
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Photoprotection conferred by low level summer sunlight exposures against pro-inflammatory UVR insult
S. J.
Felton
ab,
B. B.
Shih
 ab,
R. E. B.
Watson
ab,
R.
Kift
c,
A. R.
Webb
c and
L. E.
Rhodes
*ab
ab,
R. E. B.
Watson
ab,
R.
Kift
c,
A. R.
Webb
c and
L. E.
Rhodes
*ab
aDermatology Research Centre, School of Biological Sciences, Faculty of Biology, Medicine and Health, The University of Manchester and Salford Royal NHS Foundation Trust, Manchester Academic Health Science Centre, The University of Manchester, Manchester, UK. E-mail: lesley.e.rhodes@manchester.ac.uk; Tel: +44 (0)161 206 1150
bSalford Royal NHS Foundation Trust, Manchester Academic Health Science Centre, The University of Manchester, Manchester, UK
cSchool of Earth and Environmental Sciences, Faculty of Science and Engineering, The University of Manchester, Manchester, UK
First published on 19th May 2020
Abstract
Tanning (melanisation and epidermal thickening) is a photoprotective response to solar UVR exposure, but it's unclear to what degree low-level exposures induce this in light-skin individuals, or whether this modifies the histological inflammatory response to UVR. Objectives were to examine if, in light-skin people, a simulated summer's casual sunlight exposures induces (i) melanogenesis, (ii) epidermal thickening and (iii) demonstrable protection against both clinical (erythema) and histological (neutrophil infiltration) impacts of higher-level, pro-inflammatory UVR challenge. A UVR intervention study was designed to simulate a summer's brief sunlight exposures (95% UVA, 5% UVB) as can provide sufficient vitamin D. Ten healthy adults of phototype II, median 47 years (range 30–59 years), 2 male/8 female, received 1.3 SED 3× weekly for 6 weeks, and were subsequently challenged with 2× personal MED of UVB on small areas of UVR-exposed and UVR-protected buttock skin. Skin erythema and pigmentation were measured spectrophotometrically. Punch biopsies were taken from (i) unexposed skin (ii) skin following the ×18 low-level UVR exposures and (iii) skin at 24 h following the 2 × MED challenge, with skin sections evaluated for epidermal thickness, and for neutrophil infiltration by immunohistochemistry. The 6-weeks’ UVR exposures significantly increased skin pigmentation, skin lightness (L*) reducing from 69.37 (SD 2.8) to 65.52 (2.33) at course-end (p < 0.001), and stratum corneum thickness rising from 29.3 (9.59) to 41.5 (12.7)μm (p < 0.05); there was no influence on neutrophil numbers. Following the pro-inflammatory (2× MED) UVR challenge, there was a small (18%) reduction in erythema but a proportionately greater (71%) reduction in neutrophil infiltration in skin prior-exposed to the UVR course compared with photoprotected skin (both p < 0.05). Thus, findings add to information on risk-benefit of low-level sunlight exposure. Even very light-skin people show measurable although modest photoprotective responses to repeated low-dose UVR; greater impact is seen on histological than clinical inflammation.
Introduction
Ultraviolet radiation (UVR) from summer sunlight incident on skin comprises approximately 95% UVA (320–400 nm) and 5% UVB (290–320 nm). Biological effects include inflammation, immunomodulation, vitamin D production and DNA damage,1,2 and also tanning responses, i.e. increased pigmentation and epidermal thickening, which can provide photoprotection.3In general, the pigmentation in delayed tanning develops 2–3 days after UVR-induced DNA damage, with p53 triggering increased eumelanin production and spatial rearrangement of melanosomes over basal keratinocyte nuclei, giving some protection against further damage.1,4,5 Eumelanin acts as a broad filter, reducing penetration of UVR, and has antioxidant properties.5 Tanning shows differential UVA and UVB effects and kinetics.6–8 Thus, solar-simulated radiation (SSR) may induce greater delayed pigmentation than UVB alone, suggesting synergism of UVA and UVB.9
Epidermal thickening is a further adaptation against UVR-induced damage, reducing the UVR reaching the dividing basal cells.10,11 Both UVB and, in much higher doses, UVA can cause epidermal hyperplasia, potentially through increased activity of the pentose shunt enzyme, glucose-6-phosphate dehydrogenase.12 More recently, whole genome microarray studies identified alterations in JAK-STAT, p53 and p38 stress-responsive pathways following SSR exposure in ex vivo human skin.13 In mice, acceleration of DNA, RNA and protein synthesis, with increased mitosis and basal cell turnover, is observed 24 h prior to UVB-induced epidermal hyperplasia.14 While UVR induces epidermal hyperplasia, particularly stratum corneum thickening, under certain conditions,15,16 the extent to which repeated low-level doses, equivalent to the brief summer sunlight exposures that attain vitamin D sufficiency,17,18 also influence epidermal thickness (viable epidermis i.e. epidermis excluding stratum corneum, and the stratum corneum itself) and melanisation in humans in vivo is unknown.
Acute UVR-induced inflammation, i.e. ‘sunburn’, clinically features as skin erythema, accompanied by pain, heat and swelling at higher exposure dose; UVB is orders of magnitude more effective than UVA at inducing erythema.19 This acute inflammatory response follows release of a cascade of inflammatory mediators, including nitric oxide, tumour necrosis factor-α, interleukins and sequential eicosanoid profiles.20–22 Alongside dilatation of dermal vasculature, upregulation of endothelial adhesion molecules and neutrophil chemoattractants results in neutrophil adherence to vascular endothelium, diapedesis and migration to UVR-exposed sites20,23,24 The erythema time course is typically maximal 24 h post-exposure and is accompanied by peak neutrophil infiltration,21,25 while maximal lymphocytic and macrophage influx is at 24–72 h.22,26 Neutrophils have manifold functions,25,27–30 including release of reactive oxygen species and proteolytic enzymes, immunomodulation, and phagocytosis of damaged cells.
Presently it's unclear what degree of photoprotection occurs against sunburn erythema after repeated UVR exposures under everyday conditions, and whether clinical erythema reflects the histological inflammatory response. This study aimed to examine the impact of a simulated summer's low-level sunlight exposures (6 week course; 95% UVA, 5% UVB) on epidermal melanisation and thickening in light skin (phototype II) Caucasians, and the potential photoprotection conferred against UVR-induced inflammatory insult, assessed both clinically as erythema and histologically by neutrophil infiltration.
Methods
Study volunteers
Healthy volunteers, Fitzpatrick skin type II, were recruited in Greater Manchester, UK, and the clinical study was performed in winter-time (January–February) in the Photobiology Unit, Dermatology Research Centre, Salford Royal Hospital, Greater Manchester, UK (53.5°N). Exclusion criteria were pregnancy, breastfeeding, taking photoactive medication or supplements containing vitamin D, history of skin cancer/photosensitivity, and sunbed-usage/sunbathing within 3 months prior to or during the study. This research comprised part of a larger study in which other UVR outcomes were assessed, with Ethical approval by North Manchester Research Ethics Committee (reference 09/H1014/73). The study adhered to Declaration of Helsinki principles.Minimal erythemal dose (MED) assessment
The MED, defined as the lowest dose of UVR to produce visually discernable erythema at 24 h, was assessed for each volunteer at baseline. A geometric series of 10 doses (7–80 mJ cm−2) of erythemally-weighted UVR was applied over 2 horizontal rows of buttock skin with a Waldmann UV 236B unit containing Waldmann CF-L 36 W/UV6 lamps (peak emission: 313 nm; range: 290–400 nm; Waldmann GmbH, Villinge-Schwenningen, Germany).Simulated summer sunlight exposures
Volunteers were given a 6-week course of UVR exposures, as described.18 A Philips HB588 irradiation cabinet (Eindhoven, The Netherlands), fitted with 11 Arimed B (Cosmedico GmbH, Stuttgart, Germany) alternating with 13 Cleo Natural (Philips, Eindhoven, The Netherlands) fluorescent tubes, delivered whole body UVR with emission spectrum close to UK summer sunlight (95% UVA: 320–400 nm, 5% UVB: 290–320 nm). Cabinet emission was characterized and monitored by spectroradiometry.17 Wearing protective eye goggles, T-shirts and shorts, volunteers lay prone on the sunbed with the canopy closed. One buttock was fully photoprotected whilst a cut-out panel revealed 10 cm2 of contralateral buttock skin throughout the UVR course.The simulated summer's sunlight was given 3× weekly in winter when ambient UVB is negligible at UK latitudes.31 An exposure of 1.3 standard erythemal dose (SED)32 was given at each visit. It took ∼6.5 minutes to deliver this dose after accurate measurement of cabinet UV irradiance;33 a constant UVR dose was maintained throughout the course by adjusting for any decrease in irradiance with increase in delivery time. This is equivalent to 13–17 minutes unshaded sunlight exposure on a clear June midday at 53.5°N, 6× weekly, accounting for non-simultaneous exposure of ventral and dorsal surfaces in sunlight, and the range of postures in daily life.34
2× MED UVB-challenge
At 72 h following completion of the course of simulated summer sunlight exposures, buttock skin was exposed to a pro-inflammatory challenge, individually-dosed for each volunteer to be twice their MED of UVB. This was applied using a Waldmann UV 236B unit to two 1 cm2 areas of buttock skin, one area that had received the simulated summer's UVR exposures, and a control site that was photoprotected.Skin color measurements
A Minolta CM-2500d hand-held spectrophotometer (Konica Minolta, Tokyo, Japan) was used for non-invasive measurement of skin color. Measurements were made at baseline and weekly from UVR-exposed and UVR-protected buttock skin, and 24 h after the 2× MED UVB-challenge. Data were recorded in standard three-dimensional Commission International de l’ Eclairage L*a*b* format; L* represents white-black differentiation, where L* of 100 is pure white and L* of 0 is pure black, a* values reflect the balance between green (negative) and increasing redness (positive) whilst b* is the differentiation between blue (negative) and yellow (positive).35 Readings were made in triplicate at each site. Individual typology angle (ITA) values were also calculated as the vector direction in the L*b* plane, as arctangent [(L* − 50)/b*] × (180/π) on exposed and unexposed skin throughout the UVR course.36,37A reflectance instrument (Erythema meter, Diastron, Andover, UK) was further used for non-invasive measurement of buttock skin erythema. Erythema index (EI) readings were taken in triplicate from 2× MED UVB-challenged skin, the UVR course-treated skin and adjacent non-irradiated skin.
To obtain background-corrected data, induced erythema was calculated by subtracting a* or EI readings taken on proximal, untreated buttock from those on 2× MED UVB-challenged buttock skin.
Skin sampling
At the end of the UVR course, four 4 mm punch biopsies were taken, immediately after the last 1.3 SED exposure from buttock skin that was: (i) unexposed and (ii) exposed to the 6-week simulated summer sunlight, and a second set of biopsies was taken 24 h after 2× MED UVB exposure from buttock skin that had been (iii) previously unexposed and (iv) exposed to the 6-week simulated sunlight. All samples were formalin-fixed and paraffin-embedded prior to histological analysis.Immunostaining
3 μm skin sections were prepared for immunohistological analyses including a solubilising step utilising 0.5% Triton X-100. After washing in tris-buffered saline (TBS), hydrogen peroxide (0.3% diluted in phosphate-buffered solution, PBS) was added to inhibit endogenous peroxidase before repeated washing. Primary antibody was added (neutrophil elastase NP57, Dako, Glostrup, Denmark; dilution 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :100) in primary antibody diluent (Diagnostic Biosystems, California, USA). For control, primary antibody was not added to one slide for each staining cycle. ImmPress reagent (Vector Laboratories, California, USA) was added before repeated rinsing. Slides were developed with NovaRed (Vector Laboratories, California, USA), rinsed and counterstained with Mayer's haematoxylin. Sections were dehydrated, and permanently mounted using DPX (Sigma-Aldrich Inc., St Louis, USA).
:100) in primary antibody diluent (Diagnostic Biosystems, California, USA). For control, primary antibody was not added to one slide for each staining cycle. ImmPress reagent (Vector Laboratories, California, USA) was added before repeated rinsing. Slides were developed with NovaRed (Vector Laboratories, California, USA), rinsed and counterstained with Mayer's haematoxylin. Sections were dehydrated, and permanently mounted using DPX (Sigma-Aldrich Inc., St Louis, USA).
Quantification of staining and epidermal thickness measurements
Images were captured and scanned using Pannoramic 250 Flash II, 3DHisTech Ltd (Budapest, Hungary) scanner. Counts were made of the number of positively staining cells per high power field (hpf; original magnification ×20), in triplicate per section, with three hpf taken per section (9 hpf per slide). Cellular epidermal (from stratum basale to distal stratum granulosum) and stratum corneum thicknesses were measured using the Pannoramic Viewer, nine per slide. Epidermal and dermal areas per hpf were measured using Image J 1.48 (National Institutes of Health, Bethesda, MD). The assessor (SJF) was blinded to sample identity for counting and measurements.Statistical analyses
Data analyses, specifically paired and unpaired t-tests, and linear regressions, were performed using SPSS statistical software (version 21.0.0; IBM SPSS Statistics, Portsmouth, UK) and GraphPad Prism (version 6; GraphPad Software, La Jolla, CA, USA). Neutrophil counts per epidermal area were logarithmically transformed to stabilise their variance. Results were considered statistically significant if p < 0.05.Results
Eight participants were female, two male. Median age was 47 years (range 30–59), BMI 25 (range 22–35) kg m−2 and MED 30 (22–54) mJ cm−2 of erythemally-weighted UVR (equivalent to 3 SED).A simulated summer's repeated low-level sunlight exposures significantly darkened the skin and thickened the stratum corneum
The repeated 1.3 SED exposures increased skin pigmentation, L* reducing 5.5%, from mean 69.37 (SD 2.8) at baseline to 65.52 (2.33) at course-end (p < 0.001; Fig. 1A). Similarly, ITA decreased from 52 (5.7)° at baseline to 41 (6.4)° post-course (p < 0.0001; Fig. 1B). | ||
| Fig. 1 Weekly skin color measurements taken from buttock skin during the simulated summer's sunlight exposure. ITA represents skin color on the L*–b* plane, based on spectrophotometer readings taken from buttock skin. The vertical axis is the skin lightness (L*) and the horizontal axis the yellow-blue component (b*). (a) L* and (b) ITA show significant darkening over the simulated summer sunlight exposures, from 69.37 (2.8) to 65.52 (2.33) and from 52 (5.7)° at baseline to 41 (6.4)° at course-end for the 10 volunteers (p < 0.001; p < 0.0001). ITA represents skin color on the L*–b* plane, based on spectrophotometer readings taken from buttock skin. The vertical axis is the skin lightness (L*) and the horizontal axis the yellow-blue component (b*). ITA values were higher at baseline (open circles) representing ‘very light’ skin color and were significantly decreased (closed circles) by the simulated summer‘s UVR exposures to ‘light’ or ‘intermediate’ color. Short horizontal bars in (a) denote the mean. *p < 0.0001. | ||
Mean viable epidermal thickness did not differ significantly between unexposed (39.1 [6.6] μm) and simulated sunlight-exposed skin (43.7 [8.1] μm; p > 0.05), whilst the stratum corneum thickened significantly from 29.3 [9.6] to 41.5 [12.7] μm (p < 0.05, Fig. 2A). The stratum corneum thickening did not correlate with darkening (change in L* or ITA; R2 = 0.06, R2 = 0.24 respectively, p > 0.05 for both).
 | ||
| Fig. 2 Effect of UVR exposure on epidermal thickness. (a) The 6 week course of simulated summer sunlight exposures significantly thickened the stratum corneum, from mean 29.3 (9.59) to 41.5 (12.7) μm (p < 0.05). Acute 2× UVB MED exposure did not provoke measurable change to stratum corneum thickness. (b) Simulated summer sunlight exposures apparently increased mean viable epidermal thickness from 39.1 (6.6) to 43.7 (8.1) μm (p > 0.05). Acute 2× MED UVB exposure apparently increased viable epidermal thickness from 43.7 (8.1) to 48.6 (6.4) μm at 24 h post-challenge (p = 0.05). Markers represent mean readings from each participant as measured by Pannoramic viewer (n = 8 for photoprotected skin due to lack of epidermis in 2 individuals, n = 10 for the remainder). Horizontal bars denote the overall mean. Thickening of the stratum corneum and the viable epidermis (black arrows) is seen between photoprotected skin (c) and that following the simulated summer sunlight (d). Original magnification ×20. *p < 0.05. | ||
A pro-inflammatory (2 × MED) UVB-challenge “thickened” viable epidermis at 24 h in previously unexposed and exposed skin
In previously unexposed skin, 2× MED UVB induced significant viable epidermal “thickening”, from 39.1 [6.6] to 48.1 [7.0] μm at 24 h post-challenge (p < 0.05, Fig. 2B). In the simulated summer sunlight-exposed skin post-course, the 2 × MED UVB increased the viable epidermal thickness from 43.7 [8.1] to 48.6 [6.4] μm (p = 0.05). However, thickening appeared due to epidermal oedema (spongiosis) not acanthosis, as keratinocyte spacing rather than number increased (Fig. 2C and D).In contrast to the impact on viable epidermis, the single acute 2× MED UVB-challenge did not provoke measurable change to stratum corneum thickness, in either previously unexposed skin or skin following the simulated sunlight course (p > 0.05 for both; Fig. 2A). Degree of epidermal thickening in previously unexposed skin was significantly, negatively correlated with erythema induced by the 2 × MED challenge (a*; R2 = 0.54; p < 0.05).
Repeated low level simulated summer sunlight exposures significantly reduced erythemal response to acute 2 × MED UVB challenge
2 × MED of UVB induced significantly greater erythema in previously unexposed skin compared with the simulated summer exposed skin: a* readings were 15% higher (9.9 [3.7] versus 8.5 [4.2]) and similarly EI values were 18% higher at 80.8 [31.2] versus 66.3 [32.1] (p < 0.05 for both within group comparisons; Fig. 3A and B).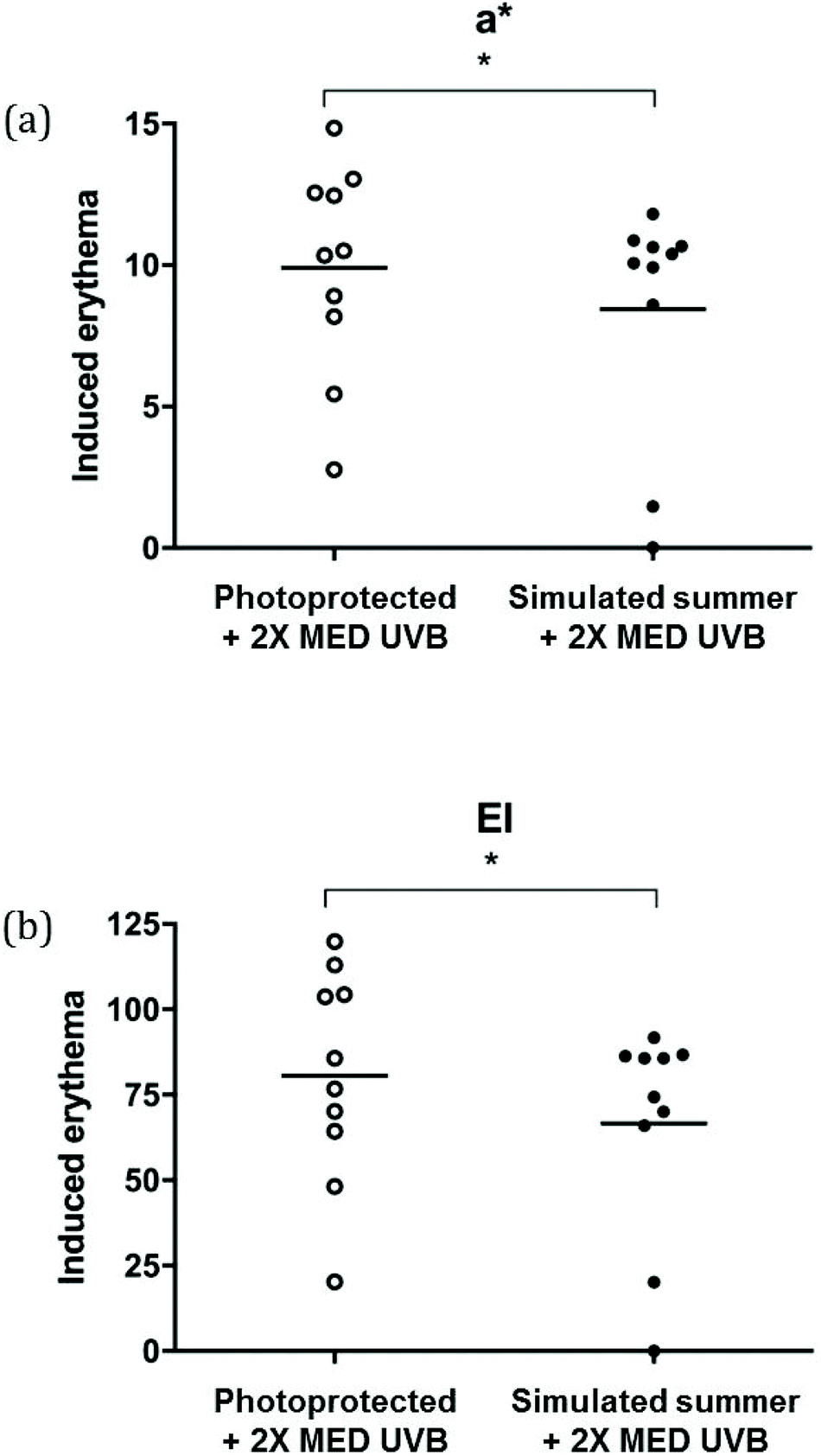 | ||
| Fig. 3 Cutaneous erythema following acute 2× MED UVB-challenge. Erythema induced by 2× MED challenge from all participants (n = 10) was greater in previously photoprotected skin, as demonstrated by (a) a* from L*a*b* scores (Konica Minolta spectrophotometer) and (b) erythema index (EI; Diastron reflectance instrument). To obtain background-corrected data, induced erythema was calculated by subtracting a* or EI readings taken on proximal, untreated buttock skin from those taken on 2× MED UVB-challenged buttock skin. Horizontal bars denote mean values. *p < 0.05. | ||
Repeated low level simulated summer sunlight exposures significantly reduced neutrophil infiltration response to acute 2 × MED UVB challenge
Dermal neutrophils were absent both in the unexposed buttock skin and that exposed to 6 weeks’ simulated sunlight course (Fig. 4A and B). However, following 2× MED UVB-challenge, the induced dermal neutrophil infiltration was 71% lower in the exposed than previously unexposed skin (21.4 [24.1] versus 73.5 [72.5] mm−2; between group T-test p < 0.05; Fig. 4C and D). | ||
| Fig. 4 Neutrophil infiltration under varying conditions of UVR exposure. (a)–(d) Neutrophil elastase NP57 staining (Dako, Denmark; black arrows), original magnification ×20 in (a) unexposed skin (b) immediately following the last 1.3 SED exposure of the 6-weeks’ simulated summer sunlight exposures (c) 24 h after 2× MED UVB exposure to skin that was photoprotected during the simulated summer's sunlight, (d) 24 h after 2× MED UVB exposure to skin that was previously exposed to the simulated summer's sunlight exposures (e) neutrophil counts per mm2 dermis. Neutrophils were absent in unexposed skin and skin that had been exposed to the simulated summer's sunlight. 24 h following acute 2× MED UVB-challenge, mean counts were significantly higher in previously unexposed skin, at 73.5 (72.5) mm−2 than in skin that had received the simulated summer's sunlight (21.4 [24.1] mm−2; p < 0.05). n = 8 for counts in photoprotected skin due to lack of epidermis/dermis in 2 individuals, n = 10 for the remainder. Horizontal bars denote the mean. *p < 0.05. | ||
In previously unexposed skin, the 2 × MED UVB-induced dermal neutrophil infiltration was positively correlated with baseline pallor (ITA, R2 = 0.40) and with erythemal response (R2 = 0.70, 0.72 for a*, EI respectively; p < 0.05 for all); no significant associations were seen post exposure course.
Discussion
This study made a detailed evaluation of the degree to which repeated low level sunlight exposures might protect against sunburn inflammation following higher level exposure in easy-sunburning white Caucasians, and found significant reduction in dermal neutrophil infiltration, as well as in erythema. A novel aspect of this study is its assessment of the photoprotection conveyed against neutrophilic infiltration as well as the erythemal aspect of UVR-induced inflammation, and following repeated UVR exposures mimicking real-life. These repeated low-level exposures to radiation similar to solar UVR reaching the earth's surface in summer (95% UVA, 5% UVB) are equivalent to ∼15 minutes UK (53.5°N) June midday sunlight, on most days of the week. We have previously demonstrated that these exposures do generate DNA damage (in the form of cyclobutane pyrimidine dimers), but that damage levels do not accumulate with repeated exposures, implying adequate repair mechanisms.18 These exposure levels approximate guidance on summer sunlight exposure for white skin people to achieve vitamin D sufficiency, and findings highlight a further impact of adopting such measures.17,38Typically, the more frequently assessed clinical erythema response to UVR is assumed to reflect cellular aspects of the early inflammatory response, where a neutrophilic infiltrate predominates,27 while this work indicates a proportionately larger effect on neutrophil numbers than on erythema. Photoprotection against sunburn erythema following the simulated summer's sunlight exposures was quantified as 15–18% reduction, using two measures (a* and EI). Interestingly, the repeated low-dose UVR exposures themselves did not induce a cutaneous inflammatory response, contrasting with findings from mouse models where multiple low-dose (sub-erythemal) UVR may cause low-level neutrophil infiltration.39 Rijken et al. (2005) found that after single SSR challenge, neutrophils were detectable in human skin only after a dose ≥1 MED;40 but the effect of multiple challenge on neutrophil infiltration appears previously unreported in humans. In mouse models,39 the increased neutrophils are thought to contribute to photoageing through releasing proteolytic enzymes, particularly neutrophil elastase and matrix metalloproteinases, that can damage elastin fibres and collagen networks, contributing to solar elastosis.
Neutrophils have several pro-inflammatory and immunomodulatory activities,41 including release of reactive oxygen species, and generation of chemotactic signals that attract further leucocytes to the site. Thus neutrophils regulate host responses to erythemal dose UVR injury. They also release Th2-associated cytokines including interleukins-4 and -10,27,29,37 which are immunosuppressive, and determine whether macrophages adopt a pro- or anti-inflammatory role. Moreover, they phagocytose damaged keratinocyte components, repairing UVR-induced injury.25,28 Consequently, while our demonstrated reduction in neutrophil infiltration following photoadaptation reflects overall reduced UVR-induced inflammation, it may also be associated with loss of beneficial, pro-resolution effects.
We quantified two key mechanisms that confer photoprotection, i.e. skin darkening and epidermal thickening. In our volunteers, who were typical skin type II including report of poor tanning ability, the low-level exposures nevertheless induced a small but statistically significant skin-darkening, with L* decrease of 5.5%. This is similar to a report of 8% L* decrease in five ‘lighter skinned’ Caucasians following 3-weeks SSR exposures.16 We also quantified ITA as a measure of skin darkening, which takes account of b* (yellow-green) spectrophotometer measurements, and thus has been considered a more accurate indicator of skin color.37 The simulated summer decreased ITA values by 11°, with individuals moving from a ‘very light’ to ‘light/intermediate’ category; although of small magnitude, correlations between ITA and sunburn cell-induction have been shown ex vivo.37 An apparent plateau occurred at ∼4 weeks, consistent with reported findings in darker, skin type V, people.42
Our data on epidermal thickness indicate that repeated low-level UVR (5% UVB, 95% UVA) exposures significantly thicken stratum corneum but have minimal impact on the viable epidermis, the latter showing an apparent though non-significant increase. This is consistent with the finding that 0.5 MED SSR (applied ×5 weekly) to skin of the back of individuals of skin type I–III, produced a 22% thickening in stratum corneum without change of viable epidermal thickness;15 our greater (42%) stratum corneum thickening possibly reflects skin type and irradiation site differences. In our homogeneous skin type subjects, stratum corneum thickening did not correlate with skin darkening. Interestingly, a UVR conduction study in ex vivo skin showed stratum corneum in phototypes I–III to filter >50% of incident radiation, whilst the filtering averaged only 20–30% in black skin samples.42 Adaptations of the stratum corneum may thus have a greater role in increasing photoprotection in lighter skin types, partially compensating for the lesser degree of skin darkening seen during a simulated summer's exposures in people of light versus dark skin types.18
Increased keratinocyte turnover is thought to be mostly responsible for stratum corneum thickening post-UVR with epidermal mitosis stimulated by increased proliferating cell nuclear antigen (PCNA) activity.43 Alterations in stratum corneum lipid content might also contribute, as increased serine palmitoyl transferase activity and associated increases in epidermal barrier lipids including ceramides, cholesterol and fatty acids have been detected following UVR exposure of murine skin.44
Following the single higher dose UVR challenge, epidermal oedema, i.e. spongiosis, appeared a key contributor to the acute epidermal “thickening” seen at 24 h. Potentially, the negative correlation we identified between epidermal thickness and erythema could be attributable to plasma leakage from capillaries limiting further capillary dilatation. Previous human studies of skin thickening post-UVR are scarce. A small study using high-frequency ultrasound imaging in four volunteers reported epidermal thickening to be maximal 48 h post single higher-dose (2–3 MED) UVR (40% UVB; 60% UVA) in healthy back skin; however, this lacked evidence that thickening was solely attributable to hyperplasia and not spongiosis.45 In mice, detailed time course studies from 3 to 72 h post-UVR showed PCNA expression initially confined to the basal skin layer, spreading upwards for 48 h post-UVR.43 This was accompanied by gradual increase in epidermal hyperplasia, with focal hyperplasia as early as 6 h, before becoming more uniform, and maximal 48 to 72 h post-UVR. Similarly, whilst metabolic changes occur within 24 h post-irradiation in mouse skin,13 it may take 24 to 72 h to detect change in thickness.
Findings of this study can be considered in the risk-benefit assessment of low level sunlight exposures in light-skin people: we show that repeated low-level simulated sunlight exposures, as sufficient for gaining vitamin D, induce modest measurable skin pigmentation accompanied by stratum corneum thickening, in light-skin people, accompanied by notable protection against higher dose UVR-induced neutrophil infiltration and slight, though statistically significant, protection against the accompanying erythema. While clinical erythema is the most frequently utilized UVR-inflammatory endpoint, including in study of photoprotective agents, this research highlights its proxy nature and the desirability of exploring histological outcomes.
Conflicts of interest
There are no conflicts of interest to declare.Acknowledgements
We are grateful to Dr S. M. Pilkington for assistance with the optimisation of immunohistochemical protocol, M. R. Rashid for assistance with immunohistochemical staining, and Prof. A. Vail for statistical guidance (all at The University of Manchester). This research was part-funded by Cancer Research UK, project C20668/A10007, and supported by the NIHR Manchester Biomedical Research Centre.References
- M. S. Eller, K. Ostrom and B. A. Gilchrest, DNA damage enhances melanogenesis, Proc. Natl. Acad. Sci. U. S. A., 1996, 93, 1087–1092 CrossRef CAS PubMed.
- T. Schwarz and A. Schwarz, Molecular mechanisms of ultraviolet radiation-induced immunosuppression, Eur. J. Cell Biol., 2011, 90, 560–564 CrossRef CAS PubMed.
- E. Kvam and R. M. Tyrrell, The role of melanin in the induction of oxidative DNA base damage by ultraviolet A irradiation of DNA or melanoma cells, J. Invest. Dermatol., 1999, 113, 209–213 CrossRef CAS PubMed.
- M. Gniadecka, H. C. Wulf, N. N. Mortensen and T. Poulsen, Photoprotection in vitiligo and normal skin. A quantitative assessment of the role of stratum corneum, viable epidermis and pigmentation, Acta Derm.-Venereol., 1996, 76, 429–432 CAS.
- M. Brenner and V. J. Hearing, The protective role of melanin against UV damage in human skin, Photochem. Photobiol., 2008, 84, 539–549 CrossRef CAS PubMed.
- C. F. Rosen, Y. Seki, W. Farinelli, R. S. Stern, T. B. Fitzpatrick, M. A. Pathak and R. W. Gange, A comparison of the melanocyte response to narrow band UVA and UVB exposure in vivo, J. Invest. Dermatol., 1987, 88, 774–779 CrossRef CAS PubMed.
- C. Duval, M. Régnier and R. Schmidt, Distinct melanogenic response of human melanocytes in mono-culture, in co-culture with keratinocytes and in reconstructed epidermis, to UV exposure, Pigm. Cell Res., 2001, 14, 348–355 CrossRef CAS PubMed.
- S. A. Miller, S. G. Coelho, B. Z. Zmudzka, H. F. Bushar, Y. Yamaguchi, V. J. Hearing and J. Z. Beer, Dynamics of pigmentation induction by repeated ultraviolet exposures: dose, dose interval and ultraviolet spectrum dependence, Br. J. Dermatol., 2008, 159, 921–930 CrossRef CAS PubMed.
- R. Wolber, K. Schlenz, K. Wakamatsu, C. Smuda, Y. Nakanishi, V. J. Hearing and S. Ito, Pigmentation effects of solar-simulated radiation as compared with UVA and UVB radiation, Pigm. Cell Melanoma Res., 2008, 21, 487–491 CrossRef PubMed.
- W. A. Bruls, H. Slaper, J. C. van der Leun and L. Berrens, Transmission of human epidermis and stratum corneum as a function of thickness in the ultraviolet and visible wavelengths, Photochem. Photobiol., 1984, 40, 485–494 CrossRef CAS PubMed.
- Y. Miyamura, S. G. Coelho, K. Schlenz, J. Batzer, C. Smuda, W. Choi, M. Brenner, T. Passeron, G. Zhang, L. Kolbe, R. Wolber and V. J. Hearing, The deceptive nature of UVA tanning versus the modest protective effects of UVB tanning on human skin, Pigm. Cell Melanoma Res., 2011, 24, 136–147 CrossRef PubMed.
- A. D. Pearse, S. A. Gaskell and R. Marks, Epidermal changes in human skin following irradiation with either UVB or UVA, J. Invest. Dermatol., 1987, 88, 83–87 CrossRef CAS PubMed.
- N. Mouchet, H. Adamski, R. Bouvet, S. Corre, Y. Courbebaisse, E. Watier, J. Mosser, C. Chesné and M. D. Galibert, In vivo identification of solar radiation-responsive gene network: role of the p38 stress-dependent kinase, PLoS One, 2010, 5, e10776 CrossRef PubMed.
- J. H. Epstein, K. Fukuyama and K. Fye, Effects of ultraviolet radiation on the mitotic cycle and DNA, RNA and protein synthesis in mammalian epidermis in vivo, Photochem. Photobiol., 1970, 12, 57–65 CrossRef CAS PubMed.
- R. M. Lavker, G. F. Gerberick, D. Veres, C. J. Irwin and K. H. Kaidbey, Cumulative effects from repeated exposures to suberythemal doses of UVB and UVA in human skin, J. Am. Acad. Dermatol., 1995, 32, 53–62 CrossRef CAS PubMed.
- S. de Winter, A. A. Vink, L. Roza and S. Pavel, Solar-simulated skin adaptation and its effect on subsequent UV-induced epidermal DNA damage, J. Invest. Dermatol., 2001, 117, 678–682 CrossRef CAS PubMed.
- L. E. Rhodes, A. R. Webb, H. I. Fraser, R. Kift, M. Durkin, D. Allan, S. J. O'Brien, A. Vail and J. L. Berry, Recommended summer sunlight exposure levels can produce sufficient (≥20 ng ml-1) but not the proposed optimal (≥32 ng ml-1) 25(OH)D levels at UK latitudes, J. Invest. Dermatol., 2010, 130, 1411–1418 CrossRef CAS PubMed.
- S. J. Felton, M. S. Cooke, R. Kift, J. L. Berry, A. R. Webb, P. M. Lam, F. R. de Gruijl, A. Vail and L. E. Rhodes, Concurrent beneficial (vitamin D production) and hazardous (cutaneous DNA damage) impact of repeated low-level summer sunlight exposures, Br. J. Dermatol., 2016, 175, 1320–1328 CrossRef CAS PubMed.
- I. Willis and L. Cylus, UVA erythema in skin: is it a sunburn?, J. Invest. Dermatol., 1977, 68, 128–129 CrossRef CAS PubMed.
- I. Strickland, L. E. Rhodes, B. F. Flanagan and P. S. Friedmann, TNF-alpha and IL-8 are upregulated in the epidermis of normal human skin after UVB exposure: correlation with neutrophil accumulation and E-selectin expression, J. Invest. Dermatol., 1997, 108, 763–768 CrossRef CAS PubMed.
- L. E. Rhodes, G. Belgi, R. Parslew, L. McLoughlin, G. F. Clough and P. S. Friedmann, Ultraviolet-B-induced erythema is mediated by nitric oxide and prostaglandin E2 in combination, J. Invest. Dermatol., 2001, 117, 880–885 CrossRef CAS PubMed.
- L. E. Rhodes, K. Gledhill, M. Masoodi, A. K. Haylett, M. Brownrigg, A. J. Thody and D. J. Tobin, The sunburn response in human skin is characterized by sequential eicosanoid profiles that may mediate its early and late phases, FASEB J., 2009, 23, 3947–3956 CrossRef CAS PubMed.
- P. H. Andersen, K. Abrams, P. Bjerring and H. Maibach, A time-correlation study of ultraviolet B-induced erythema measured by reflectance spectroscopy and laser Doppler flowmetry, Photodermatol., Photoimmunol. Photomed., 1991, 8, 123–128 CAS.
- P. G. Norris, J. N. Barker, M. H. Allen, K. M. Leiferman, D. M. MacDonald, D. O. Haskard and J. L. Hawk, Adhesion molecule expression in polymorphic light eruption, J. Invest. Dermatol., 1992, 99, 504–508 CrossRef CAS PubMed.
- M. B. Teunissen, G. Piskin, S. Di Nuzzo, R. M. Sylva-Steenland, M. A. de Rie and J. D. Bos, Ultraviolet B radiation induces a transient appearance of IL-4+ neutrophils, which support the development of Th2 responses, J. Immunol., 2002, 168, 3732–3739 CrossRef CAS PubMed.
- K. Kang, A. C. Gilliam, G. Chen, E. Tootell and K. D. Cooper, In human skin, UVB initiates early induction of IL-10 over IL-12 preferentially in the expanding dermal monocytic/macrophagic population, J. Invest. Dermatol., 1998, 111, 31–38 CrossRef CAS PubMed.
- J. L. Hawk, G. M. Murphy and C. A. Holden, The presence of neutrophils in human cutaneous ultraviolet-B inflammation, Br. J. Dermatol., 1988, 118, 27–30 CrossRef CAS PubMed.
- S. D. Shapiro, Neutrophil elastase: Path clearer, pathogen killer, or just pathologic?, Am. J. Respir. Cell Mol. Biol., 2002, 26, 266–268 CrossRef CAS PubMed.
- G. Piskin, J. D. Bos and M. B. Teunissen, Neutrophils infiltrating ultraviolet B-irradiated normal human skin display high IL-10 expression, Arch. Dermatol. Res., 2005, 296, 339–342 CrossRef CAS PubMed.
- P. L. Lee, H. van Weelden and P. L. B. Bruijnzeel, Neutrophil infiltration in normal human skin after exposure to different ultraviolet radiation sources, Photochem. Photobiol., 2008, 84, 1528–1534 CrossRef CAS PubMed.
- A. R. Webb and O. Engelsen, Calculated ultraviolet exposure levels for a healthy vitamin D status, Photochem. Photobiol., 2006, 82, 1697–1703 CrossRef CAS PubMed.
- B. L. Diffey, C. T. Jansen, F. Urbach and H. C. Wulf, The standard erythema dose: a new photobiological concept, Photodermatol., Photoimmunol. Photomed., 1997, 13, 64–66 CrossRef CAS PubMed.
- D. K. Taylor, A. V. Anstey, A. J. Coleman, B. L. Diffey, P. M. Farr, J. Ferguson, S. Ibbotson, K. Langmack, J. J. Lloyd, P. McCann, C. J. Martin, H. P. Menagé, H. Moseley, G. Murphy, S. D. Pye, L. E. Rhodes and S. Rogers, British Photodermatology Group. Guidelines for dosimetry and calibration in ultraviolet radiation therapy: a report of a British Photodermatology Group workshop, Br. J. Dermatol., 2002, 146, 755–763 CrossRef CAS PubMed.
- A. R. Webb, R. Kift, J. L. Berry and L. E. Rhodes, The vitamin D debate: translating controlled experiments into reality for human sun exposure times, Photochem. Photobiol., 2011, 87, 741–745 CrossRef CAS PubMed.
- A. R. Robertson, The CIE 1976 color-difference formulae, Color Res. Appl., 1977, 2, 7–11 CrossRef.
- A. Chardon, I. Cretois and C. Hourseau, Skin colour typology and suntanning pathways, Int. J. Cosmet. Sci., 1991, 13, 191–208 CrossRef CAS PubMed.
- S. Del Bino, J. Sok, E. Bessac and F. Bernerd, Relationship between skin response to ultraviolet exposure and skin color type, Pigm. Cell Res., 2006, 19, 606–614 CrossRef CAS PubMed.
- PHE (2017). Ultraviolet radiation, vitamin D and health: report of the independent advisory group on non-ionising radiation. https://www.gov.uk/government/publications/ultraviolet-radiation-and-vitamin-d-the-effects-on-health.
- H. Takeuchi, T. Gomi, M. Shishido, H. Watanabe and N. Suenobu, Neutrophil elastase contributes to extracellular matrix damage induced by chronic low-dose UV irradiation in a hairless mouse photoaging model, J. Dermatol. Sci., 2010, 60, 151–158 CrossRef CAS PubMed.
- F. Rijken, R. C. Kiekens and P. L. Bruijnzeel, Skin-infiltrating neutrophils following exposure to solar-simulated radiation could play an important role in photoageing of human skin, Br. J. Dermatol., 2005, 152, 321–328 CrossRef CAS PubMed.
- C. Nathan, Neutrophils and immunity: challenges and opportunities, Nat. Rev. Immunol., 2006, 6, 173–182 CrossRef CAS PubMed.
- K. H. Kaidbey, P. P. Agin, R. M. Sayre and A. M. Kilgman, Photoprotection by melanin-a comparison of black and Caucasian skin, J. Am. Acad. Dermatol., 1979, 1, 249–260 CrossRef CAS PubMed.
- A. Ouhtit, H. K. Muller, D. W. Davis, S. E. Ullrich, D. Mcconkey and H. N. Ananthaswamy, Temporal events in skin injury and the early adaptive responses in ultraviolet-irradiated mouse skin, Am. J. Pathol., 2000, 156, 201–207 CrossRef CAS PubMed.
- W. M. Holleran, Y. Uchida, L. Halkier-sorensen, A. Haratake, M. Hara, J. H. Epstein and P. M. Elias, Structural and biochemical basis for the UVB-induced alterations in epidermal barrier function, Photodermatol., Photoimmunol. Photomed., 1997, 13, 117–128 CrossRef CAS PubMed.
- H. Lopez, J. Z. Beer, S. A. Miller and B. Z. Zmudzka, Ultrasound measurements of skin thickness after UV exposure: a feasibility study, J. Photochem. Photobiol., B, 2004, 73, 123–132 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry and Owner Societies 2020 |