
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceEffects of mixed agro-residues (corn crop waste) on lignin-degrading enzyme activities, growth, and quality of Lentinula edodes
Shuai Xua,
Fei Wanga,
Yongping Fua,
Dan Lia,
Xiaozhong Sunb,
Changtian Lia,
Bing Song *a and
Yu Li*a
*a and
Yu Li*a
aEngineering Research Centre of Chinese Ministry of Education for Edible and Medicinal Fungi, Jilin Agricultural University, Changchun 130118, P. R. China. E-mail: song19800123@126.com; fungi966@126.com; Tel: +86-13500881489
bJilin Province Product Quality Supervision Test Institute, Changchun 130000, P. R. China
First published on 6th March 2020
Abstract
Using agro-residues in bioreactors to produce mushrooms is an important component of eco-agriculture. Here, we tested the cultivation of Lentinula edodes with corn cob (CC) and corn straw (CS), and then evaluated the resulting enzyme activities, agronomic traits, textures and nutrient compositions of mushrooms. Laccase (T1 formula, 3.26 g U−1) and carboxymethyl cellulase (T3 formula, 1.01 g U−1) activities were the highest at the time for a complete substrate colonization stage (TCSC), while acidic xylanase activity was the highest (CK formula, 4.05 g U−1) in the mushroom block to color-turned (TMBCT) stage. The biological efficiency of growth on the T6 formula was 8.82% higher than growth on the CK formula, wherein the low C/N ratio of the substrate had an obvious negative effect on yield while the mass ratio of pileus (MRP) of fruiting bodies did not change with mixed substrates. No significant differences were observed in mineral composition for CK formulas, but corn crop waste (CCW) formulas exhibited more optimal nutritional contents. A formula containing more corn cob and sawdust (SD) (sum of at least 70%) as the substrate can produce fruiting bodies with good hardness. These results indicate that the use of corn cobs as the main ingredient, mixed with sawdust and corn straw to grow L. edodes provides a more efficient use of agro-residues for growth. Thus, mixed agro-residue formulas have exceptional advantages in texture, nutrition of fruiting bodies, and yields.
1. Introduction
Fermentation activity is a product of the natural development of human civilization.1 Macrofungi are very important species that use mycelia to undergo solid-state fermentation (SSF) and produce characteristic fruiting bodies.2 Indeed, macrofungi grow on every type of organic matter and recycle substrate nutrients. Lentinula edodes, commonly known as shiitake mushrooms, are a highly sought after, edible, and valuable mushroom that are widely cultivated in China (10.43 million tons per year), Japan, and South Korea.3 L. edodes contains an abundance of eritadenine that can help maintain low blood pressure in humans.4 Likewise, vitamin B12 is the only known vitamin that contains metallic elements and is solely synthesized naturally by microorganisms, such as L. edodes. Indeed, it can be important for vegetarians to consume L. edodes, because vegetables do not contain vitamin B12 and its deficiency can cause anaemia.5 In addition, lentinan produced by L. edodes exhibits immunopotentiation effects and can exert anti-tumor activities by enhancing normal bodily immune functions, although it cannot directly kill tumor cells.6The highest plantation areas of maize exist in North America followed by Asia, Latin America, and Europe. There are over 3000 types of processed products made from maize. Concomitantly, significant amounts of corn crop waste (CCW) are produced from such activities. In China, agro-residues are abandoned to decay in fields or otherwise burned, thereby resulting in major air pollution.7,8 However, CCW, like other lignocellulose materials, are excellent substrates that can grow white-rot fungi. At the same time, timber has been strictly controlled by Chinese governments as a forest resource (index number: 000014672/2018-00466). In 2018, the Chinese government amended and implemented regulations to strictly control the utilization of forest resources, which then increased the cost of L. edodes cultivation. Thus, the popularization of cultivating shiitake via agricultural waste is urgently needed. In addition, the conversion speeds of biomass must be accelerated from scientific theory to application technology. Indeed, several studies have conducted experiments using different types of agricultural wastes to grow mushrooms.9–11 Nevertheless, traditional methods for selecting formulas have become outdated.
In this study, major agricultural wastes from northeast China including CS (corn straw) and CC (corn cob), were mixed with sawdust (SD) to grow shiitake mushrooms. The overall aim of the study was to identify variation in enzyme activities and bioconversion efficiencies during SSF with L. edodes grown on different CCW formulas, the enzyme secreted by mushroom through the SSF play an interesting role in the detoxification of solid-state waste materials which can be used for different applications according to the circular economy, such as these enzymes could be used in the improvement of fodder quality.10 To evaluate the inherent differences in growth between straw formulas, several types of data were measured including three enzymatic activities (laccase, carboxymethyl cellulose, and acidic xylanases), texture that is typically used to evaluate food quality in food science,12 and agronomic character to identify agronomic characteristic differences. The overall approach used here was to evaluate the specific characteristics (morphology and physiology) of mushroom growth to identify optimal approaches for cultivation.
2. Materials and methods
2.1 Mushroom cultivation and preparation
The L. edodes strain 135 (cultivated) was obtained from the Agricultural Culture Collection of China (Beijing) (Number: ACCC50903). A stock culture was maintained on substrate comprising 78% sawdust, 20% wheat bran, 1% gypsum, and 1% lime that was incubated for 30 d at 25 °C in the dark. Spawn were stored at 4 °C until further use.9Six different formula were used to grow L. edodes comprising corn cob, corn straw, and sawdust, incorporated with a simplex-lattice design. The CK formula comprised 80% sawdust, 18% wheat bran, 1% gypsum, 0.5% lime, and 0.5% sugar.13 The moisture content of each formula was maintained at 60% with a natural pH (the pH was 4.5 ± 0.5 after mycelium full colonization of bags). Further, corn cob, corn straw, and sawdust varied in proportion, while the concentrations of auxiliary materials were consistent across treatments (Table 1).
| Formulas | CC | CS | SD | Wheat bran | Gypsum | Lime | Sugar | Carbon source content/g per 100 g | Nitrogen source content/g per 100 g | C/N value |
|---|---|---|---|---|---|---|---|---|---|---|
| a CC: corn cob; CS: corn straw; SD: sawdust. | ||||||||||
| CK | 0 | 0 | 80 | 18 | 1 | 0.5 | 0.5 | 36.04 | 0.85 | 42.29 |
| T1 | 10 | 40 | 30 | 18 | 1 | 0.5 | 0.5 | 42.10 | 1.12 | 37.68 |
| T2 | 25 | 10 | 45 | 18 | 1 | 0.5 | 0.5 | 42.60 | 0.92 | 46.27 |
| T3 | 25 | 25 | 30 | 18 | 1 | 0.5 | 0.5 | 42.14 | 1.02 | 41.33 |
| T4 | 10 | 10 | 60 | 18 | 1 | 0.5 | 0.5 | 43.01 | 0.92 | 46.79 |
| T5 | 10 | 25 | 45 | 18 | 1 | 0.5 | 0.5 | 42.55 | 1.01 | 41.80 |
| T6 | 40 | 10 | 30 | 18 | 1 | 0.5 | 0.5 | 42.18 | 0.92 | 45.75 |
Well-mixed substrates were placed in mushroom bags (2000 g, polyethylene bag 17 cm × 55 cm × 0.03 cm) was then autoclaved at 121 °C for 3 h, and one interior low-pressure polyethylene bag (15 cm × 55 cm × 0.05 cm) was used after inoculated under aseptic conditions.14
2.2 Enzyme activity assays and the determination of agronomic traits
![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 g for 10 min at 4 °C and the supernatants were used to determine enzyme activity.
000 g for 10 min at 4 °C and the supernatants were used to determine enzyme activity.The three enzymatic activities were investigated using commercial test kits (Suzhou Comin Biotechnology Company Limited, China) following the manufacturer's instruction. Briefly, laccase activity was determined using the 2,2′-azinobis-(3-ethylbenzthiazoline-6-sulphonate) (ABTS) method for measuring free radical increases.15 Acidic xylanase activity was measured by evaluating acidic xylanase reduction of xylan to monosaccharides under acidic conditions, resulting in a color-change of 3,5-dinitrosalicylic acid (DNS) with the monosaccharides in aqueous solutions that can then be used to calculate acidic xylanase activity.16 Cellulase activity was measured using anthrone colorimetry.17
2.3 Analysis of texture and nutrients
3 Results and discussion
3.1 Enzyme activities
Changes in enzymatic activities among different mixture formulations were the highest during the TCSC period, and decreased during TMBCT. Activities were also higher in the third stage, but were still lower than in the first stage. Changes in enzymatic activity among different formulations also varied due to different enzyme types.Laccase enzymatic activity was much higher than that of acid xylanase and carboxymethyl cellulase in the three growth stages of L. edodes. No significant differences in activity were observed among formulations during the same period, although considerable differences were observed among different stages. The highest differences were observed for the T1 formulation, which exhibited an activity of 3.26 U g−1 during the first stage and only 0.05 U g−1 during the TMBCT stage.
In contrast to laccase, acidic xylanase activities exhibited high variation. Only a few fungi can produce acidic xylanase under acidic conditions to synthesize sugars for growth. The T1, T2, T3, and T4 formula activities were the highest in the TCSC periods, while the T5, T6, and CK formula activities were the highest in the TMBCT stage. The highest activity during the TMBCT stage for the CK formula was 4.05 U g−1, and the lowest was 0.4 U g−1 for mushrooms grown on the T2 formula. The higher levels of sawdust and corn cob in the T5, T6, and CK formulas could alter some of the physical and chemical properties in the bags, and thus explain the differences in acid xylanase activity.
Lastly, carboxymethyl cellulase activities behaved similarly to that of laccase. Cellulase high–low curves were observed during the three growth stages of L. edodes, but were much lower than that of laccase. The maximum cellulase activity was observed for the T4 formulation, but was only 0.95 U g−1 (Fig. 1).
 | ||
| Fig. 1 Radar map of enzymatic activity trends (units: U g−1). Each hexagon-separated unit in the picture is a base-10 logarithm (Y′ = log10Y). The numbers in the figure are the enzymatic activities during the TCSC, TMBCT, and TIH for growth on seven formulas. Laccase activity, acid xylanase activity and carboxymethyl cellulase activity are shown from left to right. | ||
Evaluation of three enzyme types produced by white rot fungus during SSF,20,21 indicated that laccase and carboxymethyl cellulase were important in the first growth stage of L. edodes. The above enzymes decompose lignin and cellulose from sawdust and corn straw.22,23 During the mycelial growth stage of L. edodes, abundant enzymes were secreted to decompose substrates and fuel mycelial growth.24 Mycelium activity was weak during TMBCT indicating that the enzymes produced by mycelium could only be used to maintain living cells and complete physiological structure changes on cell surfaces, bags as nutrient supplier has reached the saturation of mycelial growth. Mycelial activities have become intense for complete the reproductive growth process during TIH, their secreting more laccase and carboxymethyl cellulase to decompose lignocellulose in the substrate to form fruiting bodies. Differences in endocrine enzyme activities in bags obviously differed due to treatment ingredients. T1 (10% CC, 40% CS, 30% SD) formula with the highest corn straw proportion and has the highest laccase activity (3.26 U g−1-TCSC; 0.32 U g−1-TIH), CK formula without straw supplement had the lowest enzyme activity at TCSC stage, its indicated that straw supplement in mixed substrate may lead to more laccase secretion when mycelia flourishing. Acid xylanase activity results showed that the more sawdust in the substrate, the higher the acid xylanase activity at all three stage (CK:80% SD > T4:60% SD > T1:30% SD). Although relation between carboxymethyl cellulases and supplement was not observed that mixture could completely affect the mycelial secretion of L. edode.
3.2 Effects of substrates on L. edodes fruiting bodies
Agronomic traits were evaluated to identify the CCW decomposition capacity of L. edodes for different formulas. BE% trended with formulas as follows: T3 < T1 < T2 < CK < T5 < T4 < T6, wherein the C/N ratios of the T1 and T3 treatments were lower than that of CK (80% SD), while the C/N ratios of T6 (40% CC, 10% CS, and 30% SD) was higher than (8.18%) CK. The PT of the T2 and T3 treatments reached 1.25 cm, with T2, T3, and T5 all exhibiting thicker PT. In addition, the PD values for these three treatments were small. The longest SL value was observed for T3 at 3.26 cm. Nevertheless, statistical analysis indicated little difference between SL and SDT values among mushrooms grown with different formulas. Pileus weights also varied with single fruiting body sizes and MRP varied independent of formula treatments and generally remained in the range of 0.85 ± 0.1. Increasing weights of fruiting bodies varied with PT and PD over a particular range (e.g., similar weights exhibited a relationship between high PD and low PT). Thus, the density of L. edodes fruiting bodies remained unchanged (Table 2 and Fig. 2).| Formulas | CK | T1 | T2 | T3 | T4 | T5 | T6 |
|---|---|---|---|---|---|---|---|
| a BE (%) = (weight of fresh mushrooms harvested/substrate dry matter content) × 100%. Pileus thickness (PT), pileus diameter (PD), stipe length (SL), stipe diameter (SDT), mass ratio of pileus (MRP) = (weight of pileus/weight of single fruiting body) × 100%. Different letters (a–d) in the same column and rank indicate significant differences (P < 0.05). | |||||||
| PT (cm) | 1.20 ± 0.04ab | 1.22 ± 0.04ab | 1.25 ± 0.08a | 1.25 ± 0.03a | 1.20 ± 0.04ab | 1.21 ± 0.03ab | 1.17 ± 0.03b |
| PD (cm) | 5.29 ± 0.56abc | 4.77 ± 0.33bc | 4.94 ± 0.18abc | 4.42 ± 0.95c | 5.94 ± 0.88a | 5.23 ± 0.56abc | 5.76 ± 0.52ab |
| SL (cm) | 3.19 ± 0.19ab | 2.99 ± 0.32ab | 3.01 ± 0.26ab | 3.26 ± 0.17a | 2.83 ± 0.23b | 3.07 ± 0.10ab | 3.24 ± 0.31ab |
| SDT (cm) | 1.34 ± 0.09ab | 1.26 ± 0.06b | 1.39 ± 0.08ab | 1.31 ± 0.14ab | 1.28 ± 0.10a | 1.34 ± 0.07ab | 1.45 ± 0.10a |
| BE (%) | 54.77 ± 1.85c | 50.47 ± 1.28d | 53.82 ± 2.02c | 49.94 ± 1.34d | 61.22 ± 1.35ab | 59.67 ± 2.26b | 63.59 ± 2.13a |
| MRP | 0.85 | 0.86 | 0.86 | 0.86 | 0.86 | 0.84 | 0.85 |
 | ||
| Fig. 2 Scatter plot of agronomic characteristics of fruiting bodies. A (cm): pileus diameters, B (cm): pileus thicknesses, C (g): single fruiting body weights. | ||
For statistical analysis of agronomic traits, we focused on comparing the yield distribution and the weight of single fruiting bodies produced by each formula.25 The first and second flush significantly contributed to the ultimate BE% values (Fig. 3). The yield for the T6 formula was slightly higher than those of other formulas in the first and second flushes. Although the yield from the CK formula was not clearly high in the 1st and 2nd flushes, the yield of the 3rd flush was higher than those for the other formulas. The traditional formula exhibited an average yield, while the stalk formula in the experiment exhibited insufficient stamina. It should be noted that the requirements of mushroom cultivation in factories differs from that of farmers. Factories try to obtain high quality mushrooms over short times (e.g. the 1st and 2nd flushes), while farmers often look for further benefits and harvest until the 4th flush.26 (Fig. 3).
 | ||
| Fig. 3 Column map of yield distributions (units: g per bag). Yield distributions for the third, second, and first flushes are shown from top to bottom, respectively. Solid lines in columns shown margins of error. To facilitate comparisons, data are provided in the table below the graph. | ||
Suboptimal mushroom sizes produced in straw formulations affects mushroom prices.27 Thus, mushroom size should be considered in addition to yield28 when considering optimal growth formula. The T1 and T2 formulas did not provide the highest yields, but did produce larger fruiting bodies (Fig. 4). The median weight of the fruiting body of fresh mushrooms for the T2 formula was 23.9 g, and only the T4 formula exhibited a higher value than this. Although the T4, T5 and T6 formulas produced higher yields, the fresh mushroom fruiting bodies produced by these formulas were smaller, and most of the fruiting bodies ranged from 16.8 g and 20 g. The fresh mushroom weights produced by high yield formulas tended to vary little, while the fresh mushroom weights produced by formulas with lower yield varied considerably. Consequently, if large mushrooms are desired by consumers, the T1 and T5 growth formula are the best choices. However, the quality of small mushrooms produced by the T6 formula can often sell at higher prices. Scatter plot analysis revealed the reasons underlying the high yield of T6 (Fig. 4).
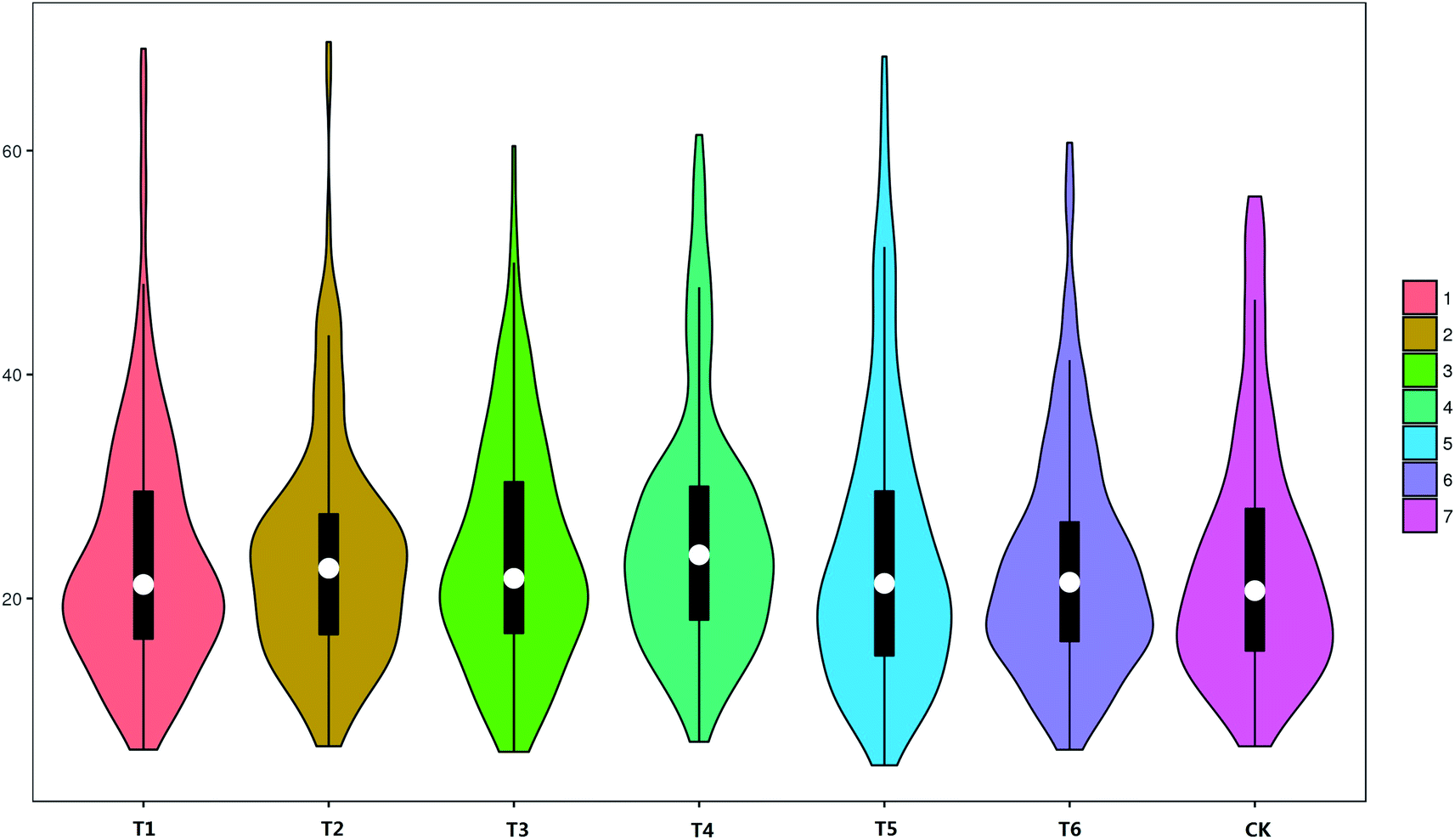 | ||
| Fig. 4 Scatter plot of single fruiting body weights. Ranges of values for weights of T1, T2, T3, T4, T5, T6 and CK formulas. Abscissa: formulas. Ordinate: weight of single fruiting body (units: g). The middle white point is the median, the upper edge of the black matrix is the upper quartile, and the lower edge is the lower quartile. | ||
Previous study showed that the C/N ratio significantly impacted yields.29 In this research, T1 formula have the lowest BE%, which also have the lowest C/N. However, T2 with the highest C/N yield is not the highest, in the mixture substrate, the T2, T5, T6, appear to be more favorable because they C/N ratio stabilizing at 45.00 ± 1.00, C/N ratio of mixture should stable within a certain range, and lead to a high BE%. The results of yield distribution showed that supplement for mixture substrate varies from purposes, the traditional formula exhibits advantages in long-term mushroom production capacity in early stages that often meet industrial requirements (costs raise as longer harvesting period). Although the yield of T1, T2 and T5 formulas did not reach the highest level, they did produce larger fruiting bodies (weight of single fresh fruiting body was larger than 60 g). The yield of T4 and T6 formula was higher than others. The scatter plot analysis reveals that the reason for the high yield is that the weight of single fruiting body varies in a small range near the median (scatter plot appears to be enlarged in the middle). The yield in mixture substrate varies inversely with the extent weight (high extent leads to low yield). No interesting results were observed in fruiting bodies traits, because they are intensely vulnerable to changes in environment condition. But, MRP was firstly presented and found that its numerical value stabilized at 0.85 ± 0.01 and did not change with formulas. MRP may respond to changes in strains, MMR as a parameter can apply it to screening high-quality strains. MRP may be the golden ratio in L. edodes.
3.3 Effects of substrates on L. edodes texture
Texture is an artificial taste recognition property that can help describe food characteristics.30 The formulas produced mushrooms with hardness in the trend of T2 > T4 > T6 > CK > T5 > T3 > T1. The hardness of mushrooms grown with the T1 formula (10% CC, 40% CS, and 30% SD) was only 2744.46 g and 55% lower than those grown with the T2 formula (25% CC, 10% CS, and 45% SD). The springiness index values of mushrooms grown with the CK and T5 formulas were 61.31% and 61.08%, which were higher than mushrooms grown with other formulations. The chewiness of T1 mushrooms was two-fold smaller than CK mushrooms, and springiness varied little (within 5%), indicating that the resilience of fresh mushrooms grown by the T1 formula was the lowest. Hardness and chewiness are two indicators requiring additional attention. Lower chewiness leads to lower energy consumption. In addition, the hardness of mushrooms is related to the cellulose and lignin content of the growth substrates.31 The T1 formula contained the largest amount of corn straw, while the T6 formula contained the highest amount of corn cob. The T4 formula had the highest amount of sawdust besides the traditional formula. The hardness and granularity of sawdust and corn cob materials are much higher than for corn straw, and thus the lignin content of sawdust and corn cob is much higher than that of corn straw. When the mixture formulas only contained 10% corn straw, the hardness of L. edodes fruiting bodies was higher (Table 3).| CK | T1 | T2 | T3 | T4 | T5 | T6 | |
|---|---|---|---|---|---|---|---|
| a Different letters (a–d) in the same column and rank indicate significant differences (P < 0.05). | |||||||
| Hardness (g) | 4815.97 ± 1467.59ab | 2744.66 ± 290.56c | 6182.59 ± 850.60a | 3957.09 ± 295.30bc | 5505.19 ± 1177.98ab | 4344.50 ± 1083.57bc | 5369.88 ± 635.48ab |
| Springiness (%) | 61.31 ± 0.65a | 56.84 ± 1.31bc | 61.94 ± 0.49a | 59.34 ± 0.63ab | 56.67 ± 3.11c | 61.08 ± 1.82a | 59.82 ± 0.05a |
| Chewiness (g sec) | 200251.58 ± 65175.23ab |
106461.60 ± 13460.38c |
255290.80 ± 34417.27a |
159877.35 ± 13609.95bc |
215178.18 ± 55364.95ab |
198023.46 ± 20016.41ab |
218776.22 ± 24932.66ab |
| Resilience (g) | 2962.26 ± 931.25abc | 1563.74 ± 201.05d | 3824.99 ± 496.27a | 2349.83 ± 200.20cd | 3274.68 ± 720.12 ab | 2868.39 ± 226.30bc | 3211.97 ± 377.54abc |
The taste evaluation of SSF products-L. edodes – often represent the characteristic of mixture substrates. However, the process is cumbersome and complex, it would be convenient to replace this process with texture test. The small particle size of corn straw affects gas flow in the bags, thereby affecting the complex process of fruiting body formation.32 Consequently, mixed formulas produced mushrooms with higher hardness than did CK. The addition of corn cobs in the bags significantly affected the hardness and resilience of fresh mushrooms. Hardness is one of the main indices to evaluate mushroom growth, because higher hardness leads to better fresh mushroom shapes due to the long-distance transport of L. edodes.33,34
3.4 Effects of substrates on the nutritional and mineral compositions of L. edodes
The potential for shiitake to enrich mineral elements and nutrients was a primary focus of our study.35,36 The T6 formula produced mushrooms with the highest contents of calcium, manganese, and iron and also produced mushrooms with higher contents of other elements except selenium. Iron, copper, and zinc contents were the highest in T5 mushrooms (10% CC, 25% CS, and 45% SD). Sodium, magnesium, and manganese contents were the highest in the CK formula. When planting corn, chemical fertilizers increase the contents of harmful elements in the straw, thereby affecting elemental compositions in the fruiting bodies of mushrooms. CCW had no effect on the harmful element contents within L. edodes, but exhibited promotion of zinc and selenium contents, while harmful elements in these mushrooms remained at concentrations lower than the Chinese standard (National Standards of the People's Republic of China, GB 2762-2017).14L. edodes fruiting bodies contained low fat contents, but high crude protein and total sugar contents.37 The carbon and nitrogen contents of substrates affect the nutrition of fruiting bodies, and especially the characteristics of sugars and amino acids.14 Previous studies have shown that as the proportion of corn stalks increased, total and essential amino acid contents first increased and then decreased during mushroom growth.9 Sugar content ranged in the trend of: T2 > T4 > T3 > T1 > T5 > T6 > CK. In contrast, crude protein and fat contents did not significantly differ among groups. Although the C/N ratio of the CK formula is suitable for L. edodes growth, its carbon and nitrogen contents were low, leading to inferior nutrient compositions in the fruiting bodies compared with stalk formulas. Amino acid types were the same across formulas, but their abundances varied. Glu, Gly, Val, Ile, Leu, Phe, Lys, His, and Arg content was the highest in T2 mushrooms, while Ser, Glu, Gly, Ala, Val, Met, Ile, Leu, Tyr, Phe, Lys, His, and Arg content was lowest in T6 mushrooms (Table 4).
| Treatments | CK | T1 | T2 | T3 | T4 | T5 | T6 | |
|---|---|---|---|---|---|---|---|---|
| a Different letters (a–f) in the same column and rank indicate significant differences (P < 0.05). * Essential amino acids. | ||||||||
| Calcium/mg kg−1 | Major element | 107.7 ± 6.52bc | 98.94 ± 14.11bc | 89.48 ± 10.51c | 110.51 ± 14.75bc | 118.89 ± 5.21ab | 98.91 ± 6.92bc | 133.94 ± 23.89a |
| Sodium/mg kg−1 | 111.32 ± 2.06a | 104.40 ± 5.98abc | 101.56 ± 12.84abc | 97.76 ± 3.10bc | 94.13 ± 7.12c | 101.79 ± 3.08abc | 108.93 ± 8.23ab | |
| Magnesium/mg kg−1 | 1185.45 ± 65.71a | 1068.20 ± 58.83b | 1062.87 ± 62.47b | 1013.68 ± 22.17b | 1003.70 ± 109.77b | 1068.56 ± 48.67b | 1091.58 ± 58.55ab | |
| Manganese/mg kg−1 | Trace element | 16.55 ± 1.16a | 15.31 ± 0.89ab | 13.49 ± 1.24b | 14.53 ± 1.34ab | 15.51 ± 1.36ab | 15.25 ± 0.02ab | 16.24 ± 1.45a |
| Iron/mg kg−1 | 47.65 ± 0.44c | 118.26 ± 1.35b | 119.32 ± 6.24b | 47.66 ± 2.86c | 145.78 ± 6.24a | 144.71 ± 2.91a | 143.71 ± 1.97a | |
| Copper/mg kg−1 | 4.18 ± 0.03c | 7.05 ± 0.53a | 6.58 ± 0.89a | 6.75 ± 0.73a | 5.32 ± 1.08bc | 7.29 ± 0.42a | 6.32 ± 0.63ab | |
| Zinc/mg kg−1 | 63.90 ± 1.60d | 69.30 ± 1.96bc | 69.63 ± 1.44bc | 71.33 ± 2.19b | 65.84 ± 2.44cd | 76.54 ± 2.15a | 69.19 ± 3.34bc | |
| Selenium/mg kg−1 | 0.12 ± 0.01d | 0.12 ± 0.01cd | 0.16 ± 0.01b | 0.18 ± 0.01a | 0.13 ± 0.01cd | 0.14 ± 0.01c | 0.11 ± 0.01d | |
| Arsenic/mg kg−1 | Harmful element | 0.20 ± 0.02d | 0.31 ± 0.04b | 0.37 ± 0.02a | 0.41 ± 0.02a | 0.26 ± 0.02c | 0.33 ± 0.03b | 0.30 ± 0.01bc |
| Cadmium/mg kg−1 | 0.33 ± 0.02a | 0.29 ± 0.02a | 0.22 ± 0.03b | 0.33 ± 0.07a | 0.28 ± 0.03a | 0.30 ± 0.04a | 0.19 ± 0.02b | |
| Mercury/mg kg−1 | 0.02 ± 0.01b | 0.02 ± 0.01b | 0.02 ± 0.01b | 0.16 ± 0.01a | 0.04 ± 0.03b | 0.04 ± 0.02b | 0.02 ± 0.01b | |
| Lead/mg kg−1 | 0.17 ± 0.01b | 0.06 ± 0.01d | 0.06 ± 0.02d | 0.12 ± 0.01c | 0.08 ± 0.01d | 0.31 ± 0.01a | 0.08 ± 0.02d | |
| Total sugar% | Nutritional | 35.60 ± 1.39c | 38.30 ± 0.71ab | 39.00 ± 0.25a | 38.40 ± 1.12ab | 38.60 ± 1.23a | 38.00 ± 0.99ab | 36.10 ± 2.45bc |
| Fat% | 0.48 ± 0.03bc | 0.52 ± 0.01ab | 0.51 ± 0.04ab | 0.53 ± 0.02a | 0.52 ± 0.01ab | 0.51 ± 0.01abc | 0.47 ± 0.03c | |
| Crude protein% | 23.60 ± 0.72ab | 22.40 ± 1.19ab | 19.27 ± 0.55c | 21.80 ± 0.69b | 23.20 ± 0.58ab | 22.70 ± 1.28ab | 22.90 ± 0.77a | |
| Ser/g per 100 g | Amino acid | 0.44 ± 0.01bc | 0.48 ± 0.01a | 0.43 ± 0.01c | 0.43 ± 0.01c | 0.42 ± 0.03c | 0.47 ± 0.02ab | 0.27 ± 0.03d |
| Glu/g per 100 g | 2.10 ± 0.12a | 1.90 ± 0.18ab | 2.10 ± 0.39a | 2.09 ± 0.05a | 1.68 ± 1.07ab | 2.00 ± 0.24a | 1.13 ± 0.03b | |
| Gly/g per 100 g | 0.39 ± 0.01a | 0.38 ± 0.01ab | 0.39 ± 0.01a | 0.38 ± 0.01ab | 0.34 ± 0.01c | 0.35 ± 0.05bc | 0.23 ± 0.02d | |
| Ala/g per 100 g | 0.50 ± 0.10a | 0.37 ± 0.16bc | 0.48 ± 0.10ab | 0.46 ± 0.12ab | 0.43 ± 0.17ab | 0.46 ± 0.17ab | 0.31 ± 0.10c | |
| Val*/g per 100 g | 0.85 ± 0.06ab | 0.86 ± 0.02a | 0.87 ± 0.02a | 0.87 ± 0.03a | 0.81 ± 0.03c | 0.82 ± 0.01bc | 0.69 ± 0.02d | |
| Met/g per 100 g | 0.16 ± 0.01ab | 0.15 ± 0.01ab | 0.16 ± 0.01ab | 0.15 ± 0.01ab | 0.14 ± 0.01b | 0.17 ± 0.02a | 0.10 ± 0.01c | |
| Ile*/g per 100 g | 0.33 ± 0.01a | 0.31 ± 0.01ab | 0.33 ± 0.01a | 0.31 ± 0.02ab | 0.30 ± 0.01b | 0.32 ± 0.01ab | 0.19 ± 0.01c | |
| Leu*/g per 100 g | 0.56 ± 0.01a | 0.54 ± 0.02ab | 0.56 ± 0.03a | 0.54 ± 0.01ab | 0.51 ± 0.01b | 0.54 ± 0.01ab | 0.33 ± 0.02c | |
| Tyr/g per 100 g | 0.16 ± 0.01d | 0.18 ± 0.01bc | 0.18 ± 0.01bc | 0.20 ± 0.01a | 0.17 ± 0.01cd | 0.19 ± 0.01ab | 0.10 ± 0.01e | |
| Phe/g per 100 g | 0.36 ± 0.03a | 0.34 ± 0.02bc | 0.36 ± 0.01a | 0.34 ± 0.01bc | 0.32 ± 0.02b | 0.34 ± 0.01bc | 0.21 ± 0.01c | |
| Lys*/g per 100 g | 0.50 ± 0.05bc | 0.50 ± 0.02bc | 0.53 ± 0.02a | 0.51 ± 0.01bc | 0.47 ± 0.03b | 0.52 ± 0.01a | 0.29 ± 0.03c | |
| His/g per 100 g | 0.16 ± 0.01ab | 0.16 ± 0.01ab | 0.17 ± 0.01a | 0.17 ± 0.03a | 0.14 ± 0.02b | 0.17 ± 0.01a | 0.09 ± 0.02c | |
| Arg/g per 100 g | 0.42 ± 0.01a | 0.42 ± 0.01a | 0.41 ± 0.03a | 0.41 ± 0.02a | 0.38 ± 0.01b | 0.41 ± 0.01a | 0.24 ± 0.01c | |
| Pro/g per 100 g | 0.5 ± 0.03de | 0.56 ± 0.01a | 0.51 ± 0.01cd | 0.48 ± 0.01ef | 0.47 ± 0.01f | 0.54 ± 0.02ab | 0.53 ± 0.02bc | |
Metal toxicity in CCW could to be the limiting factors in L. edodes cultivation,38 but there was no significant difference between CK and CCW formulas. Carbon content in substrate directly affects total sugar content in fruiting bodies, content of total sugar in CK formula was only 35.60%, 8.72% lower than T2 (38.30%). The content of total sugar in the rest of all CCW formulas stabilized at 37.00% ± 1.00. Carbon and nitrogen content did not affect amino acid contents. Thus, the use of mixed CCS substrates to grow L. edodes will not affect the elemental composition of fruiting bodies, and preserves their excellent characteristics including low fat and high protein content, thereby amplifying the advantages of mixture formulations.
4. Conclusions
Cultivation of mushrooms on mixed substrates fully utilizes agro-residues, while excessive straw formulas do not favor the growth of mushrooms due to low C/N ratios. However, when corn cob contents are increased, the resulting CCW formulas exhibit greater advantages than the traditional formula. The specific disadvantages of using CCW is that it cannot maintain yield stabilities and the activities of acid xylanase changes with formulations, while laccase and carboxymethyl cellulase activity trends remain unchanged. Our analyses indicate that the T6 (40% CC, 10% CS, and 30% SD) formula can partially replace the traditional formula and provides an easily available substrate for L. edodes cultivation.Author contributions
SX, FW and YPF conceived and designed the experiments, performed the experiments, analysed the data, wrote the paper, and prepared the figures and/or tables; these authors contributed equally to this work. DL, XZS, and CTL conceived and designed the experiments. BS and YL conceived and designed the experiments, analysed the data, contributed reagents/materials/analysis tools, wrote the paper, prepare figures and/or tables, and reviewed drafts of the paper. All authors read and approved the final manuscript.Abbreviations
| BE% | Biological efficiencies |
| CC | Corn cob |
| CS | Corn straw |
| CCW | Corn crop waste |
| MRP | The mass ratio of pileus |
| PD | Pileus diameters |
| PT | Pileus thickness |
| SD | Sawdust |
| SDT | Stipe diameters |
| SL | Stipe lengths |
| SSF | Solid-state fermentation |
| TCSC | Time for complete substrate colonization |
| TIH | Time for inoculation to harvest |
| TMBCT | Time for the mushroom block to color-turned |
Conflicts of interest
The authors declare that they have no competing interests.Ethical approval
This article does not contain any studies with animals or human participants performed by any of the authors.Informed consent
Informed consent was obtained from all individual participants included in the study.Acknowledgements
This work was supported by the Special Fund for Agro-scientific Research in the Public Interest (No. 201503137), National Key Research and Development Program of China (No. 2017YFD0601002; 2018YFD1001001), Science and Technology Projects in Jilin Province Department of Education (JJKH20180670KJ; JJKH20160191KJ), the Program of Creation and Utilization of Germplasm of Mushroom Crop of the “111” Project (No. D17014) and University S & T Innovation Platform of Jilin Province for Economic Fungi (No. #2014B-1). We would like to thank Frederick Leo Sossah for minor improvements in English language and grammar of this paper.References
- F. Ertan, B. Balkan, S. Balka and T. Aktac, Biologia, 2006, 61, 657–661 CAS.
- P. D. Postemsky, M. A. Bidegain and G. Lluberas, Bioresour. Technol., 2019, 289, 121692 CrossRef CAS PubMed.
- S. I. Yoo, H. Y. Lee, K. Markkandan, S. Moon, Y. J. Ahn, S. Ji, J. Ko, S. J. Kim, H. Ryu and C. P. Chong, BMC Genomics, 2019, 20, 121 CrossRef PubMed.
- N. Asada, R. Kairiku, M. Tobo and A. Ono, J. Diet. Suppl., 2018, 16, 345–356 CrossRef PubMed.
- T. Bito, F. Teng, N. Ohishi, S. Takenaka, E. Miyamoto, E. Sakuno, K. Terashimae, Y. Yabuta and F. Watanabe, Mycoscience, 2014, 55, 462–468 CrossRef CAS.
- W. C. Chen, W. Li, Y. Yang, H. L. Yu, S. Zhou, J. Feng, X. B. Li and Y. F. Liu, J. Agric. Food Chem., 2015, 63, 795–801 CrossRef CAS PubMed.
- Y. Hu, L. S. Wang, Y. Li, Q. H. Li, C. L. Li, J. M. Chen, D. Weng and H. P. Li, Environ. Toxicol. Pharmacol., 2017, 56, 249–258 CrossRef CAS PubMed.
- L. Zhang, Y. Liu and L. Hao, Environ. Res. Lett., 2016, 11, 014014 CrossRef.
- L. S. Meng, Y. P. Fu, D. Li, X. Z. Sun, Y. Q. Chen, X. F. Li, S. Xu, X. Li, C. T. Li, B. Song and Y. Li, RSC Adv., 2019, 9, 5347–5353 RSC.
- M. Svagelj, M. Berovic, Y. F. Liu, J. S. Zhang, F. Pohleven, D. Klinar and A. Gregori, New Biotechnol., 2009, 26, 260–262 CrossRef PubMed.
- A. S. M. Sonnenberg, W. H. Hendriks, J. W. Cone and L. Mao, J. Sci. Food Agric., 2018, 98, 1232–1239 CrossRef PubMed.
- S. Gautam, A. Sharma and P. Thomas, Int. J. Food Sci. Nutr., 1998, 49, 5–10 CrossRef CAS.
- H. L. Yu, B. B. Lv and C. Y. Song, Acta Agric. Scand., 2016, 3, 63–66 Search PubMed.
- W. Li, W. C. Chen, Y. Yang, J. S. Zhang, J. Feng, H. L. Yu, S. Zhou, X. B. Li and Y. F. Liu, Int. J. Food Sci. Technol., 2017, 4, 981–991 CrossRef.
- R. Nihei, M. Usami, T. Taguchi and S. Amachi, J. Environ. Radioact., 2018, 127–134 CrossRef CAS PubMed.
- Q. Yang, Y. Gao, Y. Huang, Q. Xu, X. M. Luo, J. L. Liu and J. X. Feng, Process Biochem., 2015, 50, 571–581 CrossRef CAS.
- Y. X. Ma, J. C. Liu, M. Li, W. Tao, Z. C. Yu and Y. B. Liu, Aquacult. Res., 2019, 1–7 Search PubMed.
- G. T. Nie, Q. H Huang, X. Li, Y. Long, L. Zhang, L. D. Lian, B. Xie, Y. J. Yu and B. G. Xie, Acta Edulis Fungi, 2016, 23, 31–36 Search PubMed.
- H. Wang, N. Pampati, W. M. Mccormick and L. Bhattacharyya, J. Pharm. Sci., 2016, 105, 1851–1857 CrossRef CAS PubMed.
- A. N. Philippoussis, P. A. Diamantopoulou and G. I. Zervakis, World J. Microbiol. Biotechnol., 2003, 19, 551–557 CrossRef.
- M. Nagai, T. Sato, H. Watanabe, K. Saito, M. Kawata and H. Enei, Appl. Microbiol. Biotechnol., 2002, 60, 327–335 CrossRef CAS PubMed.
- P. D. Postemsky, M. A. Bidegain, R. Gonzálezmatute, N. D. Figlas and M. A. Cubitto, Bioresour. Technol., 2017, 231, 85–93 CrossRef CAS PubMed.
- A. Leonowicz, N. S. Cho, J. Luterek, A. Wilkolazka, M. Wojtas-Wasilewska, A. Matuszewska, M. Hofrichter, D. Wesenberg and J. Rogalski, J. Basic Microbiol., 2001, 41, 185–227 CrossRef CAS PubMed.
- X. J. Ni, X. J. Pan, Z. Y. Feng and R. G. Li, Acta Edulis Fungi, 1995, 4, 22–27 Search PubMed.
- J. Pinela, A. B. Omarini, D. Stojkovic, L. Barros, P. D. Postemsky, R. C. Calhelha, J. Breccia, M. Fernandez-Lahore, M. Sokovic and I. C. F. R. Ferreira, Food Res. Int., 2020, 132, 109094 CrossRef.
- E. Y. Jo, J. Y. Choi, J. W. Choi and J. H. Ahn, Mycobiology, 2013, 41, 210–213 CrossRef PubMed.
- F. Wu, L. W. Zhou, Z. L. Yang, T. Bau, T. H. Li and Y. C. Dai, Fungal Divers., 2019, 98, 1–76 CrossRef.
- B. Song, J. Q. Ye, F. L. Sossah, C. T. Li, D. Li, L. S. Meng, S. Xu, Y. P. Fu and Y. Li, AMB Express, 2018, 8, 46 CrossRef PubMed.
- C. N. Economou, P. A. Diamantopoulou and A. N. Philippoussis, Appl. Microbiol. Biotechnol., 2017, 101, 5213–5222 CrossRef CAS PubMed.
- R. Ghobadi, R. Mohammadi, J. Chabavizade and M. Sami, Adv. Biomed. Res., 2018, 7, 24 CrossRef PubMed.
- T. Jiang, Q. Wang, S. Xu, M. M. Jahangir and T. Ying, J. Sci. Food Agric., 2010, 90, 742–749 CAS.
- D. J. Royse and J. E. Sanchez-Vazquez, Bioresour. Technol., 2001, 76, 229–233 CrossRef CAS PubMed.
- Y. Y. Zhao, J. Y. Yi, J. F. Bi, Q. Q. Chen and B. Zhang, Dry. Technol., 2018, 1, 1–11 Search PubMed.
- T. Nitheranont, A. Watanabe and Y. Asada, Biosci., Biotechnol., Biochem., 2011, 75, 538–543 CrossRef CAS PubMed.
- M. B. Tsegay, A. G. Asgedom and M. H. Belay, Cogent Food Agric., 2019, 5, 1605013 Search PubMed.
- F. Zhou, W. X. Yang, M. K. Wang, Y. X. Miao, Z. W. Gui, Z. Li and D. L. Liang, Food Chem., 2018, 265, 182–188 CrossRef CAS PubMed.
- R. Naraian, R. K. Sahu, S. Kumar, S. K. Garg, C. S. Singh and R. S. Kanaujia, Environmentalist, 2009, 29, 1–7 CrossRef.
- A. Wahyono, Novianti, A. Bakri and K. Uzci, J. Phys. Conf., 2018, 953, 12120 CrossRef.
| This journal is © The Royal Society of Chemistry 2020 |