilm, Microbiology, 2019, 165, 747–756 CrossRef CAS PubMed.
- C. Abriat, K. Enriquez, N. Virgilio, L. Cegelski, G. G. Fuller, F. Daigle and M. C. Heuzey, Mechanical and microstructural insights of Vibrio cholerae and Escherichia coli dual-species biofilm at the air-liquid interface, Colloids Surf., B, 2020, 188, 110786 CrossRef CAS PubMed.
- S. Pandit, M. Fazilati, K. Gaska, A. Derouiche, T. Nypelö, I. Mijakovic and R. Kádár, The exo-polysaccharide component of extracellular matrix is essential for the viscoelastic properties of Bacillus subtilis biofilms, Int. J. Mol. Sci., 2020, 21, 6755 CrossRef CAS PubMed.
- J. Bergfreund, M. Diener, T. Geue, N. Nussbaum, N. Kummer, P. Bertsch, G. Nyström and P. Fischer, Globular protein assembly and network formation at fluid interfaces: Effect of oil, Soft Matter, 2021, 17, 1692–1700 RSC.
- G. Subbiahdoss and E. Reimhult, Biofilm formation at oil-water interfaces is not a simple function of bacterial hydrophobicity, Colloids Surf., B, 2020, 194, 111163 CrossRef CAS PubMed.
- S. La China, G. Zanichelli, L. De Vero and M. Gullo, Oxidative fermentations and exopolysaccharides production by acetic acid bacteria: a mini review, Biotechnol. Lett., 2018, 40, 1289–1302 CrossRef CAS PubMed.
- P. Bertsch, A. Thoma, J. Bergfreund, T. Geue and P. Fischer, Transient measurement and structure analysis of protein-polysaccharide multilayers at fluid interfaces, Soft Matter, 2019, 15, 6362–6368 RSC.
- D. R. Ruka, G. P. Simon and K. M. Dean, Altering the growth conditions of Gluconacetobacter xylinus to maximize the yield of bacterial cellulose, Carbohydr. Polym., 2012, 89, 613–622 CrossRef CAS PubMed.
- M. Gullo, A. Sola, G. Zanichelli, M. Montorsi, M. Messori and P. Giudici, Increased production of bacterial cellulose as starting point for scaled-up applications, Appl. Microbiol. Biotechnol., 2017, 101, 8115–8127 CrossRef CAS PubMed.
- X. Gao, Z. Shi, C. Liu, G. Yang, I. Sevostianov and V. V. Silberschmidt, Inelastic behaviour of bacterial cellulose hydrogel: In aqua cyclic tests, Polym. Test., 2015, 44, 82–92 CrossRef CAS.
- P. A. Rühs, K. G. Malollari, M. R. Binelli, R. Crockett, D. W. Balkenende, A. R. Studart and P. B. Messersmith, Conformal bacterial cellulose coatings as lubricious surfaces, ACS Nano, 2020, 14, 3885–3895 CrossRef PubMed.
- H. Oliver-Ortega, S. Geng, F. X. Espinach, K. Oksman and F. Vilaseca, Bacterial cellulose network from kombucha fermentation impregnated with emulsion-polymerized poly(methyl methacrylate) to form nanocomposite, Polymer, 2021, 13, 664 CAS.
| This journal is © The Royal Society of Chemistry 2021 |