
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceNutritional quality and in vitro digestion of immature rice-based processed products
Kulwa F.
Miraji
 ab,
Anita R.
Linnemann
b,
Vincenzo
Fogliano
b,
Henry S.
Laswai
c and
Edoardo
Capuano
*b
ab,
Anita R.
Linnemann
b,
Vincenzo
Fogliano
b,
Henry S.
Laswai
c and
Edoardo
Capuano
*b
aTanzania Agricultural Research Institute, Ifakara Centre, Ifakara, Tanzania
bFood Quality and Design, Wageningen University and Research, Wageningen, The Netherlands. E-mail: edoardo.capuano@wur.nl
cMwenge Catholic University, Moshi, Tanzania
First published on 3rd September 2020
Abstract
Rice is commonly consumed as fully mature grain, but immature rice is considered to have better nutrient and technological properties. This is attributed to changes in content and profile of nutritional and functional compounds during maturation. This study assessed the effect of maturity on nutrient content of rice grains, and in vitro digestibility of starch and protein, for immature rice grains of TXD306 and Lawama varieties. The effect of processing of immature rice into so-called pepeta, traditionally produced from immature rice grains and widely consumed in Tanzania, was studied as well. The results showed reductions in lipid, protein, ash, thiamine, nicotinic acid, nicotinamide, and soluble and insoluble dietary fibre contents during rice grain development. However, no effect of maturity on in vitro starch and protein digestibility was observed. The contents of protein, ash, lipid, nicotinamide, iron, zinc, and total, soluble and insoluble dietary fibre were higher in pepeta from both varieties than in the corresponding rice grains. Protein digestibility of pepeta flour was 58.9% higher than that of cooked rice for variety TXD306, and 73.8% higher for Lawama. Differential scanning calorimetry indicated that starch of processed immature rice was completely gelatinized whereas its susceptibility to digestion in vitro was slightly lower than for cooked rice, possibly due to the higher cellular integrity retained after processing. These results demonstrate that pepeta-type processing improves the nutritional properties of rice and its potential use as a snack or ingredient in cereal-based formulas.
1. Introduction
Rice-based food products are widely consumed worldwide, providing up to 46% and 43% of dietary energy and dietary protein in the sub-Saharan region, respectively.1,2 Rice is often consumed as fully mature grains in the form of whole kernels after cooking using a particular amount of water or by boiling in excess water.3 However, several researches reported higher amounts of protein, reducing sugars, calcium, potassium, iron, β-carotene, vitamin C, and vitamin B2, B3 and B6 in immature cereal grains,4,5 making them nutritionally superior to fully mature grains. These variations in the accumulation of nutrients in developing seeds are related to cellular and physiological changes that occur between different development stages.6 The proteomic analyses revealed expression patterns (clusters) of proteins which are differentially expressed, and associated with starch synthesis and nutrients accumulation during rice grain development.7,8Besides positive nutritional benefits, immature cereal grains are considered to have better technological properties than fully mature grains due to changes in content and profile of functional compounds during maturation. Lin and Lai9 reported higher ratios of soluble dietary fibre (SDF) to total dietary fibre (TDF) in immature dehusked unpolished grains ranging from 9.4–17.2% compared to 5.8%–7.4% in fully mature dehusked unpolished grains. Moreover, the content of bioactive compounds, such as phenolics and flavonoids, is significantly higher in developing rice grains than in fully mature seeds.9,10 Similar studies on immature wheat have reported high contents of total dietary fibre, fructo-oligosaccharides and phytochemicals, such as phenolics and flavonoids.11,12
Wholegrain and polished rice are utilized in a very wide range of foods,13 including flaked rice. Flaked rice is a popular snack in rice-consuming countries. Pepeta, a locally produced flaked product from immature rice grains, is a sought-after snack in Tanzania for its popcorn taste and flavour. The traditional process of making pepeta involves roasting of harvested immature rice grains, dehusking and pounding of grains in a mortar to obtain flattened rice grains and husks, followed by winnowing to remove the husks prior to consumption (Fig. 1). The effect of cooking on the physical and nutritional behaviour of rice kernels is well-documented: swelling and crack formation due to hydration, starch gelatinization which enhances starch digestibility and the glycaemic index, and leaching of soluble nutrients like starch and vitamin of the B group in water.14,15 However, in pepeta a dry heating processing step is used, which bears resemblance to dry-heat parboiling (i.e. a technique involving rapid roasting of sufficiently soaked paddy.16 This technique is widely used in processing of flaked, puffed and expanded rice-based snacks.17 It increases the digestibility of starch in processed products compared to raw products,16–18 and perhaps the nutrient content as well due an increased total ash in rice- and wheat-based roasted products.19,20 These positive effects may also apply to pepeta processing, but this still needs to be confirmed by research. This study assessed the effect of maturation on the nutrient content of rice grain in the products of two domestic Tanzanian rice varieties. Effect of maturation on nutritional components on in vitro digestibility of pepeta starch and protein during grain development were also examined.
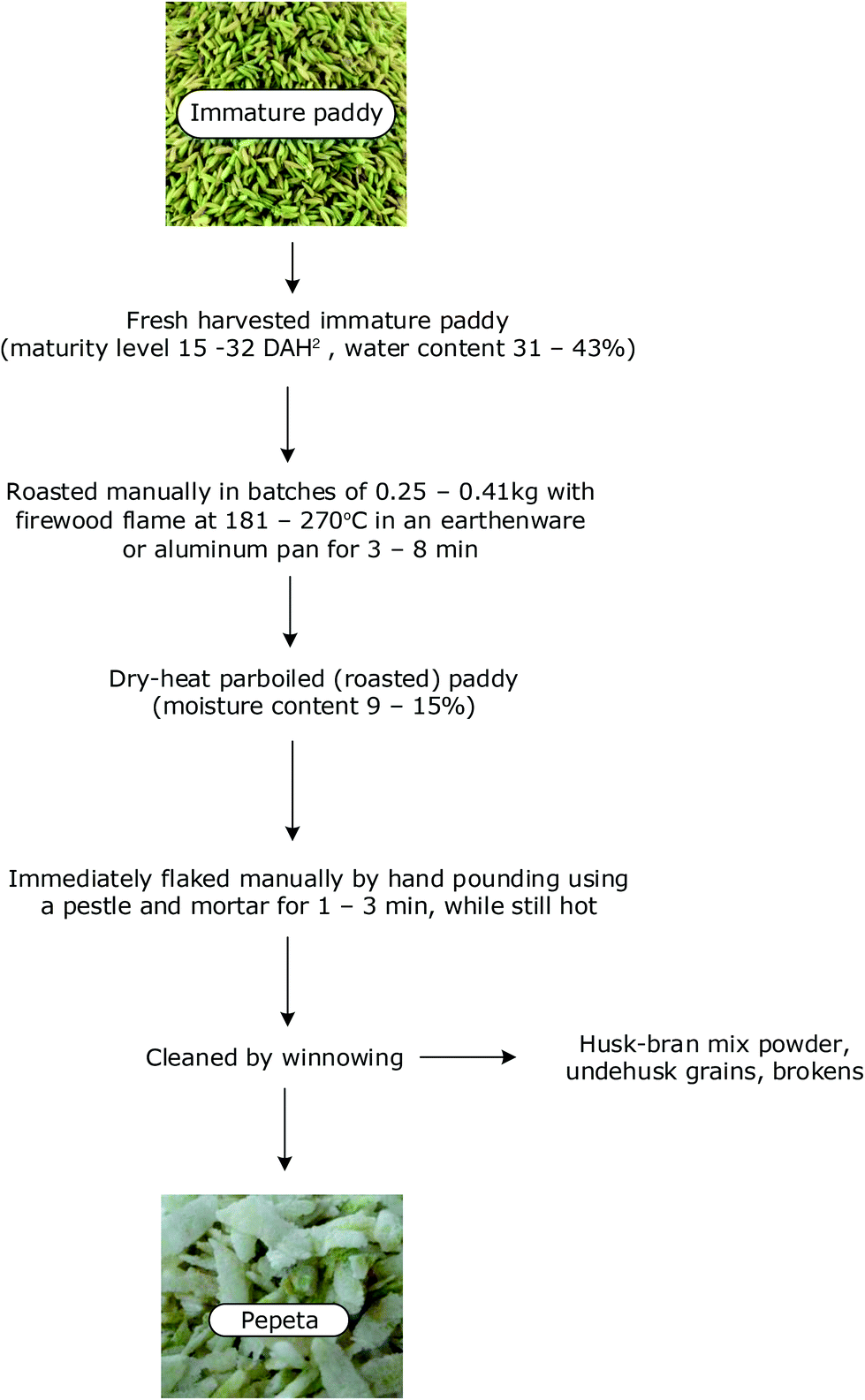 | ||
| Fig. 1 Pepeta processing flow chart. aDays after 50% heading/flowering. | ||
2. Materials and methods
2.1 Materials
![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 113, 129.3 U mg−1), trypsin from porcine pancreas (T7409, 1000–2000 U mg−1), α-chymotrypsin from bovine pancreas (C4129, ≥40 U mg−1), α-amylase from porcine pancreas (A4268, 700–1400 U mg−1), sodium dodecyl sulfate (SDS), o-phthaldialdehyde (OPA), DL-dithiothreitol (DTT), L-serine, thiamine hydrochloride (B1), nicotinic acid (B3, ≥99.5% HPLC), and nicotinamide (B3) were purchased from Sigma-Aldrich Ltd (St Louis, MO, USA). Trichloroacetic acid (CAS 76-03-9) and disodium tetraborate decahydrate (CAS 1303-96-4) were bought from Merck & Co. (Darmstadt, Germany). Assay kits for total starch, resistant starch, amylose/amylopectin, D-glucose (GOPOD) and dietary fibre analyses were acquired from Megazyme Inc. (Wicklow, Ireland). Other chemicals used in this study were of analytical grade.
113, 129.3 U mg−1), trypsin from porcine pancreas (T7409, 1000–2000 U mg−1), α-chymotrypsin from bovine pancreas (C4129, ≥40 U mg−1), α-amylase from porcine pancreas (A4268, 700–1400 U mg−1), sodium dodecyl sulfate (SDS), o-phthaldialdehyde (OPA), DL-dithiothreitol (DTT), L-serine, thiamine hydrochloride (B1), nicotinic acid (B3, ≥99.5% HPLC), and nicotinamide (B3) were purchased from Sigma-Aldrich Ltd (St Louis, MO, USA). Trichloroacetic acid (CAS 76-03-9) and disodium tetraborate decahydrate (CAS 1303-96-4) were bought from Merck & Co. (Darmstadt, Germany). Assay kits for total starch, resistant starch, amylose/amylopectin, D-glucose (GOPOD) and dietary fibre analyses were acquired from Megazyme Inc. (Wicklow, Ireland). Other chemicals used in this study were of analytical grade.
2.2 Sample preparation
:25) until fully gelatinized. Cooked rice kernels were pressed between two glass slides to check for full gelatinization, i.e. when no clear white core was observed anymore. Immediately after cooking, the samples were cryo-milled and stored for further analysis at −20 °C to avoid retrogradation. The remained broken grains after hand picking of head rice grains were stored at 4 °C and used as raw milled white rice samples.
2.3 Proximate analysis
A specific conversion factor (Jones’ factor for rice products) of 5.95 was used to convert the nitrogen content in the sample to protein.22
2.4 Crude dietary fibre
Soluble (SDF) and insoluble (IDF) dietary fibre were measured using a commercial Megazyme kit (K-TDFR, Megazyme Int, Wicklow, Ireland). In brief, 1 g of ground sample in MES-TRIS buffer solution was incubated with α-amylase, protease, and amyloglucosidase enzymes, in series. The mixture was then filtered to obtain the IDF, while SDF was determined by precipitating the filtrate with 95% of hot (60 °C) ethanol. Ash and protein residues were corrected for corresponding SDF and IDF values. Total dietary fibre (TDF) was calculated as the sum of SDF and IDF.2.5 Surface lipid content
Surface lipid content (SLC) was determined to quantify the degree of milling (DOM) according to Matsler and Siebenmorgen,23 following same procedure as in section 2.3.3. In this analysis, whole kernels of pepeta and milled rice samples were used.2.6 Analysis of Fe and Zn
Fe and Zn were measured by ICP-AES (inductively coupled plasma-atomic emission spectrometry) (Thermo iCAP-6500 DV; Thermo Fisher Scientific, Waltham, USA), following the validated Chemisch Biologisch Laboratorium Bodem (CBLB, Wageningen, the Netherlands) protocol. Briefly, 300 mg of ground sample was digested with concentrated nitric-hydrochloric acid mixture, and hydrogen-peroxide in a microwave digestion system (MarsXpress; CEM Corporation, Matthews, USA), consecutively. After settling of the undissolved silica particles, the supernatant was analysed on ICP-AES. Prior to analysis, samples were dried at 70 °C overnight and ground to 0.425 mm.2.7 Determination of selected vitamin B1 and B3
Determination of thiamine, nicotinic acid and nicotinamide was by a modified procedure of Chen et al.24 The B vitamins were extracted by adding 25 ml milli Q water to 0.5 g ground sample, sonicated at 40 °C for 5 h and then centrifuged at 3000g for 10 min at room temperature. The supernatant was filtered by a 0.2 μm membrane filter and 20 μL extract was used for HPLC analysis. The mobile phase was 25 mM KH2PO4 (pH3):CNCH3 in a ratio of 97:3 at a flow rate of 1.0 mL min−1. The HPLC system was operated with a Prevail C18 column (5 μm, 4.6 × 250 mm) and a UV detector set at 245 and 270 nm at room temperature.
2.8 Starch characterisation analysis
:3 (w/w) to ensure complete gelatinization of the samples. The samples were left to equilibrate for 5 h at room temperature after immediate hermetical sealing. Both samples and empty reference pans were then heated twice from 10 to 120 °C, with an heating rate of 10 °C min−1. Onset (To), peak (Tp) and conclusion (Tc) temperatures, and enthalpy change (ΔH) were analysed using Pyris™ software (Version 11, PerkinElmer, Inc. Waltham, USA).
2.9 In vitro digestion
In vitro digestion of starch and protein hydrolysis was performed based on the harmonized INFOGEST protocol27 with some modifications. The digestion procedure consisted of a gastric and an intestinal phase. Moistened pepeta (1:4 w/v) and cooked rice samples were mixed with pre-warmed simulated salivary fluid (SSF) without salivary α-amylase, simulated gastric fluids (SGF) and freshly prepared pepsin (2000 U ml−1), respectively. Prior to gastric digestion, the pH of the mixture was adjusted to 3 with 1 M HCl and incubated at 37 °C for 2 h. After the gastric phase, the gastric chyme was combined with warmed simulated intestinal fluids (ISF), fresh bile (28.8 mg mL−1), and a pancreatic enzyme solution consisting of α-amylase (200 U ml−1), trypsin (100 U ml−1) and chymotrypsin (25 U ml−1). The pH was adjusted to 7 with 1 M NaOH, after which the mixture was incubated at 37 °C for 2 h to complete intestinal digestion. Sample tubes were placed in a rotator (Multi Rs-60, Biosan, Riga, Latvia) set at 40 rpm throughout the whole digestion procedure. Separate sample tubes were used for aliquot sampling at 0, 60, 120 minutes during the gastric step and at 5, 10, 20, 30, 60 and 120 minutes of the intestinal step for starch and protein analysis, and immediately snap-frozen in liquid nitrogen for 20 s to minimize further enzymatic reactions. After the complete digestion procedure, amylase and protease activity in the aliquot samples (0.4 ml for each starch and protein) were stopped by addition of absolute ethanol (1:4) and 20% TCA (1:2), respectively. Subsequently, the aliquots sample mixtures were centrifuged at 3000g for 10 min at 0 °C, and the supernatant obtained stored at −20 °C until further analysis.
The effect of particle size in the rate and extent of starch and protein digestion was also evaluated. For this analysis, coarse particles from pepeta samples, the fractions retained between 2 and 1 mm sieves, were used.
| Ct = C∞(1 − e−kt) |
where: NH2 (DS), free amino groups from digested sample; NH2 (t = 0), free amino groups from samples at time 0 of digestion; NH2 (total), maximum amount of NH2 present in the sample.
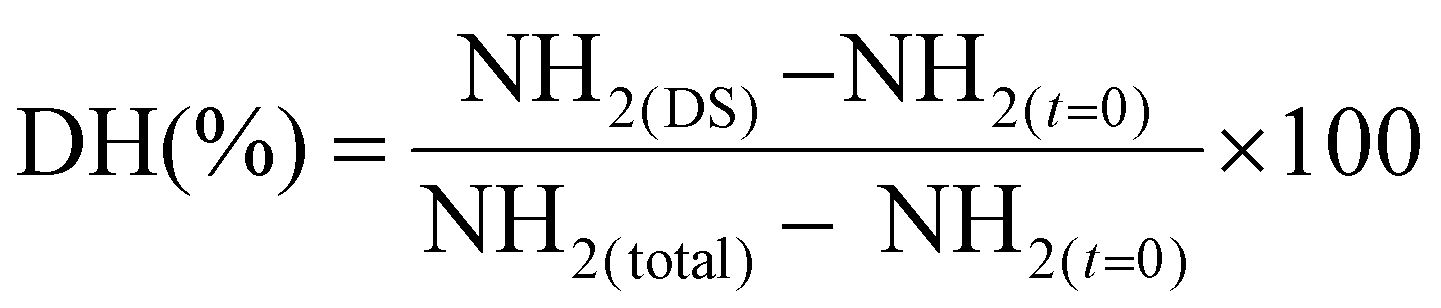
2.10 Confocal laser scanning microscopy
The endosperm cell wall morphology of non-digested raw rice, cooked rice and pepeta samples was visualized using a Zeiss 510 inverted microscope (Carl Zeiss microscopy, Oberkochen, Germany). For this, raw rice and pepeta samples were moistened between water-wetted tissue paper for 3 h. Cooked rice samples were freshly prepared in excess water (ratio of 1:25) to full gelatinization. All samples were manually sectioned into thin cross section slices, stained with 0.02% calcofluor-white dye and left to incubate for 5 min before being excited at 405 nm. Images were taken using 40× (N.A. 1.3 oil immersion) objective lenses.
2.11 Statistical analysis
One-way ANOVA analysis was performed with SPSS version 25 (IBM Statistics, Armon, USA) to evaluate the effects of maturity level for the individual varieties. When significant effects were observed, Tukey's test (p < 0.05) was used for multiple comparison. A paired-sample t test was used to assess the effect of pepeta processing method between rice and pepeta for each maturation level and variety, whereas independent t test was computed to evaluate statistical difference between pepeta samples in the same variety. Data are presented as means ± standard deviation of at least two replicates.3. Results and discussion
3.1 Proximate analysis
Table 1 summarizes the proximate composition of rice grains harvested at different grain development stages. The total lipid content of milled white rice grains ranged from 0.44 (FRS)–1.17 (MGS) g per 100 g in TXD306 and 0.80 (DGS)–0.99 (ORS) g per 100 g in Lawama, showing lower amounts than in previous research29 where paddy, i.e. whole rice grains, were used. In Lawama, the total lipid content increased as rice grains matured from DGS to ORS, while showing an inconsistent trend in TXD306. The observed total lipid trend could be due to variations in degree of milling (DOM) among maturity levels as a similar trend for surface lipid content (SLC) was observed in both varieties during rice maturation. In this study, SLC was used to measure the DOM, i.e. the amount of bran and germ remains after milling (section 2.5), as rice bran contains about 20% lipid.30 The protein content was 6.54 (DGS)–9.74 (MGS) g per 100 g in TXD306 and 6.19 (DGS)–8.33 (MGS) g per 100 g in Lawama, and showed significant differences among growth stages in both varieties. The results are within the range of values reported from fully mature rice.31 However, the protein content increased from DGS to MGS, decreased at FRS before increasing again at ORS for both varieties. This trend is in contrast to Chen-Ming et al.5 who reported a general decrease in protein content during rice grain development. A significant change in ash content was observed; immature grains showing higher amounts in TXD306 (1.03 g per 100 g, DGS) and Lawama (0.96 g per 100 g, DGS) than in fully matured grains in FRS (0.41 g per 100 g) and ORS (0.41 g per 100 g) respectively. Similar results have been reported for other rice varieties during their development.5| Nutritional compound | Sample | TXD306 | Lawama | ||||||
|---|---|---|---|---|---|---|---|---|---|
| DGS (15–21 DAH) | MGS (22–28 DAH) | FRS (29–35 DAH) | ORS (36–43 DAH) | DGS (15–21 DAH) | MGS (22–28 DAH) | FRS (29–35 DAH) | ORS (36–43 DAH) | ||
| Values in each row (small letter) and column (capital letter) bearing different superscripted letters are statistically different (p ≤ 0.05) for each rice variety. DGS – dough grain stage, MGS – mature grain stage, FRS – fully ripe stage, ORS – over ripe stage, DAH – days after 50% heading, db – dry basis, nd – not detected. Data expressed as mean ± standard deviation. | |||||||||
| Proximate | |||||||||
| Surface lipid content (g per 100 g, db) | Rice | 0.76 ± 0.09aB | 0.75 ± 0.07aA | 0.26 ± 0.03b | 0.68 ± 0.15a | 0.44 ± 0.06aA | 0.41 ± 0.03aB | 0.58 ± 0.06ab | 0.66 ± 0.01b |
| Pepeta | 1.66 ± 0.06aA | 1.32 ± 0.05bA | 1.10 ± 0.10aA | 1.37 ± 0.03aA | |||||
| Total lipids (g per 100 g, db) | Rice | 1.17 ± 0.02aB | 1.18 ± 0.04aB | 0.44 ± 0.02b | 0.85 ± 0.27ab | 0.80 ± 0.18aA | 0.89 ± 0.20aA | 0.92 ± 0.14a | 0.99 ± 0.03a |
| Pepeta | 2.14 ± 0.12aA | 1.55 ± 0.00bA | 1.68 ± 0.04aA | 1.65 ± 0.09aA | |||||
| Total protein (g per 100 g, db) | Rice | 6.54 ± 0.09aA | 9.74 ± 0.21bA | 6.63 ± 0.73a | 9.53 ± 0.14b | 6.19 ± 0.08aA | 8.33 ± 0.29bA | 6.30 ± 0.18a | 7.12 ± 0.02c |
| Pepeta | 7.33 ± 0.51bA | 9.66 ± 0.16aA | 6.58 ± 0.03aA | 8.36 ± 0.17bA | |||||
| Total ash (g per 100 g, db) | Rice | 1.03 ± 0.01aA | 0.76 ± 0.01bA | 0.41 ± 0.01c | 0.45 ± 0.01d | 0.96 ± 0.03aA | 0.59 ± 0.04bB | 0.96 ± 0.02a | 0.41 ± 0.01c |
| Pepeta | 1.10 ± 0.03aA | 0.86 ± 0.03bA | 1.03 ± 0.04aA | 1.09 ± 0.03aA | |||||
| Vitamins | |||||||||
| Thiamine (mg per 100 g, db) | Rice | 1.88 ± 0.17aA | 0.46 ± 0.01bB | 0.47 ± 0.05b | 0.43 ± 0.01b | 0.51 ± 0.00aA | 0.65 ± 0.00bA | 0.60 ± 0.03b | 0.56 ± 0.02a |
| Pepeta | 1.48 ± 0.03bA | 2.49 ± 0.03aA | 0.52 ± 0.02aA | 0.55 ± 0.06aA | |||||
| Nicotinic acid (mg per 100 g, db) | Rice | 49.7 ± 3.92aA | 45.2 ± 0.37aA | 16.1 ± 0.16b | 18.5 ± 3.47b | 34.2 ± 7.30abA | 46.04± 4.45aA | 28.06 ± 4.09ab | 21.3 ± 4.04b |
| Pepeta | 32.6 ± 4.81aA | 21.7 ± 0.43aB | 24.8 ± 0.48aA | 31.0 ± 8.51aA | |||||
| Nicotinamide (mg per 100 g, db) | Rice | 2.79 ± 0.09aA | 1.22 ± 0.06bB | 1.21 ± 0.00b | 1.48 ± 0.22b | 1.95 ± 0.17aA | 2.32 ± 0.75aA | 2.06 ± 0.37a | 1.67 ± 0.24a |
| Pepeta | 3.13 ± 0.35aA | 2.45 ± 0.19aA | 3.43 ± 0.85aA | 2.87 ± 0.61aA | |||||
| Mineral | |||||||||
| Iron (mg kg−1, db) | Rice | 0.59 ± 0.44aA | 0.64 ± 0.17aA | 0.31 ± 0.06a | 1.27 ± 0.37a | 0.58 ± 0.03bA | 2.05 ± 0.02aA | 0.89 ± 0.45b | 1.06 ± 0.23ab |
| Pepeta | 0.82 ± 0.14bA | 1.66 ± 0.05aA | 1.34 ± 0.44aA | 1.85 ± 0.17aA | |||||
| Zinc (mg kg−1, db) | Rice | 0.75 ± 0.00cB | 0.93 ± 0.01cB | 1.73 ± 0.07b | 2.11 ± 0.13a | 0.62 ± 0.05dB | 1.53 ± 0.08bA | 0.96 ± 0.05c | 2.14 ± 0.00a |
| Pepeta | 1.88 ± 0.00aA | 1.77 ± 0.01bA | 1.65 ± 0.05aA | 1.78 ± 0.01aA | |||||
Fig. 2 (panels a and b) reports the results of SDF, IDF and TDF. The amounts of TDF are in the same range as reported in previous work on rice maturation, and so is the ratio between SDF and IDF.9 The SDF content slightly increased during grain development in both varieties. However, the IDF showed an inconsistent trend in both varieties, whereas high values were observed at ORS and MGS for TXD306 and Lawama, respectively. The content of TDF, which is the sum of SDF and IDF, ranged from 2.90 (DGS)–5.04 (ORS) % in TXD306 and 2.64 (DGS)–3.19 (MGS) % in Lawama.
 | ||
| Fig. 2 Dietary fibre contents of two rice varieties (TXD306 and Lawama) as affected by maturity level (a and b) and effect of processing into pepeta (c–h). TDF – total dietary fibre, IDF – insoluble dietary fibre, SDF – soluble dietary fibre, rice – raw rice flour, Pepeta – locally prepared rice flake. Labels with different letters within maturity level indicate a statistically significant difference in fibre content (p ≤ 0.05). | ||
Pepeta products prepared from immature rice grains at DGS and MGS were also analysed to evaluate the effect of pepeta processing (Table 1 and Fig. 2 (panels c–h)). This study found more protein, TDF, IDF and SDF in pepeta compared to rice, though the values did not differ significantly. Pepeta processing significant increased total lipid content in TXD306 and total ash content in Lawama at MGS. These results are likely because of the bran residue.30 The results indicate that hand pounding and dehusking, i.e. an intermediate processing step in pepeta, is less efficient in removing bran and germ compared to mechanical milling. This caused pepeta to have significantly higher amount of SLC compared to milled rice. Further assessment of pepeta samples within variety showed significance differences between DGS and MGS for total protein, total lipid and total ash content in TXD306, and only total protein content in Lawama. The higher values were observed at DGS except for protein content in both variety, possibly due to associated trend with SLC.
3.2 Microstructural characterization
Confocal laser scanning microscopy (CLSM) was employed to visualize the morphology of endosperm cells of raw rice, cooked rice and pepeta in three different cell regions, i.e. the outer kernel layers, the central starchy endosperm and the crease region of the kernel sections (Fig. 3). Staining of the cell walls (light blue) in the micrographs of raw rice (Fig. 3 panels a–c), clearly shows different shapes of the cells in different regions of the endosperm. The endosperm cell wall profile elongates inwards when moving towards the inner region of the kernel sections, in line with previous findings.32,33 CLSM micrographs of pepeta (Fig. 3 panels d and e) resemble those of raw rice, indicating a structural organization of endosperm cell walls even after pepeta processing. However, the cell profile of the outer kernel layers in pepeta was partially disrupted as compared to those of raw rice, probably due to physical damage during pounding. Pepeta processing also involves roasting of paddy grain (dry heat), which apparently produced limited damage on cell wall structure compared with cooking (wet heat). A previous study34,35 reported that cell expansion and disruption in cooked rice depend on the extent of starch gelatinization, amount of water absorption and temperatures close to 100 °C. This is also the case in our study: cooked rice micrographs (Fig. 3 panels g–i), showed endosperm cell wall organization was lost after cooking with no visible intact cells. Note that rice grains were cooked in excess boiling water until fully gelatinized (section 2.2.2). | ||
| Fig. 3 Confocal images of raw rice (a–c), pepeta (d–f) and cooked rice (g–i) in different area of the cross section of the kernel; outer kernel layer (a, d and g), central starchy endosperm (b, e and h), and inner region of kernel (c, f and i). Cell walls were stained in light blue. Micrographs a–f were taken using 40× objective lens while g–i 20×. | ||
3.3 Vitamins and minerals
Table 1 shows the concentration of selected water-soluble B vitamins, i.e. thiamine, nicotinic acid and nicotinamide, measured in this study. Nicotinic acid (16.1 (FRS)–49.7 (DGS) mg per 100 g in TXD306 and 21.3 (ORS)–46.06 (MGS) mg per 100 g in Lawama) was abundant, followed by nicotinamide (1.21 (FRS)–2.79 (DGS) mg per 100 g in TXD306 and 1.67 (ORS)–2.32 (MGS) g per 100 g in Lawama). The amount of nicotinic acid and nicotinamide significant decreased during grain development; TXD306 at FRS and Lawama at ORS contained the lowers amounts. Significantly higher thiamine contents, 1.88 mg per 100 g for TXD306 and 0.65 mg per 100 g for Lawama were detected at DGS and MGS, respectively. Thiamine (except for TXD306 at DGS) and nicotinamide were in range with a study5 on two Korean rice varieties during maturation. Other studies5,36,37 reported decreases in vitamins B contents and/or their conjugates during maturation of cereal grains. The decrease could be due to the biochemical function of the B vitamins, which are cofactors and precursors in regulating plant metabolism.38–40Pepeta processing showed no significant effect on nicotinic acid and nicotinamide content (except at the MGS of TXD306) when comparing pepeta flour with rice flour (Table 1). In both varieties, pepeta contained higher levels of nicotinamide (2.45 (MGS)–3.13 (DGS) mg per 100 g in TXD306 and 2.87 (MGS)–3.43 (DGS) mg per 100 g in Lawama) than the corresponding rice grains. This trend is opposite for the nicotinic acid content. Though pepeta processing significantly affected thiamine content in TXD306 and Lawama, no consistent changes were observed. Except for thiamine in TXD306, no significant differences between DGS and MGS was observed on thiamine, nicotinic acid and nicotinamide in pepeta samples. The results indicate that maturity has very limited effect on analysed vitamin Bs composition in pepeta samples.
The amount of iron ranged from 0.31–1.27 mg per 100 g for TXD306 and 0.58–2.05 mg per 100 g for Lawama during rice maturation (Table 1), with the highest amounts at ORS for TXD306 and MGS for Lawama. The amount of zinc significantly increased as rice grains developed, with the highest levels at ORS of 2.11 and 2.14 mg per 100 g for TXD306 and Lawama, respectively. These results differ from previous work,5 possibly due to the use of different rice varieties and cultivation conditions.41,42
For pepeta processing, no significant difference was found for Lawama in the iron content between pepeta and the rice grains, although pepeta contained a higher amount than the grains. In TXD306, pepeta at MGS had a significantly higher iron content. Zinc content significant increased during pepeta processing at DGS (1.88 mg per 100 g) and MGS (1.77 mg per 100 g) in TXD306, and at DGS (1.65 mg per 100 g) in Lawama. These results are attributed to the high amount of bran and germ remains in pepeta (section 3.1). Maturity level significantly affect iron and zinc composition of pepeta in TXD306, high values found at MGS and DGS respectively. However, no significant difference was observed in Lawama, possibly due to observed concomitant SLC trend.
3.4 Starch characterisation
Total starch of rice grain ranged from 79.0 (ORS) to 82.8 (DGS) g per 100 g in TXD306 and from 80.9 (FRS) to 84.0 (DGS) g per 100 g in Lawama (Table 1), similar to previous research.43 Grain maturation did not significantly affect the total starch content in both varieties. Previous studies showed a constant level or small increase in starch content after 18 DAH during rice grain development.4,44 However, we observed a slight decrease of starch content during maturation, which may be associated with a concomitant slight increase in accumulation of protein observed during maturation (section 3.1). Reportedly, an increase in protein content in cereal grains negatively correlates with amylose accumulation, a component of starch.45,46 In addition, a rice proteomic study7,8 showed that considerably upregulated proteins were involved in starch synthesis and accumulation in developing grains. Though pepeta contained a slightly lower total starch content, no significant effect was observed between pepeta and its corresponding rice grains in both varieties. We attribute this to the high amount of bran residues in pepeta (section 3.1).Amylose content ranged from 17.1 (FRS) to 20.2 (ORS) g per 100 g in TXD306 and 15.2 (ORS)–20.1 (DGS) g per 100 g in Lawama, similar to previous work,47 which reported 1.6–21.7% amylose in polished rice samples of three different varieties. Except for TXD306 at ORS, the amylose content showed a decreased trend as rice grains matured, even though the differences were not significant. This is in line with previous study,48 suggested the accumulation of amylose in rice during the mid-stage of grain filling. Rice grains are classified according to amylose content.49 The varieties in this study are regarded as low amylose rice (10–19%).
RS contributes to gastrointestinal health as part of dietary fibre, reported to considerably reduce the postprandial blood glucose level.50 For a realistic comparison between the RS content of pepeta and rice, cooked rice grains were used as rice is normally eaten cooked,51 and the results were expressed per 100 g total starch (Table 2). The RS in cooked rice ranged from 0.28 to 0.54 g per 100 g in TXD306 and 0.05 to 0.40 g per 100 g in Lawama, the lowest values in both varieties recorded at FRS. The results corroborate previous research that reported less than 1% RS in cooked low-amylose rice varieties.52 Cooking (wet heat) and pepeta (dry heat) processing methods show no significant difference in RS content for both varieties. Dutta et al.16 and Sagum et al.53 also reported no significant difference in RS between boiling and pressure-cooking, and among dry heat parboiling methods in low amylose rice varieties, respectively. TXD306 and Lawama, like other low amylose varieties, both exhibit very low levels of RS when cooked and processed into pepeta.16,52,53
| Nutritional component | Sample | TXD306 | Lawama | ||||||
|---|---|---|---|---|---|---|---|---|---|
| DGS (15–21 DAH) | MGS (22–28 DAH) | FRS (29–35 DAH) | ORS (36–43 DAH) | DGS (15–21 DAH) | MGS (22–28 DAH) | FRS (29–35 DAH) | ORS (36–43 DAH) | ||
| a Expressed in g per 100 g of total starch, values in each row (small letter) and column (capital letter) bearing different superscripted letters are statistically different (p ≤ 0.05) for each rice variety. DGS – dough grain stage, MGS – mature grain stage, FRS – fully ripe stage, ORS – over ripe stage, DAH – days after 50% heading, db – dry basis, nd – not detected, D × 10 – average particle size of smallest 10%, D × 50 – average particle size of the smallest 50%, D × 90 – average particle size of the smallest 90%, C∞ – equilibrium hydrolysis, k – rate constant, To – onset temperature, Tp – peak temperature, Tc – conclusion temperature, ΔH – enthalpy change. Data expressed as mean ± standard deviation. | |||||||||
| Starch characterization | |||||||||
| Total starch (g per 100 g, db) | Rice | 82.8 ± 2.30aA | 81.4 ± 2.85aA | 81.2 ± 2.84a | 79.0 ± 0.14a | 84.0 ± 2.20aA | 81.5 ± 5.57aA | 80.9 ± 2.22a | 81.4 ± 2.51a |
| Pepeta | 78.2 ± 2.31aA | 77.1 ± 1.18aA | 78.7 ± 1.20aA | 78.2 ± 2.31aA | |||||
| Resistant starch (g per 100 g starch db)a | Rice | 0.54 ± 0.09aA | 0.41 ± 0.01abA | 0.28 ± 0.00b | 0.33 ± 0.05ab | 0.35 ± 0.05abA | 0.40 ± 0.13aA | 0.05 ± 0.01b | 0.24 ± 0.09ab |
| Pepeta | 0.35 ± 0.13aA | 0.34 ± 0.21aA | 0.33 ± 0.11aA | 0.21 ± 0.07aA | |||||
| Amylose (g per 100 g, db) | Rice | 19.2 ± 1.35a | 18.6 ± 3.16a | 17.1 ± 2.43a | 20.2 ± 0.03a | 20.1 ± 0.81a | 18.2 ± 2.74a | 19.1 ± 2.97a | 15.2 ± 2.03a |
| Particle size distribution | |||||||||
| Mean particle size (μm) | Rice | 5.35 ± 0.20a | 5.24 ± 0.29a | 5.54 ± 0.06a | 5.35 ± 0.07a | 5.23 ± 0.19a | 5.16 ± 0.05a | 5.21 ± 0.22a | 5.23 ± 0.04a |
| D × 10 (μm) | Rice | 3.68 ± 0.05a | 3.49 ± 0.09a | 3.73 ± 0.08a | 3.55 ± 0.02a | 3.62 ± 0.08a | 3.42 ± 0.14a | 3.55 ± 0.12a | 3.49 ± 0.09a |
| D × 50 (μm) | Rice | 5.17 ± 0.18a | 5.02 ± 0.18a | 5.34 ± 0.03a | 5.12 ± 0.05a | 5.05 ± 0.18a | 4.94 ± 0.08a | 5.03 ± 0.02a | 5.04 ± 0.05a |
| D × 90 (μm) | Rice | 7.21 ± 0.37a | 7.20 ± 0.75a | 7.56 ± 0.24a | 7.38 ± 0.18a | 7.02 ± 0.33a | 7.10 ± 0.06a | 7.06 ± 0.35a | 7.26 ± 0.01a |
| Starch hydrolysis | |||||||||
| C ∞ (g per 100 g, db)a | Cooked rice | 98.45 ± 2.35aA | 91.28 ± 8.15aA | 95.85 ± 1.39a | 99.07 ± 00a | 88.47 ± 1.90aA | 85.11 ± 1.92aA | 94.48 ± 5.11a | 93.80 ± 0.51a |
| Finely ground pepeta | 82.99 ± 2.13aB | 76.69 ± 3.97aA | 85.04 ± 2.30aB | 83.81 ± 6.79aA | |||||
| Coarsely ground pepeta | 81.81 ± 3.73aB | 79.32 ± 0.91aA | 79.78 ± 1.57aB | 76.05 ± 0.81aA | |||||
| k (min−1) | Cooked rice | 0.32 ± 0.02aA | 0.33 ± 0.06aA | 0.29 ± 0.00a | 0.25 ± 0.02a | 0.35 ± 0.02aA | 0.32 ± 0.00aA | 0.24 ± 0.02a | 0.26 ± 0.16a |
| Finely ground pepeta | 0.29 ± 0.02aA | 0.31 ± 0.02aA | 0.34 ± 0.05aA | 0.27 ± 0.00aB | |||||
| Coarsely ground pepeta | 0.20 ± 0.00aB | 0.18 ± 0.05aA | 0.25 ± 0.04aA | 0.20 ± 0.01aC | |||||
| Protein hydrolysis | |||||||||
| Gastric digestion (g per 100 g, db) | Cooked rice | 19.17 ± 6.16aA | 05.98 ± 0.98aA | 13.41 ± 5.53a | 09.42 ± 0.80a | 13.77 ± 4.74aA | 08.34 ± 2.36aB | 12.04 ± 7.60a | 14.75 ± 8.57a |
| Finely ground pepeta | 31.02 ± 8.85aA | 26.92 ± 18.4aA | 12.94 ± 2.67aA | 20.94 ± 0.69aA | |||||
| Coarsely ground pepeta | 08.60 ± 3.96aA | 08.74 ± 3.25aA | 21.07 ± 13.4aA | 11.86 ± 4.18aAB | |||||
| Intestinal digestion (g per 100 g, db) | Cooked rice | 71.66 ± 14.2aA | 46.87 ± 4.54aA | 68.92 ± 11.1a | 61.44 ± 0.94a | 73.10 ± 8.50aA | 54.36 ± 5.88aB | 70.47 ± 0.39a | 63.97 ± 19.3a |
| Finely ground pepeta | 84.06 ± 3.60aA | 74.50 ± 9.74aA | 79.47 ± 15.6aA | 94.48 ± 4.50aA | |||||
| Coarsely ground pepeta | 75.77 ± 8.66aA | 50.77 ± 16.1aA | 77.51 ± 7.32aA | 63.68 ± 3.74aB | |||||
| Gelatinization behaviour | |||||||||
| T o (°C), | Rice | 68.53 ± 0.37aA | 68.14 ± 0.25aA | 67.17 ± 0.18aA | 65.74 ± 0.17bA | ||||
| Rice starch | 64.90 ± 0.26aA | 63.99 ± 0.12bB | 64.07 ± 0.17aB | 61.99 ± 1.12aA | |||||
| Pepeta | nd | nd | nd | nd | |||||
| T p (°C) | Rice | 75.75 ± 0.18aA | 75.27 ± 0.22aA | 74.19 ± 0.03aA | 72.47 ± 0.17bA | ||||
| Rice starch | 71.32 ± 0.29aB | 70.71 ± 0.41aB | 70.32 ± 0.04aB | 67.95 ± 0.99aA | |||||
| Pepeta | nd | nd | nd | nd | |||||
| T c (°C) | Rice | 85.36 ± 0.30aA | 83.06 ± 0.18bA | 84.21 ± 0.01aA | 81.41 ± 0.22bA | ||||
| Rice starch | 80.05 ± 0.69aA | 78.93 ± 0.36aB | 79.26 ± 0.06aB | 75.57 ± 0.94bA | |||||
| Pepeta | nd | nd | nd | nd | |||||
| ΔH (J g−1) | Rice | 11.21 ± 1.15aA | 11.04 ± 0.26aA | 11.67 ± 0.15aA | 11.21 ± 0.06aB | ||||
| Rice starch | 14.80 ± 1.27aA | 15.82 ± 1.22aA | 15.33 ± 0.58aA | 15.12 ± 0.27aA | |||||
| Pepeta | nd | nd | nd | nd | |||||
In both varieties, particle size distribution analysis of rice starch granules showed no significant effect for mean particle size, D × 10, D × 50 and D × 90 as rice grains matured (Table 2), indicating that rice starch granules are fully developed at DGS. The mean particle size ranged from 5.24 (MGS) to 5.54 (FRS) μm in TXD306 and 5.16 (MGS) to 5.23 (ORS) μm in Lawama, which was similar as reported for six other rice varieties.54
DSC was conducted on raw rice flour of DGS and MGS, their corresponding starch and pepeta flour to investigate the effect of maturity, food matrix and pepeta processing on the gelatinization behaviour of starch (Table 2). Significant effects were found between DGS and MGS on Tp, Tp and Tp of rice flour and rice starch samples. However, there was no significant effect between maturity levels for the ΔH associated with gelatinization. The To, Tp, and Tc values shifted to lower temperatures in all samples (rice flour and rice starch) as rice grain developed from DGS to MGS. The results suggest possible differences in the way the polysaccharides are organized in the cell during grain development, which exhibit variation in thermal stability and crystallinity.55,56 In addition, we observed significant differences in To, Tp and Tc in both varieties, and ΔH of Lawama variety for rice flour as compared with rice starch samples. The To, Tp and Tc values in all varieties shifted towards lower temperatures in rice starch samples, likely due to absence of cell walls, limiting heat and water transfer to starch. The removal of proteins and lipids may also contribute to the shift as in Ye et al.57 who reported an increase in susceptibility of starch granules to gelatinization when lipids and/or protein were removed from the rice flour. In this study, no endothermic peak was observed in the DSC scans of pepeta flour from both varieties, indicating that pepeta processing conditions (initial moisture content of rice grains, and roasting temperature and duration) are sufficient to fully gelatinize pepeta starch, with very limited retrogradation.
3.5 In vitro digestibility of starch and proteins
 | ||
| Fig. 4 In vitro hydrolysis of starch in two rice varieties, TXD306 and Lawama during different stages of development (a and b) and as a function of processing method (c and d) and particles sizes (e and f). R-DGS – cooked rice at dough grain stage (15–21 days after 50% heading (DAH)), R-MGS – cooked rice at mature grain stage (22–28 DAH), R-FRS – cooked rice at fully ripe stage (29–35 DAH), R-ORS – cooked rice at over ripe stage (36–43 DAH), P-DGS – pepeta powder (≤0.425 mm) prepared at dough grain stage, P-MGS – pepeta powder (≤0.425 mm) prepared at mature grain stage, CP-DGS – pepeta coarse (2–1 mm) prepared at dough grain stage, an CP-MGS – pepeta coarse (2–1 mm) prepared at mature grain stage. | ||
According to Fig. 4 panels a and b, the ORS in TXD306 and MGS in Lawama had slightly lower digestibility under simulated conditions. The C∞ and k, estimated by fitting the experimental data to the first order kinetic equation (section 2.9), showed an inconsistent trend for maturation for both varieties (Table 2). The C∞ (91.28 (MGS)–99.07 (ORS) % in TXD306 and 85.11 (MGS)–94.48 (FRS) % in Lawama) and k (0.25 (ORS)–0.33 (MGS) min−1 in TXD306 and 0.24 (FRS)–0.32 (DGS) min−1 in Lawama) values for cooked rice are in line with previous work.31,57,61 However, no significant effect on C∞ and k values were observed in both varieties as rice grain developed from DGS to FRS. These results suggest that the susceptibility of cooked rice starch to in vitro digestion is not influenced by maturity level. The digestibility of rice starch by enzymatic hydrolysis has been extensively studied,16–18 but there appear to be relatively few reports concerning the susceptibility of immature rice starch as affected by maturation. It is well known that intrinsic characteristics such as granule size and shape, size and amount of amylose and amylopectin in the granules, molecular and supramolecular structure (crystallinity, growth rings, packing in cell), and amount of lipids and proteins and their interactions with starch granules may change during grain maturation, and are important factors affecting in vitro starch digestibility.56 However, none of these factors seems to play a role when starch is fully gelatinized (section 2.2.2) as in our study.
Digestograms (Fig. 4 panels c and d) clearly show the low digestibility of pepeta starch as compared to cooked rice in both varieties. No significant difference existed for estimated hydrolysis parameters (except C∞ at DGS in both varieties and k of Lawama variety at MGS) between cooked rice and pepeta. Slightly lower C∞ and k values were observed in pepeta as compared to cooked rice (Table 2). Chitra et al.17 reported lower rice starch digestion after a dry heat treatment as compared to a wet heat treatment (parboiling), suggesting RS formation in dry heat products to be the reason for low digestibility. However, in the present study, both cooked rice and pepeta samples contained similar, and very small amounts of RS, and starch was completely gelatinized (section 2.2.2 and 3.4). Therefore, the observed difference may be due to a more extensive disruption of cell walls by excess water during cooking (wet heat) compared to pepeta processing (dry heat) (section 3.2). It must be noted that the difference in starch digestibility between pepeta and cooked rice could even been higher should the cooked rice flour been passed through the 0.425 mm sieve. Starch digestibility in rice is known to be inversely proportional to the rice particle size.62,63
The effect of food bolus (particle size) on susceptibility of starch to in vitro enzymic hydrolysis was also evaluated in pepeta samples. Coarsely ground pepeta (CGP, 1–2 mm), which is an estimation of the swallowed particle size of pepeta,64 and fine ground pepeta (FGP, <0.425 mm) were compared for their in vitro starch digestibility. Fig. 4 (panels e and f) show that CGP had a slightly low digestibility than FGP under in vitro simulated digestion conditions, which was expected as digestion of grain flours is controlled by diffusion of enzymes through the milled grain fragments.65 Though no significant decrease in C∞ was observed in both varieties, k values significantly increased when the particle size of pepeta was increased, which indicates that the integrity of the rice matrix, and of the cell walls in particular (section 3.2), has an effect on pepeta starch digestibility, as repeatedly reported for other cereals.66–68 The slightly lower starch digestibility in pepeta is therefore possibly related to the higher level of structural integrity as compared to cooked rice.
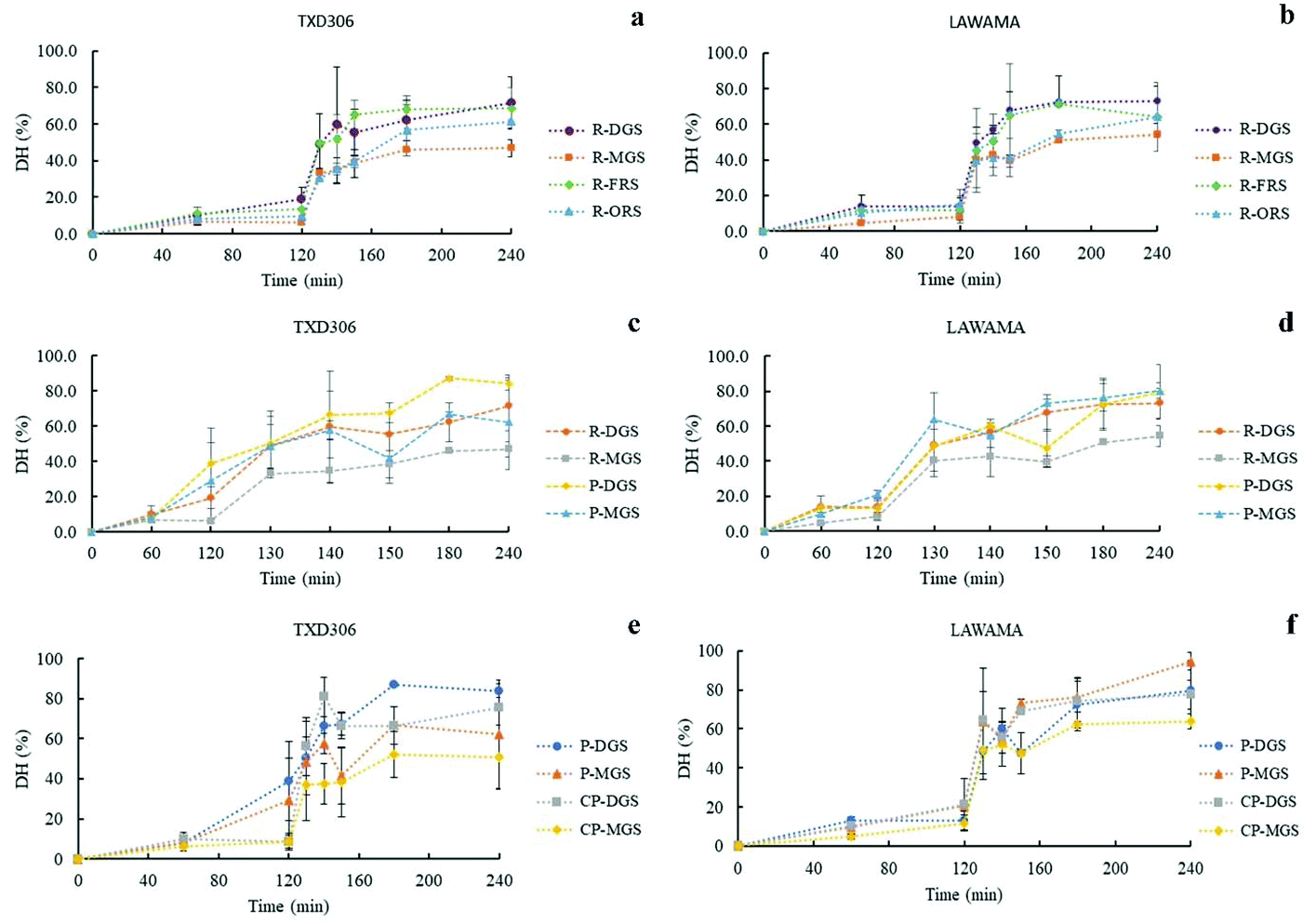 | ||
| Fig. 5 Digestograms of protein enzymatic hydrolysis in vitro for TXD306 and Lawama rice varieties during different stages of development (a and b), and as a function of processing method (c and d) and particles sizes (e and f). R-DGS – cooked rice at dough grain stage (15–21 days after 50% heading (DAH)), R-MGS – cooked rice at mature grain stage (22–28 DAH), R-FRS – cooked rice at fully ripe stage (29–35 DAH), R-ORS – cooked rice at over ripe stage (36–43 DAH), P-DGS – pepeta powder (≤0.425 mm) prepared at dough grain stage, P-MGS – pepeta powder (≤0.425 mm) prepared at mature grain stage, CP-DGS – pepeta coarse (2–1 mm) prepared at dough grain stage, an CP-MGS – pepeta coarse (2–1 mm) prepared at mature grain stage. | ||
The in vitro protein digestion also decreased with an increase in particle size in CGP (Fig. 5 panels e and f). As expected, low gastric and intestinal digestion values were observed in CGP compared to FGP due to likely hindrance of digestive enzymes by cell wall fragments in the larger particle sizes (section 3.2). The results are consistent with previous studies on the effect of particle size on in vitro digestibility of legume and cereal flours.28,65 However, differently from what observed with starch digestion, the presence of a higher degree of cellular integrity in CGP and FGP did not result in a reduced protein digestibility compared to cooked rice.
4. Conclusion
The present study investigated the changes in nutritional quality of rice grains during maturation. Immature rice represents an important source of micro- and macronutrients. This study demonstrates that a mild processing step such as the one used for pepeta can improve the nutritional potential of rice and its use as a food ingredient. The results showed a reduction in most nutritive components of rice grains during ripening. The highest levels of SLC, ash, thiamine, nicotinic acid and nicotinamide contents were observed at dough grain stage, whereas that of lipid, protein, SDF and IDF contents at mature stage. No effect of maturity on in vitro starch and protein digestibility was observed. On the other hand, conversion of rice into pepeta increased the content of some nutrients: pepeta contained up to twice as much iron and zinc, and three times more SDF compared to rice grains. Ash, lipid, nicotinamide, IDF and TDF contents in pepeta were increased by about 80%. The in vitro protein digestibility of pepeta was higher than for rice, indicating pepeta as a good source of high digestible protein. Pepeta starch susceptibility to hydrolytic enzymes in vitro was slightly lower than for cooked rice although the starch was completely gelatinized. Further investigations are necessary to understand the effects of processing conditions to optimize the nutritional quality of food products made from immature cereal grains, such as pepeta, and develop rice-based products with a more favourable glycaemic response than cooked rice.Conflicts of interest
The authors declare that they have no conflict of interest.Acknowledgements
We thank Jelle Sebastian Jabulani de Vries and Erik Meulenbroeks for helping performing part of the experimental work. We also grateful for the assistance given by Dr Ing. Norbert de Ruijter from Wageningen Light Microscopy Center for his support in confocal laser scanning microscopy analysis. This work was supported by the Netherlands Fellowship Programme (grant award number CF13182/2017).References
- P. A. Seck, A. Diagne, S. Mohanty and M. C. S. Wopereis, Crops that feed the world 7: Rice, Food Secur., 2012, 4(1), 7–24 CrossRef.
- H. Zhang, C. Yu, D. Hou, H. Liu, H. Zhang and R. Tao, et al., Changes in mineral elements and starch quality of grains during the improvement of japonica rice cultivars, J. Sci. Food Agric., 2018, 98(1), 122–133 CrossRef CAS.
- H. Li, M. A. Fitzgerald, S. Prakash, T. M. Nicholson and R. G. Gilbert, The molecular structural features controlling stickiness in cooked rice, a major palatability determinant, Sci. Rep., 2017, 7, 43713 CrossRef.
- R. Singh and B. O. Juliano, Free Sugars in Relation to Starch Accumulation in Developing Rice Grain, Plant Physiol., 1977, 59, 417–421 CrossRef CAS.
- J. Chen-Ming, J.-A. Shin, J.-W. Cho and K.-T. Lee, Nutritional evaluation of immature grains in two Korean rice cultivars during maturation, Food Sci. Biotechnol., 2013, 22(4), 903–908 CrossRef.
- O.-A. Olsen, Endosperm development: cellularization and cell fate specification, Annu. Rev. Plant Physiol. Plant Mol. Biol., 2001, 52, 233–267 CrossRef CAS.
- S. B. Xu, T. Li, Z. Y. Deng, K. Chong, Y. Xue and T. Wang, Dynamic Proteomic Analysis Reveals a Switch between Central Carbon Metabolism and Alcoholic Fermentation in Rice Filling Grains, Plant Physiol., 2008, 148(2), 908 CrossRef CAS.
- J. Lee and H. Koh, A label-free quantitative shotgun proteomics analysis of rice grain development, Proteome Sci., 2011, 9(61), 1–10 Search PubMed.
- P.-Y. Lin and H.-M. Lai, Bioactive compounds in rice during grain development, Food Chem., 2011, 127(1), 86–93 CrossRef CAS.
- S. Jiamyangyuen, N. Nuengchamnong and P. Ngamdee, Bioactivity and chemical components of Thai rice in five stages of grain development, J. Cereal Sci., 2017, 74, 136–144 CrossRef CAS.
- Ö. Özboy, B. Özkaya, H. Özkaya and H. Köksel, Effects of wheat maturation stage and cooking method on dietary fiber and phytic acid contents of firik, a wheat-based local food, Nahrung/Food, 2001, 45(5), 347–349 CrossRef.
- B. Ozkaya, S. Turksoy, H. Ozkaya, B. Baumgartner, I. Ozkeser and H. Koksel, Changes in the functional constituents and phytic acid contents of firiks produced from wheats at different maturation stages, Food Chem., 2018, 246, 150–155 CrossRef CAS.
- G.-X. Sun, P. N. Williams, Y.-G. Zhu, C. Deacon, A.-M. Carey and A. Raab, et al., Survey of arsenic and its speciation in rice products such as breakfast cereals, rice crackers and Japanese rice condiments, Environ. Int., 2009, 35(3), 473–475 CrossRef CAS.
- M. Tamura and Y. Ogawa, Visualization of the coated layer at the surface of rice grain cooked with varying amounts of cooking water, J. Cereal Sci., 2012, 56(2), 404–409 CrossRef.
- H. Li, Y. Wen, J. Wang and B. Sun, The molecular structures of leached starch during rice cooking are controlled by thermodynamic effects, rather than kinetic effects, Food Hydrocolloids, 2017, 73, 295–299 CrossRef CAS.
- H. Dutta, C. L. Mahanta and V. Singh, Changes in the properties of rice varieties with different amylose content on dry heat parboiling, J. Cereal Sci., 2015, 65, 227–235 CrossRef CAS.
- M. Chitra, V. Singh and S. Z. Ali, Effect of processing paddy on digestibility of rice starch by in vitro studies, J. Food Sci. Technol., 2010, 47(4), 414–419 CrossRef CAS.
- H. Dutta, C. L. Mahanta, V. Singh, B. B. Das and N. Rahman, Physical, physicochemical and nutritional characteristics of Bhoja chaul, a traditional ready-to-eat dry heat parboiled rice product processed by an improvised soaking technique, Food Chem., 2016, 191, 152–162 CrossRef CAS.
- K. V. Murthy, R. Ravi, K. K. Bhat and K. S. M. S. Raghavarao, Studies on roasting of wheat using fluidized bed roaster, J. Food Eng., 2008, 89(3), 336–342 CrossRef.
- S. Kumar and K. Prasad, Optimization of flaked rice dry roasting in common salt and studies on associated changes in chemical, nutritional, optical, physical, rheological and textural attributes, Asian – J. Chem., 2017, 29(6), 1380–1392 CrossRef CAS.
- AOAC, Official Methods for Analysis, AOAC International, Gaithersburg, MD, 2000, available from: http://hdl.handle.net/10637/3158 Search PubMed.
- L. Amagliani, J. O'Regan, A. L. Kelly and J. A. O'Mahony, Composition and protein profile analysis of rice protein ingredients, J. Food Compos. Anal., 2017, 59, 18–26 CrossRef CAS.
- A. L. Matsler and T. J. Siebenmorgen, Evaluation of operating conditions for surface lipid extraction from rice using a Soxtec system, Cereal Chem., 2005, 82(3), 282–286 CrossRef CAS.
- C. H. Chen, Y. H. Yang, C. T. Shen, S. M. Lai, C. M. J. Chang and C. J. Shieh, Recovery of vitamins B from supercritical carbon dioxide-defatted rice bran powder using ultrasound water extraction, J. Taiwan Inst. Chem. Eng., 2011, 42(1), 124–128 CrossRef CAS.
- Z. A. Syahariza, E. Li and J. Hasjim, Extraction and dissolution of starch from rice and sorghum grains for accurate structural analysis, Carbohydr. Polym., 2010, 82(1), 14–20 CrossRef CAS.
- J. A. Lock and G. Gouesbet, Generalized Lorenz–Mie theory and applications, J. Quant. Spectrosc. Radiat. Transfer., 2009, 110(11), 800–807 CrossRef CAS.
- M. Minekus, M. Alminger, P. Alvito, S. Ballance, T. Bohn and C. Bourlieu, et al., A standardised static in vitro digestion method suitable for food - an international consensus, Food Funct., 2014, 5(6), 1113–1124 RSC.
- M. Zahir, V. Fogliano and E. Capuano, Food matrix and processing modulate in vitro protein digestibility in soybeans, Food Funct., 2018, 9(12), 6327–6337 RSC.
- N. H. Kim, J. Kwak, J. Y. Baik, M. R. Yoon, J. S. Lee and S. W. Yoon, et al., Changes in lipid substances in rice during grain development, Phytochemistry, 2015, 116, 170–179 CrossRef CAS.
- N. T. W. Cooper and T. J. Siebenmorgen, Correcting head rice yield for surface lipid content (Degree of Milling) variation, Cereal Chem., 2007, 84(1), 88–91 CrossRef CAS.
- R. Azizi, E. Capuano, A. Nasirpour, N. Pellegrini, M.-T. Golmakani and S. M. H. Hosseini, et al., Varietal differences in the effect of rice ageing on starch digestion, Food Hydrocolloids, 2019, 95, 358–366 CrossRef CAS.
- E. Dornez, U. Holopainen, S. Cuyvers, K. Poutanen, J. A. Delcour and C. M. Courtin, et al., Study of grain cell wall structures by microscopic analysis with four different staining techniques, J. Cereal Sci., 2011, 54(3), 363–373 CrossRef CAS.
- L. Zhao, T. Pan, C. Cai, J. Wang and C. Wei, Application of whole sections of mature cereal seeds to visualize the morphology of endosperm cell and starch and the distribution of storage protein, J. Cereal Sci., 2016, 71, 19–27 CrossRef CAS.
- F. van de Velde, J. van Riel and R. H. Tromp, Visualisation of starch granule morphologies using confocal scanning laser microscopy (CSLM), J. Sci. Food Agric., 2002, 82(13), 1528–1536 CrossRef CAS.
- M. Tamura, T. Nagai, Y. Hidaka, T. Noda, M. Yokoe and Y. Ogawa, Changes in histological tissue structure and textural characteristics of rice grain during cooking process, Food Struct., 2014, 1(2), 164–170 CrossRef.
- M. Buchholz, A. M. Drotleff and W. Ternes, Thiamin (vitamin B1) and thiamin phosphate esters in five cereal grains during maturation, J. Cereal Sci., 2012, 56(1), 109–114 CrossRef CAS.
- N. Petrovska-Avramenko, D. Karklina and I. Gedrovica, Investigation of immature wheat grain chemical composition, Research for Rural Development - International Scientific Conference [Internet], 2016, 1, [102-5 pp.]. Available from: http://search.ebscohost.com/login.aspx?direct=true&db=asn&AN=120260192&site=ehost-live Search PubMed.
- A. Matsui, Y. Yin, K. Yamanaka, M. Iwasaki and H. Ashihara, Metabolic fate of nicotinamide in higher plants, Physiol. Plant., 2007, 131(2), 191–200 CAS.
- S. E. Bocobza, S. Malitsky, W. L. Araujo, A. Nunes-Nesi, S. Meir and M. Shapira, et al., Orchestration of thiamin biosynthesis and central metabolism by combined action of the thiamin pyrophosphate riboswitch and the circadian clock in Arabidopsis, Plant Cell, 2013, 25(1), 288–307 CrossRef CAS.
- J. A. Miret and S. Munné-bosch, Plant amino acid-derived vitamins: biosynthesis and function, Amino Acids, 2014, 46(4), 809–824 CrossRef CAS.
- C. Wd, Y. Hg, Z. Hm and T. Xg, Influences of Cadmium on Grain Mineral Nutrient Contents of Two Rice Genotypes Differing in Grain Cadmium Accumulation, Rice Sci., 2009, 16(2), 151–156 CrossRef.
- G. Chandel, S. Banerjee, S. See, R. Meena, D. J. Sharma and S. B. Verulkar, Effects of Different Nitrogen Fertilizer Levels and Native Soil Properties on Rice Grain Fe, Zn and Protein Contents, Rice Sci., 2010, 17(3), 213–227 CrossRef.
- P. S. Hu, H. J. Zhao, Z. Y. Duan, L. L. Zhang and D. X. Wu, Starch digestibility and the estimated glycemic score of different types of rice differing in amylose contents, J. Cereal Sci., 2004, 40(3), 231–237 CrossRef CAS.
- J. Chrastil, Changes of oryzenin and starch during preharvest maturation of rice grains, J. Agric. Food Chem., 1993, 41(12), 2242–2244 CrossRef CAS.
- W. Yu, X. Tan, W. Zou, Z. Hu, G. P. Fox and M. J. Gidley, et al., Relationships between protein content, starch molecular structure and grain size in barley, Carbohydr. Polym., 2017, 155, 271–279 CrossRef CAS.
- J. Bao, P. Xiao, M. Hiratsuka, M. Sun and T. Umemoto, Granule-bound SSIIa Protein Content and its Relationship with Amylopectin Structure and Gelatinization Temperature of Rice Starch, Starch/Staerke, 2009, 61(8), 431–437 CrossRef CAS.
- X. Yang, C. Chi, X. Liu, Y. Zhang, H. Zhang and H. Wang, Understanding the structural and digestion changes of starch in heat-moisture treated polished rice grains with varying amylose content, Int. J. Biol. Macromol., 2019, 139, 785–792 CrossRef CAS.
- A. Kumar, U. Sahoo, B. Baisakha, O. A. Okpani, U. Ngangkham and C. Parameswaran, et al., Resistant starch could be decisive in determining the glycemic index of rice cultivars, J. Cereal Sci., 2018, 79, 348–353 CrossRef CAS.
- C. J. Bergman, J. T. Delgado, A. M. McClung and R. G. Fjellstrom, An Improved Method for Using a Microsatellite in the Rice Waxy Gene to Determine Amylose Class, Cereal Chem., 2001, 78(3), 257–260 CrossRef CAS.
- T. A. Zafar, High amylose cornstarch preloads stabilized postprandial blood glucose but failed to reduce satiety or food intake in healthy women, Appetite, 2018, 131, 1–6 CrossRef.
- M. G. Sajilata, R. S. Singhal, R. Pushpa and P. R. Kulkarni, Resistant Starch - A Review, Compr. Rev. Food Sci. Food Saf., 2006, 5, 1–17 CrossRef CAS.
- Y. Ding, J. Huang, N. Zhang, S. K. Rasmussen, D. Wu and X. Shu, Physiochemical properties of rice with contrasting resistant starch content, J. Cereal Sci., 2019, 89 Search PubMed.
- R. Sagum and J. Arcot, Effect of domestic processing methods on the starch, non-starch polysaccharides and in vitro starch and protein digestibility of three varieties of rice with varying levels of amylose, Food Chem., 2000, 70(1), 107–111 CrossRef CAS.
- D. Yao, J. Wu, Q. Luo, J. Li, W. Zhuang and G. Xiao, et al., Influence of high natural field temperature during grain filling stage on the morphological structure and physicochemical properties of rice (Oryza sativa L.) starch, Food Chem., 2020, 310, 125817 CrossRef.
- H. Wang, Y. Liu, L. Chen, X. Li, J. Wang and F. Xie, Insights into the multi-scale structure and digestibility of heat-moisture treated rice starch, Food Chem., 2018, 242, 323–329 CrossRef CAS.
- M. R. Toutounji, A. Farahnaky, A. B. Santhakumar, P. Oli, V. M. Butardo and C. L. Blanchard, Intrinsic and extrinsic factors affecting rice starch digestibility, Trends Food Sci. Technol., 2019, 88, 10–22 CrossRef CAS.
- J. Ye, X. Hu, S. Luo, D. J. McClements, L. Liang and C. Liu, Effect of endogenous proteins and lipids on starch digestibility in rice flour, Food Res. Int., 2018, 106, 404–409 CrossRef CAS.
- J.-M. Fernandes, D. A. Madalena, A. C. Pinheiro and A. A. Vicente, Rice in vitro digestion: application of INFOGEST harmonized protocol for glycemic index determination and starch morphological study, J. Food Sci. Technol., 2020, 57, 1393–1404 CrossRef CAS.
- D. C. Whitcomb and M. E. Lowe, Human pancreatic digestive enzymes, Dig. Dis. Sci., 2007, 52(1), 1–17 CrossRef CAS.
- K.-C. Cheng, S.-H. Chen and A.-I. Yeh, Physicochemical properties and in vitro digestibility of rice after parboiling with heat moisture treatment, J. Cereal Sci., 2019, 85, 98–104 CrossRef CAS.
- R. Mujoo and S. Z. Ali, Susceptibility of Starch to in vitro Enzyme Hydrolysis in Rice, Rice Flakes and Intermediary Products, Lebensm.-Wiss. Technol., 1998, 31(2), 114–121 CrossRef CAS.
- A. M. Farooq, C. Li, S. Chen, X. Fu, B. Zhang and Q. Huang, Particle size affects structural and in vitro digestion properties of cooked rice flours, Int. J. Biol. Macromol., 2018, 118(Pt A), 160–167 CrossRef CAS.
- E. de la Hera, M. Gomez and C. M. Rosell, Particle size distribution of rice flour affecting the starch enzymatic hydrolysis and hydration properties, Carbohydr. Polym., 2013, 98(1), 421–427 CrossRef CAS.
- M. L. Jalabert-Malbos, A. Mishellany-Dutour, A. Woda and M. A. Peyron, Particle size distribution in the food bolus after mastication of natural foods, Food Qual. Prefer., 2007, 18(5), 803–812 CrossRef.
- P. Guo, J. Yu, S. Wang, S. Wang and L. Copeland, Effects of particle size and water content during cooking on the physicochemical properties and in vitro starch digestibility of milled durum wheat grains, Food Hydrocolloids, 2018, 77, 445–453 CrossRef CAS.
- G. J. S. Al-Rabadi, R. G. Gilbert and M. J. Gidley, Effect of particle size on kinetics of starch digestion in milled barley and sorghum grains by porcine alpha-amylase, J. Cereal Sci., 2009, 50(2), 198–204 CrossRef CAS.
- C. H. Edwards, M. M. Grundy, T. Grassby, D. Vasilopoulou, G. S. Frost and P. J. Butterworth, et al., Manipulation of starch bioaccessibility in wheat endosperm to regulate starch digestion, postprandial glycemia, insulinemia, and gut hormone responses: a randomized controlled trial in healthy ileostomy participants, Am. J. Clin. Nutr., 2015, 102(4), 791–800 CrossRef CAS.
- M. Tamura, J. Singh, L. Kaur and Y. Ogawa, Impact of structural characteristics on starch digestibility of cooked rice, Food Chem., 2016, 191, 91–97 CrossRef CAS.
- K. Liu, J. Zheng and F. Chen, Effect of domestic cooking on rice protein digestibility, Food Sci. Nutr., 2019, 7(2), 608–616 CrossRef CAS.
- R. Mujoo, A. Chandrashekar and S. Z. Ali, Rice Protein Aggregation During the Flaking Process, J. Cereal Sci., 1998, 28, 187–195 CrossRef CAS.
- W. Zhang, S. Liu, Y. Wang, L. Zheng, F. Liu and X. Han, et al., A study of the in vitro protein digestibility of indica and japonica cultivars, Food Chem., 2010, 122(4), 1199–1204 CrossRef CAS.
| This journal is © The Royal Society of Chemistry 2020 |