
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Wheat-derived arabinoxylans reduced M2-macrophage functional activity, but enhanced monocyte-recruitment capacity†
Coen
Govers‡
 *a,
Yongfu
Tang‡
abc,
Ellen H.
Stolte
d,
Harry J.
Wichers
ab and
Jurriaan J.
Mes
a
*a,
Yongfu
Tang‡
abc,
Ellen H.
Stolte
d,
Harry J.
Wichers
ab and
Jurriaan J.
Mes
a
aWageningen - Food & Biobased Research, Wageningen University & Research, Wageningen, The Netherlands. E-mail: coen.govers@wur.nl
bLaboratory of Food Chemistry, Wageningen University & Research, Wageningen, The Netherlands
cAgricultural Products Processing Research Institute, Chinese Academy of Tropical Agricultural Sciences, Zhanjiang, China
dHost-Microbe Interactomics Group, Animal Sciences Department, Wageningen University & Research, Wageningen, The Netherlands
First published on 22nd July 2020
Abstract
The immunomodulatory properties of non-digestible polysaccharides (NDPs) have been recognized in in vitro and in vivo studies. The latter mostly demonstrated altered frequencies and inflammatory status of immune cells as clinical parameters. Most of the NDP activity will be exerted in the intestine where they can directly interact with macrophages. The predominant macrophage phenotype in the intestine is M2-like, with M1-like macrophages arising during inflammation. Here, we investigated transcriptional and functional impact on these macrophage phenotypes by NDP-treatment (i.e. yeast-derived soluble β-glucan (yeast-βG), apple-derived RG-I (apple-RGI), shiitake-derived β-glucan (shiitake-βG) or wheat-derived arabinoxylan (wheat-AX)). Wheat-AX, and to a lesser extent shiitake-βG and apple-RGI but not yeast-βG, reduced endocytosis and antigen processing capacity of M1- and M2-like macrophages. Moreover, the NDPs, and most notably wheat-AX, strongly induced transcription and secretion of a unique set of cytokines and chemokines. Conditioned medium from wheat-AX-treated M2-like macrophages subsequently demonstrated strongly increased monocyte recruitment capacity. These findings are in line with clinically observed immunomodulatory aspects of NDPs making it tempting to speculate that clinical activity of some NDPs is mediated through enhanced chemoattraction and modifying activity of intestinal immune cells.
Introduction
Non-digestible polysaccharides (NDPs) constitute a large family of different molecules, including β-glucans, pectins, resistant starch, cellulose and arabinoxylans, and are known for their immune-modulatory properties and beneficial health effects.1,2 These properties were demonstrated in vitro as well as reported in clinical studies using different NDPs. For example, yeast β-glucan intake by obese people lowered pro-inflammatory cytokine levels (i.e. IL-6 and TNF-α) and increased anti-inflammatory IL-10 levels in peripheral blood, accompanied by lowering risk factors for wellbeing such as waist circumference and blood pressure.3 Also, rice bran arabinoxylan increased NK cell activity and peripheral blood levels of myeloid-derived dendritic cells and TH1 related cytokines in multiple myeloma patients, unfortunately without reported beneficial effects on disease progress.4 A randomized dietary intervention in healthy young adults demonstrated that consuming shiitake mushrooms containing lentinan reduced CRP levels and therefore lowered peripheral inflammation but also increased immune cell effector potency.5 This coincided with an altered cytokine balance of increased IL-4, IL-10, TNF-α and IL-1α levels and decreased MIP1α/CCL3 level. Other beneficial health effects related to NDP intake reported in clinical trials include increased NK cell activity linked to a reduction in metastasis in lung and breast cancer patients;6 increased blood leukocytes, neutrophils, IgG and IgM frequencies accompanied by delayed progression of non-small cell lung cancer;7 and transiently increased blood levels of IgG and IgM and NK cell counts linked to a reduction in flu and flu-like symptoms and respiratory tract infections in children.8The large variation in NDP functionality and structure resulting from source, extraction method, saccharide composition and/or branching hampers identification of the mechanism of action.9 However, many studies have demonstrated that NDPs are recognized by immune cells through binding to pattern recognition receptors (PRRs). Dectin-1 was the first receptor shown to bind β-glucans to initiate functional responses in macrophages.10 Increasingly, PRRs are being identified through which NDPs can modulate immune responses including dectin-2,11 mannose receptor,12 complement receptor 3![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 13 and scavenger receptors.14
13 and scavenger receptors.14
Orally consumed NDPs will first and foremost exert their immunomodulatory activity in the intestine. NDPs can interact with a variety of intestinal immune cells located in the Peyer's patches through microfold cell-mediated transcytosis across the intestinal epithelium.15 Also, NDPs can directly be sampled in the lumen by CX3CR1+ antigen presenting cells which were shown to consist for 75–80% of macrophages16 in mice. Furthermore, studies into macrophage populations in human mucosa of the duodenal-proximal jejunum revealed different subsets, but all positive for the CX3CR1 fractalkine receptor.17 These CX3CR1+ macrophages were demonstrated to be of an immune suppressive subset (often referred to as M2).18 In case of intestinal immune activation, a CCR2-dependent influx of circulating CD14+ monocytes is initiated which under local conditions polarize into an inflammatory macrophage subset (often referred to as M1).19,20 The immune suppressive and inflammatory macrophage balance is key for maintaining intestinal homeostasis.21,22 Furthermore, intestinal macrophages have been shown to endocytose and degrade β-glucans.23 Beta-glucan fragments were subsequently transported towards the bone marrow, spleen and peripheral lymph nodes and locally released to exert immunomodulatory effects. We therefore pose that in vitro models based on macrophages are relevant to understand the immunomodulatory potential of NDPs and study the mechanism of action. Given the central position of macrophages in intestinal, and potentially peripheral, immune homeostasis we used an M1/M2 dichotomy in vitro model system to further understand the functional impact of NDP intake.
Materials and methods
NDP characteristics
The NDPs and their treatment to remove lipopolysaccharide were described previously.24 Briefly, a Saccharomyces cerevisiae (yeast)-derived soluble 94% pure β-1,3/1,6-glucan preparation (yeast-βG) was provided by BioThera (Eagan, MN, USA); a Lentinula edodus (shiitake)-derived 73% pure β-1,3/1,6-glucan (shiitake-βG) was produced in our lab; an apple-derived 80% pure rhamno-galacturonan-I (apple-RGI) preparation was provided by INRA (Paris, France); and a wheat-derived 77% pure arabinoxylan (wheat-AX) was provided by BioActor (Maastricht, The Netherlands). The NDPs, except yeast-βG, were treated with alkaline-ethanol for 5 hours, neutralized with HCl and lyophilized, resulting in lipopolysaccharide levels below immune stimulatory thresholds.25Primary macrophage differentiation and polarization
Primary macrophages were differentiated as previously described.24 In brief, human monocytes were obtained from buffy coats from healthy donors (Sanquin, Nijmegen, The Netherlands) using the QuadroMACS system and CD14 microbeads according to the manufacturer's protocol (Miltenyi Biotec, Leiden, The Netherlands). A written informed consent was obtained before sample collection. Monocytes were differentiated into macrophages through a 7 day culture in RPMI 1640 – glutamax medium (Gibco, Bleiswijk, The Netherlands) supplemented with 1% of pen/strep (Sigma, St Louis, MO, USA), 1% of sodium-pyruvate (Lonza, Breda, The Netherlands), 1% of MEM non-essential amino acids (Gibco), 10% of fetal bovine serum (FBS, Hyclone, Eindhoven, The Netherlands) and 50 ng ml−1 MCSF (R&D systems, Minneapolis, MN, USA) at 1 × 106 cells per 2 ml per well in a 24-well culture plate. After three and five days of culture half of the medium was replaced with fresh culture medium containing 100 ng ml−1 MCSF (providing a final concentration of 50 ng ml−1). After 7 days of culture all of the medium was replaced with culture medium either without additions to generate M0, with 20 ng ml−1 IFN-γ (R&D systems) and 20 ng ml−1 TNF-α (R&D systems) to generate M1, with 20 ng ml−1 IL-4 (R&D systems) to generate M2 or with 500 μg ml−1 of NDP preparation and incubated for 18 hours. Validation of M0, M1 and M2 macrophage phenotype was performed via Q-PCR and based on gene transcription markers as established previously.24 Regarding M1 and M2 macrophages, when applicable the culture medium was again completely replaced with either culture medium without additions or 500 μg ml−1 of NDP preparations.RNA extraction, cDNA synthesis and qPCR
RNA extraction, cDNA synthesis and qPCR was performed as described previously.24 In brief, RNA was extracted with TRIzol (Invitrogen, Bleiswijk, The Netherlands) and RNeasy kits (Qiagen, Venlo, The Netherlands) and quality and quantity controlled. Subsequently, cDNA was synthesized with iScript (Bio-Rad, Veenendaal, The Netherlands). Next, 5 μl cDNA was mixed with SYBRGreen supermix (Bio-Rad), forward and reverse primers, and milliQ water to total 20 μl. Primers were derived from the Harvard Primerbank (http://pga.mgh.harvard.edu/primerbank/) and synthesized by Biolegio (Nijmegen, The Netherlands). Primers’ information is available upon request. Amplification was performed with the CFX96 Touch Real-Time PCR Detection System (Bio-Rad) in technical duplicate and all samples were normalized to reference genes and medium control stimulations using the qBase+ software (Biogazelle, Gent, Belgium).Endocytosis assay and antigen processing analysis
Macrophages that were polarized, NDP- or medium-treated in a 24-well plate were washed with PBS and incubated for 1 h in 0.5 ml culture medium containing 2 μg ml−1 FITC-labelled E. coli fragments (excitation at 495 nm, emission at 519 nm; Molecular Probes, Leiden, The Netherlands) to analyze for phagocytic activity or 10 ng ml−1 DQ-OVA (D12053, Molecular Probes) to analyze antigen processing. OVA is heavily labelled with fluorescent green BDY dyes (excitation at 502 nm, emission at 510 nm) resulting in a quenching effect. Protease mediated hydrolysis of OVA, as expected upon processing, reliefs the quenching effect. Before addition, E. coli fragments were re-suspended in non-heat-inactivated human serum (2 mg per 100 μl) and incubated for 60 minutes at 37 °C. Following incubation with either E. coli or DQ-OVA, macrophages were washed with PBS without calcium and magnesium and detached following 10 minutes incubation with 0.25% Trypsin/EDTA (Gibco, Bleiswijk, The Netherlands) at 37 °C. Subsequently, cells were at least 4 times diluted in PBS with 2% FBS and centrifugation at 300g for 10 minutes. Cells were re-suspended in 200 μl PBS, 5 μl trypan blue was added to cells incubated with E. coli fragments for quenching extracellular fluorophores, and analyzed using flow cytometry (Accuri™ C6, BD Biosciences) with Accuri C6 software. For each individual experiment MFI values were collected of marker positive cells (marker set at 1% on non-stained cells) after which medium stimulated MFI values were set at 100% to correct for donor differences.Multiplex cytokine and chemokine analysis
Secretion of cytokines and chemokines (i.e. CCL1, CCL5, CCL20, CCL24, CXCL8, IL-1β, TNFα, MMP1, CXCL9, CXCL10 and CXCL11) by macrophages was measured using Bio-Plex Pro Reagent Kit (BioRad, Veenendaal, The Netherlands) according to the manufacturer's instructions. In brief, 50 μL magnetic beads were dispersed in a plate together with 50 μl of undiluted sample or standard and incubated for 1 h at RT. Subsequently, the beads were incubated with 25 μl detection antibodies for 30 min and with 50 μl streptavidin-PE for 10 min at RT. The beads were re-suspended in 125 μl assay buffer and read by Bio-Plex® MAGPIX™ Multiplex Reader (BioRad). Data processing was performed using Bio-Plex Manager 5.0, and concentrations (in pg ml−1) were interpolated from standard curves. To identify relations and to display our results most effectively, we used lower or upper limit of the standard curve for data points that were below or above the standard curve, respectively.Cell migration assay
Cell migration assays were performed using an xCELLigence RTCA DP system (xCELLigence, ACEA Biosciences, San Diego, USA) and CIM-16 well plates, according to the manufacturer's instructions. In brief, 160 μl of macrophage-conditioned medium were added to the lower chamber of CIM-16 well plates. Fresh macrophage culture medium was used as negative control and CCL5 (400 ng ml−1) as positive control. Following placement of the upper chamber the upper chamber wells were filled with 50 μl THP-1 cell culture medium and equilibrated at 37 °C for 1 h which was recorded as background cell index. Next, 50 μl THP-1 cell suspension (8 × 106 cells per ml) was added to the upper chamber after which cells settled for 5 minutes at RT. The cell index is a unit-less parameter used to indicate the impedance of electron flow caused by adherent cells and defined as (impedance at time point impedance n − impedance in the absence of cells)/nominal impedance value. The cell index was measured every 5 min over the course of 22 h. Monocyte migration was subsequently calculated as area under the curve using Prism 6 software (Graphpad, La Jolla, USA).Statistics
Statistical analysis and graphing were performed using GraphPad Prism (version 6, Graphpad Software). The data are presented as means ± SD and differences were assessed by one- or two-way ANOVA, followed by multiple comparisons test with the Fisher's LSD test. Differences with p < 0.05 were considered significant.Results
Functional characterization of non-polarized macrophages following exposure to NDPs
Previously, we performed a transcriptional analysis of non-polarized macrophages following their exposure to NDPs. Functionality of these NDP-treated macrophages was assessed here by comparing endocytosis capacity, antigen processing and cytokine and chemokine production to that of M1- or M2-like macrophages. To this end, non-polarized macrophages were treated with yeast-βG (i.e. soluble), shiitake-βG, apple-RGI or wheat-AX or polarized towards M1- or M2-like macrophages.Macrophage endocytosis capacity following NDP-treatment of polarization was investigated by incubation with fluorescent E. coli fragments and detection using flow cytometry (Fig. 1A and ESI Fig. 1A†). M2-like macrophages demonstrated a trend towards increased E. coli uptake whereas M1-like macrophages significantly reduced E. coli uptake. Similarly, macrophages treated with shiitake-βG or wheat-AX revealed a significant reduced E. coli uptake. Next, to investigate the potential for antigen processing, the NDP-treated or polarized macrophages were incubated with DQ-OVA. Polarization towards M1 or M2 phenotypes did not alter DQ-OVA processing by macrophages (Fig. 1B and ESI Fig. 1B†). In contrast, processing of DQ-OVA was significantly reduced by apple-RGI, shiitake-βG and wheat-AX. Finally, the secretion of chemokines and cytokines by NDP-treated or polarized macrophages was assessed. Tested signaling molecules were selected based on gene transcription levels of M1-like macrophages, with typically and/or significantly increased transcription of CXCL9, CXCL10, CXCL11, and TNFα, M2-like macrophages, with a strongest and significant increase in CCL24 transcription, and NDP-treated macrophages, with typically and/or significantly increased CCL1, CCL5, CCL20, CXCL8, IL-1β, and MMP-1 transcription for most NDPs (ESI Table 1†). Polarization of macrophages into an M1-like phenotype resulted in significantly increased secretion of TNFα, CXCL9, CXCL10, and CXCL11, whereas macrophage polarization into an M2-phenotype did not significantly affect secretion of tested cytokines or chemokines (Table 1). Exposure of macrophages to NDPs, and in particular wheat-AX, affected secretion of signaling molecules. Wheat-AX significantly increased the secretion of CCL1, CCL5, CCL20, CXCL8 and IL1β. These signaling molecules are described to induce migration of monocytes (ESI Table 1†). Therefore, a monocyte migration assay was performed using cell culture medium as negative control, CCL5 as positive control and conditioned medium from non-polarized macrophages, macrophages polarized towards an M1-like phenotype and wheat-AX-treated macrophages. In line with the cytokine and chemokine secretion profiles, conditioned medium from M1-like polarized macrophages or wheat-AX-treated macrophages significantly increased monocyte migration when compared to medium from non-polarized macrophages (Fig. 1C).
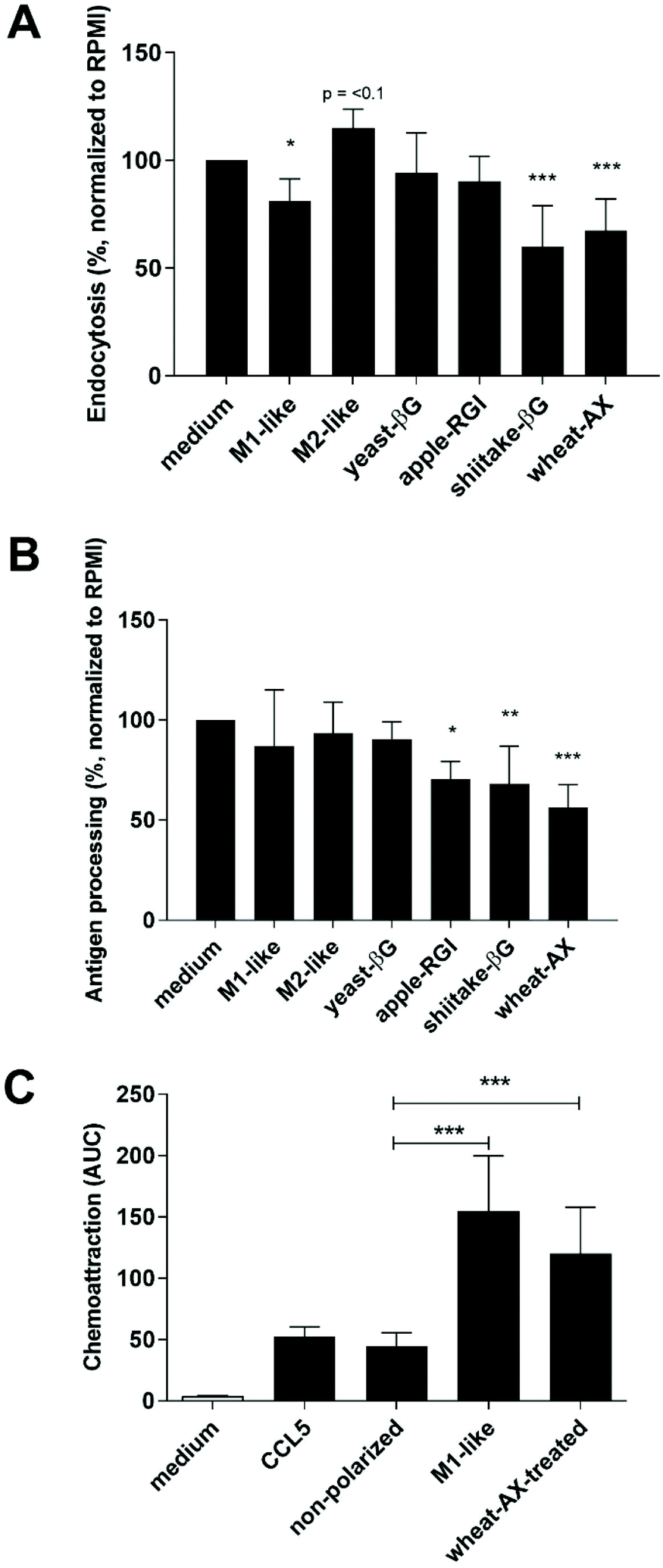 | ||
| Fig. 1 NDP-treatment induces different functionality in non-polarized macrophages compared to polarization towards M1- or M2-like macrophages. Macrophages were non-treated (medium), polarized towards M1-like or M2-like macrophages or treated with 500 μg ml−1 yeast-βG, apple-RGI, shiitake-βG or wheat-AX for 18 hours and subsequently incubated for 1 h with fluorescent E. coli-fragments (A) or DQ-OVA (B) and uptake measured with flow cytometry as MFI. Cell culture medium, CCL5 or conditioned medium from medium- or wheat-AX-treated macrophages or M1-like polarized macrophages was analyzed for the potency to induce monocyte migration (C) using the xCeLLigence RTCA DP system and THP-1 monocytes. Bars represent mean values (normalized to medium stimulation (100%) for A and B) ± SD of n = 2–5 independent donors. Statistically significant differences were analyzed compared to medium control by one-way ANOVA: * p < 0.05, ** p < 0.01, *** p < 0.001. | ||
| Proteins | Medium | M1 | M2 | Yeast-βG | Apple-RGI | Shiitake-βG | Wheat-AX |
|---|---|---|---|---|---|---|---|
| Macrophages were non-treated (medium), polarized towards M1-like or M2-like macrophages or treated with 500 μg ml−1 yeast-βG, apple-RGI, shiitake-βG or wheat-AX for 18 h. Cytokine and chemokine secretion (pg ml−1) was analyzed with Bio-Plex multiplex analysis. Values represent mean values ± SD of n = 3 different donors. Statistically significant differences were analyzed compared to medium by one-way ANOVA: * p < 0.05. | |||||||
| CCL1 | 22 ± 8 | 394 ± 365 | 8.3 ± 0 | 82 ± 104 | 322 ± 297 | 3295 ± 2463* | 7088 ± 3535* |
| CCL5 | 23 ± 14 | 1635 ± 289 | 22 ± 13 | 32 ± 18 | 441 ± 194 | 207 ± 53 | 2744 ± 2650* |
| CCL20 | 5.9 ± 8 | 174 ± 173 | 1.2 ± 1 | 5.6 ± 4 | 60 ± 16 | 7.4 ± 3 | 3504 ± 2672* |
| CCL24 | 64 ± 39 | 36 ± 4 | 97 ± 46 | 119 ± 129 | 37 ± 21 | 1321 ± 1041* | 775 ± 586 |
| CXCL8 | 2280 ± 1653 | 2764 ± 622 | 688 ± 353 | 5457 ± 1229 | 16581 ± 1840* |
18956 ± 1785* |
19008 ± 1593* |
| IL-1β | 0.1 ± 0 | 0.3 ± 0 | 0.1 ± 0 | 0.4 ± 0 | 2.3 ± 2 | 0.7 ± 1 | 195 ± 251* |
| TNFα | 8.1 ± 3 | 31395 ± 16953* |
4.0 ± 1 | 13 ± 6 | 548 ± 477 | 387 ± 366 | 13215 ± 14182 |
| MMP1 | 123 ± 0 | 166 ± 37 | 123 ± 0 | 144 ± 37 | 187 ± 65 | 166 ± 37 | 797 ± 909 |
| CXCL9 | 44 ± 44 | >24199* |
200 ± 227 | 47 ± 69 | 53 ± 12 | 371 ± 314 | 92 ± 102 |
| CXCL10 | 136 ± 103 | 27617 ± 17822* |
60 ± 31 | 93 ± 30 | 25 ± 29 | 55 ± 19 | 45 ± 39 |
| CXCL11 | 0.5 ± 0 | 5830 ± 3927* | 0.3 ± 0 | 0.4 ± 0 | 0.1 ± 0 | 0.4 ± 0 | 0.5 ± 1 |
Apple-RGI, shiitake-βG and wheat-AX induced similar transcriptional profiles in M1- and M2-like macrophages
In a steady state most of the macrophages in the intestine will have an M2-like phenotype.18 Therefore, M2-like macrophages were exposed to medium or NDPs and analyzed for their transcriptional expression of in part previously established gene markers.24 Yeast-βG only demonstrated to significantly increase transcription of CXCL8 and MINCLE (Fig. 2). Apple-RGI, shiitake-βG and wheat-AX in addition also significantly increased transcription of CD80, DECTIN-2, IDO1 and LAMP3 and decreased transcription of TLR7. Shiitake-βG and wheat-AX also significantly increased transcription of IL-1B, TNFA, LOX1 and CCL20 and decreased transcription of MGL. Finally, wheat-AX also significantly increased transcription of IRF1 and decreased transcription of CCL13. These results indicate that NDPs can indeed modify the gene transcription of M2-like macrophages, as they did for non-polarized macrophages.24 | ||
| Fig. 2 NDPs induced an M (NDP) transcriptional phenotype in M2-like macrophages. M2-like macrophages were stimulated with medium, yeast-βG, apple-RGI, shiitake-βG or wheat-AX at 500 μg ml−1 for 24 h and analyzed for gene transcription using qPCR. Bars represent mean fold change ± SD of n = 2–3 different donors. Statistically significant differences compared to medium control were analyzed by one-way ANOVA: * p < 0.05, ** p < 0.01, *** p < 0.001. | ||
To test whether NDPs similarly could impact M1-like macrophages, the transcription of the same set of marker genes was analyzed. Overall, NDPs modified gene transcription similarly as upon treatment of M2-like macrophages, albeit less pronounced (ESI Fig. 2†), with wheat-AX again being most effective. In contrast to modifications to the M2-like macrophage transcriptional profile, however, apple-RGI was more effective than shiitake-βG in inducing gene transcriptional changes in M1-like macrophages.
NDP-treatment decreased endocytosis and antigen processing capacity in M1- and M2-like macrophages
Upon identifying transcriptional modifications of M1- and M2-like macrophages by apple-RGI, shiitake-βG and wheat-AX, the functionality of NDP-treated macrophages was investigated regarding endocytosis and antigen processing. Wheat-AX-treatment of M1- and M2-like macrophages resulted in significantly reduced endocytosis of E. coli fragments and DQ-OVA processing (Fig. 3). Although similar to the impact on non-polarized macrophages (Fig. 1A and B), the inhibitory effect of wheat-AX was more pronounced in M1- and M2-like macrophages than in non-polarized macrophages. | ||
| Fig. 3 NDPs affected endocytosis and antigen procession activity in M1-like and M2-like macrophages. M1-like macrophages (A and C) and M2-like macrophages (B and D) were stimulated with medium, yeast-βG, apple-RGI, shiitake-βG or wheat-AX at 500 μg ml−1 for 24 h followed by 1 h incubation with E. coli-fragments (A and B) or DQ-OVA (C and D). E. coli-fragment and DQ-OVA uptake was analyzed by measuring MFI using flow cytometry. Bars represent mean values normalized to medium control (100%) ± SD of n = 3 different donors. Statistically significant differences compared to medium control were analyzed by one-way ANOVA: * p < 0.05, ** p < 0.01, *** p < 0.001. | ||
Apple-RGI and shiitake-βG non-significantly lowered E. coli uptake and significantly lowered DQ-OVA processing by M1-like macrophages which was similar to their effect on non-polarized macrophages. Upon exposure to M2-like macrophages, shiitake-βG also appeared to reduce E. coli uptake and DQ-OVA processing. In contrast, apple-RGI did not affect the M2-like macrophage functionality regarding E. coli uptake and DQ-OVA processing. Finally, yeast-βG did not modulate the endocytosis of E. coli fragments nor processing of DQ-OVA by M1- or M2-like re-polarized macrophages, similar to non-polarized macrophages.
Wheat-AX-treatment induces signaling molecule secretion in M2-like macrophages with strong chemotactic capacity towards monocytes
To test whether NDP-mediated altered functionality of M1- and M2-like macrophages was also reflected in alternative cytokine and chemokine production, and as a result monocyte recruitment, the cytokines and chemokines secreted by M1- and M2-like macrophages from a single donor were analyzed following incubation with apple-RGI, shiitake-βG and wheat-AX (Table 2). NDPs demonstrated to increase levels of cytokine and chemokine secretion of M1- and M2-like macrophages similar as they did upon exposure to non-polarized macrophages. Again, wheat-AX treatment of macrophages induced the highest levels of cytokine and chemokine secretion. Therefore, the conditioned medium from wheat-AX-treated and medium-treated M1- and M2-like macrophages was tested for their potency to attract monocytes (Fig. 4). Wheat-AX treatment in comparison to medium-incubation of M1-like macrophages did not lead to conditioned medium that increased levels of monocyte migration, which was in line with findings as shown in Fig. 1C. Surprisingly, conditioned medium from medium-incubated M2-like macrophages demonstrated similar levels of monocyte recruitment and conditioned medium from wheat-AX-treated M2-like macrophages revealed a significantly higher (>2 fold) recruitment of monocytes. | ||
| Fig. 4 Conditioned medium from wheat-AX-treated M2-like macrophages demonstrated increased monocyte recruitment capacity. M1-like macrophage and M2 macrophages were stimulated with medium or wheat-AX at 500 μg ml−1 for 24 h. Macrophage-conditioned medium was used to induce THP-1 monocyte migration measured with the xCeLLigence RTCA DP system. Bars represent average cumulative value over time ± SD of n = 3 different donors. Statistically significant differences were analyzed compared to non-treated macrophage-conditioned medium by one-way ANOVA: ns: not significant; ** p < 0.01. | ||
| Macrophage polarization status | M1 + medium | M1 + Apple-RGI | M1 + Shiitake-βG | M1 + Wheat-AX | M2 + medium | M2 + Apple-RGI | M2 + Shiitake-βG | M2 + Wheat-AX |
|---|---|---|---|---|---|---|---|---|
| M1- or M2-like macrophages were incubated with medium or treated with 500 μg ml−1 apple-RGI, shiitake-βG or wheat-AX for 24 h. Cytokine and chemokine secretion (pg ml−1) was analyzed with Bio-Plex multiplex analysis. Values represent average of a technical duplicate measurement of a single donor. | ||||||||
| CCL5 | 916 | 1898 | 705 | 4961 | 9.9 | 375 | 890 | 1929 |
| CCL20 | 17.8 | 110 | 13 | 1784 | 0.8 | 18 | 223 | 859 |
| CCL24 | 14.9 | 54 | 16 | 648 | 352 | 272 | 1970 | 1784 |
| CXCL8 | 921 | 15865 |
4336 | 19590 |
298 | 21968 |
23592 |
23213 |
| IL-1β | 0.2 | 0.3 | 0.2 | 3.1 | 0.1 | 0.2 | 1.3 | 4.1 |
| TNFα | 203 | 3035 | 284 | 30331 |
2.3 | 549 | 10012 |
23877 |
| CXCL9 | >24199 |
>24199 |
>24199 |
>24199 |
25.2 | 37.7 | 37.7 | 126 |
| CXCL10 | 224944 |
2603 | 699198 |
17327 |
13.5 | 18.7 | 28.8 | 64.8 |
| CXCL11 | 1188 | 36.6 | 410 | 698 | 0.1 | 0.1 | 0.2 | 0.4 |
Discussion
NDPs are considered immunomodulatory food components with variable bioactive properties. Here we investigated the functional impact of NDP-exposure to different macrophage phenotypes. NDP-treatment of non-polarized macrophages revealed that NDPs in general hampered typical M2-like macrophage functionality such as antigen endocytosis and processing (Fig. 1A and B).26 These findings are in line with reduced gene transcription in NDP-treated macrophages of NDP-binding lectin-receptors (i.e. CD302, MERTK and dectin-124,27,28) and HLA-class II molecules for lysosomal antigen loading (i.e. HLA-DM and HLA-DO (ESI Table 2†)29). Of note, gene transcription of the receptor for ovalbumin (i.e. mannose receptor),30 which is likely involved in the uptake of DQ-OVA, was not affected. Results further revealed that NDPs induced secretion of a unique mixture of chemokines and cytokines in non-polarized macrophages which is in line with the M(NDP) transcriptional phenotype. Cytokine and chemokine production patterns can be considered as markers to phenotype macrophages31 as CXCL9, CXCL10, CXCL11 and TNFα which induce monocyte recruitment are for M1-like macrophages (Table 1 and Fig. 1C). This is in line with literature which indicates that an influx of bone-marrow-derived monocytes replenishes intestinal macrophage populations especially in case of inflammation.20 NDPs, in particular wheat-AX, induced secretion of a signature mix of CCL1, CCL5, CCL20, CCL24, CXCL8 and IL-1β in the absence of CXCL9, CXCL10 and CXCL11 in non-polarized macrophages. CCL1 and CCL5 were shown in literature to mediate recruitment of CCR8- and CCR5-positive regulatory T cells,32,33 respectively, that might limit (chronic) inflammation in the intestine. In contrast, CCL20 is essential for migration of CCR6-positive TH17 cells towards the intestine,34 a cell type linked to chronic inflammatory disease,35 and IL-1β is crucial for Th17 polarization.36 Moreover, CCL24 was shown to induce inflammatory infiltration of eosinophils, neutrophils and basophils37 and CXCL8 has been shown to induce migration and activation of a large variety of immune cells.38 Conditioned medium from wheat-AX-treated macrophages instigated monocyte migration equal to the conditioned medium from M1-like macrophages (Fig. 1C). Theoretically the wheat-AX-treated macrophages could also induce migration of other immune cell types such as T cells (ESI Table 1†). Unfortunately, our model system appears limited to assessing migration of adherent cell types, preventing analysis of T cell migration as function of conditioned medium from treated macrophages. Previously, exposing non-polarized macrophages to various NDPs induced a general transcriptional phenotype that was termed M(NDP) and was distinct from M1- and M2-like macrophages.24 Functionally, however, the various NDPs have demonstrated clear differences exemplified by on the one hand soluble yeast-βG that apparently does not affect non-polarized macrophage functionality and on the other hand wheat-AX that strongly impacts non-polarized macrophage functionality. This indicates that the term M(NDP) to classify a phenotypical macrophage subset is limited in scope to transcriptional changes.In homeostasis intestinal macrophages are in general of an M2-like subset whereas in case of inflammation the macrophages are, locally, predominantly of the M1-like subset.18–20 Therefore M1- and M2-like macrophages were exposed to NDPs to assess the impact on their phenotype. NDPs, and again in particular wheat-AX, indeed induced gene transcription in M2-like macrophages and to some lesser extent M1-like macrophages similar to the M(NDP) transcriptional phenotype (Fig. 2 and ESI Fig. 2†). In addition, NDP-treatment similarly altered endocytosis and antigen processing of M1- and M2-like macrophages (Fig. 3). These changes in functional responses depend on both the NDP as well as the polarization state of the cells. When comparing apple-RGI and shiitake-βG, apple-RGI demonstrated more potency to alter M1-like macrophage functionality whereas shiitake-βG was more effective in changing M2-like macrophage functionality (Fig. 3 and Table 2). This suggests that the macrophage polarization status dictates towards which NDP it is most responsive, which in turn might relate to the expressed PRRs to recognize and bind NDPs.24 Indeed, PRRs dectin-1, LacCer, scavenger-, mannose-, and complement receptor were shown to bind to the shiitake-derived β-glucan termed lentinan28,39 of which transcription was higher in M2-like macrophages, whereas toll-like receptors 1 and 2 were shown to bind RG-I40 of which transcription was higher in M1-like macrophages.
Cytokine and chemokine secretion of M1- and M2-like macrophages following NDP-treatment (Table 2) revealed a similar unique mixture as NDP-treated non-polarized macrophages. The cumulative effect of wheat-AX-treatment of M2-like macrophages was the strongly increased recruitment of monocytes (Fig. 4). Results shown here on the induction of monocyte migration need to be extended to other immune cell population, but also investigated at lower NDP concentrations, to acquire a more complete overview of the recruitment potential of NDP-treated macrophages. Also, to increase translation to the in vivo situation, macrophages could be used that are phenotypically more similar to those found in the intestine. Recently, based on transcriptional profiling, four phenotypically different macrophage subsets were identified in the mucosa of duodenal-proximal jejunum in humans.17 The third subset, termed ‘Mf3’; and identified by CD45+HLA-DR+CD14+CD11c−CD11b−CXCR1low, is primarily located in the lamina propria and positioned to collected and transport luminal compounds. As these intestinal macrophages likely will be derived from circulating monocytes,18 further translational research could be based on protocols aimed to develop Mf3-like cells from PBMC-derived monocytes.
Clinical trials have demonstrated that β-glucans induced a lowered inflammatory status and increased activity and frequency of immune cells in patients.3,4,6–8 Furthermore, studies demonstrated that arabinoxylans but also lentinan reduce or prevent LPS-mediated inflammatory responses in macrophages.41,42 The oral administration of lentinan or arabinoxylan also provided beneficial health effects by significantly reducing intestinal inflammatory conditions in mice and oral administration of arabinoxylan resulted in significantly reducing serum inflammatory marker levels in patients and lowered PBMC responses.43–45 On the other hand, NDPs such as fucoidan and inulin-type fructan have demonstrated to support vaccination efficiencies by increasing titers towards influenza type B and hepatitis B, respectively, when orally consumed pre- and post-vaccination.46,47 Clinical research appears to show that NDPs modify immune responses towards a lowered activation state, but with enhanced vigilance. Our findings, albeit in vitro and with suboptimal macrophage phenotypes, are in line with these results. NDPs reduced M2-macrophage, but also M1-macrophage, capacity to endocytose and process antigen but increased the macrophage potential to recruit monocytes providing a rationale for the observed effects in clinical studies.
Conflicts of interest
There are no conflicts to declare.Acknowledgements
This work was supported by the European Community's Seventh Framework Programme [FP7/2007-2013] ‘EU Fibebiotics’ under Grant Agreement no. 289517, by the Dutch Cancer Society [WUR 2015-7734] ‘Food-derived beta-glucans and fungal proteins to support anti-melanoma immune therapy’, by the Dutch Ministry of Agriculture Nature and Food Quality, KB-23-001-012 and KB-23-001-009 and Yongfu Tang was funded by the China Scholarship Council.Authors wish to thank Dr Le Galle and Dr Saulnier from INRA (France) and Dr Rieder and Dr H. Knutsen from Nofima (Norway) for chemical characterization of NDPs; Prof. Debets from the Erasmus MC (Netherlands) for critically reviewing the manuscript and Dr Verschuren and the research group of life science analysis techniques of the Avans University of applied sciences for providing access to the xCELLigence apparatus.
References
- B. Novakovic, E. Habibi, S. Y. Wang, R. J. Arts, R. Davar, W. Megchelenbrink, B. Kim, T. Kuznetsova, M. Kox, J. Zwaag, F. Matarese, S. J. van Heeringen, E. M. Janssen-Megens, N. Sharifi, C. Wang, F. Keramati, V. Schoonenberg, P. Flicek, L. Clarke, P. Pickkers, S. Heath, I. Gut, M. G. Netea, J. H. Martens, C. Logie and H. G. Stunnenberg, β-Glucan Reverses the Epigenetic State of LPS-Induced Immunological Tolerance, Cell, 2016, 167, 1354–1368.e14 CrossRef CAS PubMed.
- I. Noss, G. Doekes, P. S. Thorne, D. J. Heederik and I. M. Wouters, Comparison of the potency of a variety of beta-glucans to induce cytokine production in human whole blood, Innate Immun., 2013, 19, 10–19 CrossRef CAS PubMed.
- K. Mosikanon, D. Arthan, A. Kettawan, R. Tungtrongchitr and P. Prangthip, Yeast beta-Glucan Modulates Inflammation and Waist Circumference in Overweight and Obese Subjects, J. Diet. Suppl., 2017, 14, 173–185 CrossRef CAS PubMed.
- D. Cholujova, J. Jakubikova, B. Czako, M. Martisova, L. Hunakova, J. Duraj, M. Mistrik and J. Sedlak, MGN-3 arabinoxylan rice bran modulates innate immunity in multiple myeloma patients, Cancer Immunol. Immunother., 2013, 62, 437–445 CrossRef CAS PubMed.
- X. Dai, J. M. Stanilka, C. A. Rowe, E. A. Esteves, C. Nieves Jr., S. J. Spaiser, M. C. Christman, B. Langkamp-Henken and S. S. Percival, Consuming Lentinula edodes (Shiitake) Mushrooms Daily Improves Human Immunity: A Randomized Dietary Intervention in Healthy Young Adults, J. Am. Coll. Nutr., 2015, 34, 478–487 CrossRef CAS PubMed.
- N. Kodama, K. Komuta and H. Nanba, Effect of Maitake (Grifola frondosa) D-Fraction on the activation of NK cells in cancer patients, J. Med. Food, 2003, 6, 371–377 CrossRef CAS PubMed.
- K. W. Tsang, C. L. Lam, C. Yan, J. C. Mak, G. C. Ooi, J. C. Ho, B. Lam, R. Man, J. S. Sham and W. K. Lam, Coriolus versicolor polysaccharide peptide slows progression of advanced non-small cell lung cancer, Respir. Med., 2003, 97, 618–624 CrossRef CAS PubMed.
- M. Jesenak, J. Majtan, Z. Rennerova, J. Kyselovic, P. Banovcin and M. Hrubisko, Immunomodulatory effect of pleuran (beta-glucan from Pleurotus ostreatus) in children with recurrent respiratory tract infections, Int. Immunopharmacol., 2013, 15, 395–399 CrossRef CAS PubMed.
- J. L. Buttriss and C. S. Stokes, Dietary fibre and health: an overview, Nutr. Bull., 2008, 33, 186–200 CrossRef.
- G. D. Brown, J. Herre, D. L. Williams, J. A. Willment, A. S. Marshall and S. Gordon, Dectin-1 mediates the biological effects of beta-glucans, J. Exp. Med., 2003, 197, 1119–1124 CrossRef CAS PubMed.
- S. Saijo, S. Ikeda, K. Yamabe, S. Kakuta, H. Ishigame, A. Akitsu, N. Fujikado, T. Kusaka, S. Kubo, S. H. Chung, R. Komatsu, N. Miura, Y. Adachi, N. Ohno, K. Shibuya, N. Yamamoto, K. Kawakami, S. Yamasaki, T. Saito, S. Akira and Y. Iwakura, Dectin-2 recognition of alpha-mannans and induction of Th17 cell differentiation is essential for host defense against Candida albicans, Immunity, 2010, 32, 681–691 CrossRef CAS PubMed.
- F. L. van de Veerdonk, R. J. Marijnissen, B. J. Kullberg, H. J. Koenen, S. C. Cheng, I. Joosten, W. B. van den Berg, D. L. Williams, J. W. van der Meer, L. A. Joosten and M. G. Netea, The macrophage mannose receptor induces IL-17 in response to Candida albicans, Cell Host Microbe, 2009, 5, 329–340 CrossRef CAS PubMed.
- V. Vetvicka, B. P. Thornton and G. D. Ross, Soluble beta-glucan polysaccharide binding to the lectin site of neutrophil or natural killer cell complement receptor type 3 (CD11b/CD18) generates a primed state of the receptor capable of mediating cytotoxicity of iC3b-opsonized target cells, J. Clin. Invest., 1996, 98, 50–61 CrossRef CAS PubMed.
- P. J. Rice, J. L. Kelley, G. Kogan, H. E. Ensley, J. H. Kalbfleisch, I. W. Browder and D. L. Williams, Human monocyte scavenger receptors are pattern recognition receptors for (1–>3)-beta-D-glucans, J. Leukocyte Biol., 2002, 72, 140–146 CAS.
- N. A. Mabbott, D. S. Donaldson, H. Ohno, I. R. Williams and A. Mahajan, Microfold (M) cells: important immunosurveillance posts in the intestinal epithelium, Mucosal Immunol., 2013, 6, 666–677 CrossRef CAS PubMed.
- G. Marelli, C. Belgiovine, A. Mantovani, M. Erreni and P. Allavena, Non-redundant role of the chemokine receptor CX3CR1 in the anti-inflammatory function of gut macrophages, Immunobiology, 2017, 222, 463–472 CrossRef CAS PubMed.
- A. Bujko, N. Atlasy, O. J. B. Landsverk, L. Richter, S. Yaqub, R. Horneland, O. Oyen, E. M. Aandahl, L. Aabakken, H. G. Stunnenberg, E. S. Baekkevold and F. L. Jahnsen, Transcriptional and functional profiling defines human small intestinal macrophage subsets, J. Exp. Med., 2018, 215, 441–458 CrossRef CAS PubMed.
- K. L. Flannigan, D. Geem, A. Harusato and T. L. Denning, Intestinal Antigen-Presenting Cells: Key Regulators of Immune Homeostasis and Inflammation, Am. J. Pathol., 2015, 185, 1809–1819 CrossRef CAS PubMed.
- I. Gabanyi, P. A. Muller, L. Feighery, T. Y. Oliveira, F. A. Costa-Pinto and D. Mucida, Neuro-immune Interactions Drive Tissue Programming in Intestinal Macrophages, Cell, 2016, 164, 378–391 CrossRef CAS PubMed.
- E. Zigmond, C. Varol, J. Farache, E. Elmaliah, A. T. Satpathy, G. Friedlander, M. Mack, N. Shpigel, I. G. Boneca, K. M. Murphy, G. Shakhar, Z. Halpern and S. Jung, Ly6C hi monocytes in the inflamed colon give rise to proinflammatory effector cells and migratory antigen-presenting cells, Immunity, 2012, 37, 1076–1090 CrossRef CAS PubMed.
- R. Rani, A. G. Smulian, D. R. Greaves, S. P. Hogan and D. R. Herbert, TGF-beta limits IL-33 production and promotes the resolution of colitis through regulation of macrophage function, Eur. J. Immunol., 2011, 41, 2000–2009 CrossRef CAS PubMed.
- R. J. Cummings, G. Barbet, G. Bongers, B. M. Hartmann, K. Gettler, L. Muniz, G. C. Furtado, J. Cho, S. A. Lira and J. M. Blander, Different tissue phagocytes sample apoptotic cells to direct distinct homeostasis programs, Nature, 2016, 539, 565–569 CrossRef CAS PubMed.
- F. Hong, J. Yan, J. T. Baran, D. J. Allendorf, R. D. Hansen, G. R. Ostroff, P. X. Xing, N. K. Cheung and G. D. Ross, Mechanism by which orally administered beta-1,3-glucans enhance the tumoricidal activity of antitumor monoclonal antibodies in murine tumor models, J. Immunol., 2004, 173, 797–806 CrossRef CAS PubMed.
- Y. Tang, C. Govers, H. J. Wichers and J. J. Mes, Macrophages treated with non-digestible polysaccharides reveal a transcriptionally unique phenotype, J. Funct. Foods, 2017, 36, 280–289 CrossRef CAS.
- C. Govers, M. M. M. Tomassen, A. Rieder, S. Ballance, S. H. Knutsen and J. J. Mes, Lipopolysaccharide quantification and alkali-based inactivation in polysaccharide preparations to enable in vitro immune modulatory studies, Bioact. Carbohydr. Diet. Fibre, 2016, 8, 15–25 CrossRef CAS.
- A. A. Tarique, J. Logan, E. Thomas, P. G. Holt, P. D. Sly and E. Fantino, Phenotypic, functional, and plasticity features of classical and alternatively activated human macrophages, Am. J. Respir. Cell Mol. Biol., 2015, 53, 676–688 CrossRef CAS PubMed.
- M. Kato, S. Khan, E. d'Aniello, K. J. McDonald and D. N. Hart, The novel endocytic and phagocytic C-Type lectin receptor DCL-1/CD302 on macrophages is colocalized with F-actin, suggesting a role in cell adhesion and migration, J. Immunol., 2007, 179, 6052–6063 CrossRef CAS PubMed.
- H. S. Goodridge, A. J. Wolf and D. M. Underhill, Beta-glucan recognition by the innate immune system, Immunol. Rev., 2009, 230, 38–50 CrossRef CAS PubMed.
- H. Kropshofer, A. B. Vogt, C. Thery, E. A. Armandola, B. C. Li, G. Moldenhauer, S. Amigorena and G. J. Hammerling, A role for HLA-DO as a co-chaperone of HLA-DM in peptide loading of MHC class II molecules, EMBO J., 1998, 17, 2971–2981 CrossRef CAS PubMed.
- S. Burgdorf, V. Lukacs-Kornek and C. Kurts, The mannose receptor mediates uptake of soluble but not of cell-associated antigen for cross-presentation, J. Immunol., 2006, 176, 6770–6776 CrossRef CAS PubMed.
- A. Mantovani, A. Sica, S. Sozzani, P. Allavena, A. Vecchi and M. Locati, The chemokine system in diverse forms of macrophage activation and polarization, Trends Immunol., 2004, 25, 677–686 CrossRef CAS PubMed.
- S. G. Kang, R. J. Piniecki, H. Hogenesch, H. W. Lim, E. Wiebke, S. E. Braun, S. Matsumoto and C. H. Kim, Identification of a chemokine network that recruits FoxP3(+) regulatory T cells into chronically inflamed intestine, Gastroenterology, 2007, 132, 966–981 CrossRef CAS PubMed.
- Y. Barsheshet, G. Wildbaum, E. Levy, A. Vitenshtein, C. Akinseye, J. Griggs, S. A. Lira and N. Karin, CCR8(+)FOXp3(+) Treg cells as master drivers of immune regulation, Proc. Natl. Acad. Sci. U. S. A., 2017, 114, 6086–6091 CrossRef CAS PubMed.
- E. Esplugues, S. Huber, N. Gagliani, A. E. Hauser, T. Town, Y. Y. Wan, W. O'Connor Jr., A. Rongvaux, N. Van Rooijen, A. M. Haberman, Y. Iwakura, V. K. Kuchroo, J. K. Kolls, J. A. Bluestone, K. C. Herold and R. A. Flavell, Control of TH17 cells occurs in the small intestine, Nature, 2011, 475, 514–518 CrossRef CAS PubMed.
- C. T. Weaver, C. O. Elson, L. A. Fouser and J. K. Kolls, The Th17 pathway and inflammatory diseases of the intestines, lungs, and skin, Annu. Rev. Pathol., 2013, 8, 477–512 CrossRef CAS PubMed.
- R. Mak'Anyengo, P. Duewell, C. Reichl, C. Horth, H. A. Lehr, S. Fischer, T. Clavel, G. Denk, S. Hohenester, S. Kobold, S. Endres, M. Schnurr and C. Bauer, Nlrp3-dependent IL-1beta inhibits CD103+ dendritic cell differentiation in the gut, JCI Insight, 2018, 3(5), e96322 CrossRef PubMed.
- A. Menzies-Gow, S. Ying, I. Sabroe, V. L. Stubbs, D. Soler, T. J. Williams and A. B. Kay, Eotaxin (CCL11) and eotaxin-2 (CCL24) induce recruitment of eosinophils, basophils, neutrophils, and macrophages as well as features of early- and late-phase allergic reactions following cutaneous injection in human atopic and nonatopic volunteers, J. Immunol., 2002, 169, 2712–2718 CrossRef CAS PubMed.
- R. C. Russo, C. C. Garcia, M. M. Teixeira and F. A. Amaral, The CXCL8/IL-8 chemokine family and its receptors in inflammatory diseases, Expert Rev. Clin. Immunol., 2014, 10, 593–619 CrossRef CAS PubMed.
- P. de Graaff, C. Govers, H. J. Wichers and R. Debets, Consumption of beta-glucans to spice up T cell treatment of tumors: a review, Expert Opin. Biol. Ther., 2018, 18, 1023–1040 CrossRef CAS PubMed.
- N. M. Sahasrabudhe, M. Beukema, L. Tian, B. Troost, J. Scholte, E. Bruininx, G. Bruggeman, M. van den Berg, A. Scheurink, H. A. Schols, M. M. Faas and P. de Vos, Dietary Fiber Pectin Directly Blocks Toll-Like Receptor 2–1 and Prevents Doxorubicin-Induced Ileitis, Front. Immunol., 2018, 9, 383 CrossRef PubMed.
- M. Mendis, E. Leclerc and S. Simsek, Arabinoxylan hydrolyzates as immunomodulators in lipopolysaccharide-induced RAW264.7 macrophages, Food Funct., 2016, 7, 3039–3045 RSC.
- M. M. M. Tomassen, E. A. H. J. Hendrix, A. S. M. Sonnenberg, H. J. Wichers and J. J. Mes, Variation of bioactive lentinan-containing preparations in Lentinula edodes strains and stored products, 7th International Conference of the World Society for Mushroom Biology and Mushroom Products, Arcachon, France, 2011, pp. 254–262 Search PubMed.
- Y. Nishitani, L. Zhang, M. Yoshida, T. Azuma, K. Kanazawa, T. Hashimoto and M. Mizuno, Intestinal anti-inflammatory activity of lentinan: influence on IL-8 and TNFR1 expression in intestinal epithelial cells, PLoS One, 2013, 8, e62441 CrossRef CAS PubMed.
- T. Kamiya, M. Shikano, M. Tanaka, K. Ozeki, M. Ebi, T. Katano, S. Hamano, H. Nishiwaki, H. Tsukamoto, T. Mizoshita, Y. Mori, E. Kubota, S. Tanida, H. Kataoka, N. Okuda and T. Joh, Therapeutic effects of biobran, modified arabinoxylan rice bran, in improving symptoms of diarrhea predominant or mixed type irritable bowel syndrome: a pilot, randomized controlled study, J. Evidence-Based Complementary Altern. Med., 2014, 2014, 828137 Search PubMed.
- B. N. Salden, F. J. Troost, E. Wilms, P. Truchado, R. Vilchez-Vargas, D. H. Pieper, R. Jauregui, M. Marzorati, T. van de Wiele, S. Possemiers and A. A. Masclee, Reinforcement of intestinal epithelial barrier by arabinoxylans in overweight and obese subjects: A randomized controlled trial: Arabinoxylans in gut barrier, Clin. Nutr., 2018, 37, 471–480 CrossRef CAS PubMed.
- H. Negishi, M. Mori, H. Mori and Y. Yamori, Supplementation of elderly Japanese men and women with fucoidan from seaweed increases immune responses to seasonal influenza vaccination, J. Nutr., 2013, 143, 1794–1798 CrossRef CAS PubMed.
- L. M. Vogt, M. E. Elderman, T. Borghuis, B. J. de Haan, M. M. Faas and P. de Vos, Chain length-dependent effects of inulin-type fructan dietary fiber on human systemic immune responses against hepatitis-B, Mol. Nutr. Food Res., 2017, 61(10), 1700171 CrossRef PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d0fo00316f |
| ‡ Shared first authorship. |
| This journal is © The Royal Society of Chemistry 2020 |