
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
A standardised semi-dynamic in vitro digestion method suitable for food – an international consensus†
Ana-Isabel
Mulet-Cabero
 a,
Lotti
Egger
b,
Reto
Portmann
b,
Olivia
Ménard
c,
Sébastien
Marze
d,
Mans
Minekus
e,
Steven
Le Feunteun
c,
Anwesha
Sarkar
f,
Myriam M.-L.
Grundy
g,
Frédéric
Carrière
h,
Matt
Golding
i,
Didier
Dupont
c,
Isidra
Recio
j,
André
Brodkorb
k and
Alan
Mackie
*f
a,
Lotti
Egger
b,
Reto
Portmann
b,
Olivia
Ménard
c,
Sébastien
Marze
d,
Mans
Minekus
e,
Steven
Le Feunteun
c,
Anwesha
Sarkar
f,
Myriam M.-L.
Grundy
g,
Frédéric
Carrière
h,
Matt
Golding
i,
Didier
Dupont
c,
Isidra
Recio
j,
André
Brodkorb
k and
Alan
Mackie
*f
aQuadram Institute Bioscience, Norwich Research Park, Norwich, NR4 7UQ, UK
bAgroscope, 3003 Bern, Switzerland
cSTLO, INRA, AGROCAMPUS OUEST, 35042, Rennes, France
dBIA, INRA, 44316 Nantes, France
eTriskelion, 3700 AV Zeist, The Netherlands
fSchool of Food Science & Nutrition, University of Leeds, Leeds, LS2 9JT, UK. E-mail: A.R.Mackie@leeds.ac.uk; Tel: +44 (0)113 34 32959
gUniversity of Reading, School of Agriculture, Policy and Development, Earley Gate, Reading RG6 6AR, UK
hAix-Marseille Université, CNRS, UMR7281 Bioénergétique et Ingénierie des Protéines, 31 chemin Joseph Aiguier, 13402 Marseille cedex 09, France
iSchool of Food and Nutrition, Massey University, Palmerston North, New Zealand
jInstituto de Investigación en Ciencias de la Alimentación (CIAL, CSIC-UAM), 28049 Madrid, Spain
kTeagasc Food Research Centre, Moorepark, Fermoy, County Cork, Ireland
First published on 10th February 2020
Abstract
The link between food and human health is increasingly a topic of interest. One avenue of study has been to assess food disintegration and interactions within the gastrointestinal tract. In vitro digestion models have been widely used to overcome the constrictions associated with in vivo methodology. The COST Action INFOGEST developed an international, harmonised protocol for static simulation of digestion in the upper gastrointestinal tract of adults. This protocol is widely used; however, it is restricted to providing end-point assessment without considering the possible structural changes. On the other hand, there are dynamic models that provide more physiologically relevant data but are expensive and difficult to access. There is a gap between these models. The method outlined in this article provides an intermediate model; it builds upon the harmonised static model and now includes crucial kinetic aspects associated with the gastric phase of digestion, including gradual acidification, fluid and enzyme secretion and emptying. This paper provides guidance and standardised recommendations of a physiologically relevant semi-dynamic in vitro simulation of upper gastrointestinal tract digestion, with particular focus on the gastric phase. Adaptations of this model have already been used to provide kinetic data on nutrient digestion and structural changes during the gastric phase that impact on nutrient absorption. Moreover, it provides a simple tool that can be used in a wide range of laboratories.
1. Introduction
The emergence of diet-related diseases and conditions such as obesity, diabetes, cardiovascular diseases and food allergy, led to a need to understand how different foods behave in the human gut, which will eventually help to develop healthier food products and formulate appropriate dietary advice. To obtain detailed information about the mechanisms behind the degradation/digestion of foods or specific nutrient(s), it is essential to monitor their behaviour within the different compartments of the gastrointestinal (GI) tract using consistent and well-controlled conditions.In vitro GI models can overcome many of the difficulties associated with human studies as the latter are often variable, costly, time-consuming and might generate ethical issues, depending on the study design and food being tested. However, the GI tract is a complex system that relies on a range of physical and biochemical processes (i.e. hormonal response, gastric emptying (GE), secretion of enzymes and digestive fluids, and motility) contingent on the individual and the food consumed. Sampling is often challenging in studies involving humans; the digesta may simply not be accessible without invasive procedures or it may not be homogeneous and thus not representative of the overall GI content. Moreover, the measurement of certain dynamic parameters, such as pH, may not be possible. Hence, an ideal model has to simulate the dynamic conditions of the different digestive compartments as closely as possible while keeping them consistent and rigorously monitored in order to measure the variable parameters of interest (e.g. products of digestion).1,2
Our previous harmonized in vitro digestion model developed by the COST Action INFOGEST addresses the key biochemical components of in vivo digestion and simulates the compartments of the digestive tract where most of the digestion and absorption of nutrients is taking place (i.e. oral, gastric and duodenal phases).3,4 However, this model suffers from some limitations. Gastric lipase, which is known to contribute to up to 25% (ref. 5) of lipid digestion in healthy adults,6,7 was not included in the previous protocol due to unavailability of commercial gastric lipases and non-relevant use of fungal lipases as gastric lipase substitutes in previous static models.8 Nevertheless, sources of gastric lipase are now suggested. In addition, our static model recommends using a constant gastric pH value of 3.0, which is a good compromise considering that pepsin has an activity maxima at pH 2 and still displays 70% thereof at pH 4.5.9 However, the optimum pH of gastric lipase lies between pH 4 and 6 depending on the triglyceride substrate,3,10 and salivary amylase activity, which is optimal between 6 and 7, is totally lost at pH 3.0.11 Even though in the fasted state the gastric pH is usually around 2, it is well recognized that consumption of a meal elevates the pH of the gastric food–fluid mixture, potentially to almost 7 (ref. 8, 9, 12–16) due to the buffering capacity of the food ingested, particularly driven by the major buffering nutrient (i.e. protein). Subsequently, this pH rise is followed by meal-stimulated secretion of HCl, which together with the progressive GE, gradually decreases the gastric pH back to the fasted state value. The dynamic nature of gastric secretion is not currently captured in the INFOGEST static model; however, it is crucial for understanding the pH-mediated changes occurring in food structure.16,17 For instance, during this transient acidification of the gastric content, “slow” proteins, such as caseins, are likely to coagulate around their isoelectric point resulting in a strong protein network with reduced accessibility to pepsin, which may result in delayed GE. On the other hand, “fast” whey proteins, which are partly soluble at low pH, might be easily emptied into the duodenum.18,19
It could be argued that sophisticated dynamic in vitro models developed by the Nederlandse Organisatie voor Toegepast Natuurwetenschappelijk Onderzoek (TNO), the former Institute of Food Research (IFR), Institut National de la Recherche Agronomique (INRA), etc., accurately simulate such meal-driven gastric secretions and represent in vivo conditions closely,20–22 however these computerised dynamic models are highly complex, time-consuming, require higher amounts of expensive enzymes and are thus far less accessible than static models.
Therefore, the aim of the current recommendation is to address these gaps and provide a semi-dynamic protocol based on the previous static version of the harmonized protocol with specific attention to mimicking the transient nature of gastric secretions and emptying. The overall objective of this work is to design a standardised semi-dynamic in vitro model of the digestion occurring in the upper GI tract of an adult human that can easily be used in laboratories across the world and to a wide range of foods.
2. Materials and equipment
2.1. Reagents
All chemicals are standard analytical grade. CaCl2(H2O)2 (Merck, 2382), NaOH (Merck, 9141), HCl (J. T. Baker, 6081), KCl (Merck, 4936), KH2PO4 (J. T. Baker, 0240), NaHCO3 (Merck, 6329); NaCl (Merck, 6404), MgCl2(H2O)6 (Merck, 5833), (NH4)2CO3 (Sigma-Aldrich, 207861). Enzyme inhibitor options: Pepstatin A (Sigma-Aldrich, P 4265), Orlistat (Sigma-Aldrich, O4139), Pefabloc SC (4-(2-aminoethyl)benzenesulfonyl fluoride (Sigma-Aldrich, 76307) and 4-bromophenylboronic acid (Sigma Aldrich, B75956). Ultrapure type I water, (e.g. generated by a Milli-Q® system or similar).2.2. Enzymes and activity determination
The type of enzyme products, mostly provided by Sigma Aldrich (St Louis, MO, USA), are only suggested examples and similar products of comparable quality from other providers can be used. Human salivary α-amylase (e.g. Sigma-Aldrich, A1031 or pooled human saliva (LEE Biosolutions, 991-05-P), porcine pepsin (Sigma-Aldrich, P7012), rabbit gastric extract (RGE) from Lipolytech®, France are selected as a preferred option as recommended in the static digestion protocol of INFOGEST.23 Note that the latter contains gastric lipase and pepsin24 (RGE 25–25 U mg−1 lipase and 800 U mg−1 pepsin – that corresponds to the lipase/pepsin ratio similar to the one found in humans). Bovine bile (Sigma-Aldrich, B3883) is selected as a preferred option. Alternatively, porcine bile extract can be used (Sigma-Aldrich, P8631). Pancreatin from porcine pancreas (Sigma-Aldrich, P7545) or, alternatively, individual pancreatic enzymes, namely, porcine trypsin (Sigma-Aldrich, T0303), porcine chymotrypsin (Sigma-Aldrich, C7762), pancreatic α-amylase (Sigma-Aldrich, A3176), porcine intestinal lipase (Sigma-Aldrich, L3126) and co-lipase (Sigma-Aldrich, C3028) can be used.The protocol and the chemicals needed for the determination of the activity have been detailed in the recently published protocol.23 The α-amylase (EC 3.2.1.1) activity is based on soluble potato starch: one unit liberates 1.0 mg of maltose equivalent from starch in 3 minutes at pH 6.9 at 20 °C. Rabbit gastric lipase (EC 3.1.1.3) activity is based on tributyrin as a substrate: one unit liberates 1 μmol butyric acid per minute at 37 °C and at pH 5.5. Porcine and/or rabbit pepsin (EC 3.4.23.1) activity is based on bovine blood haemoglobin as a substrate: one unit will produce a ΔA280 of 0.001 per minute at pH 2.0 and 37 °C, measured as TCA-soluble products. Porcine trypsin (EC 3.4.21.4) activity is based on p-toluene-sulfonyl-L-arginine methyl ester (TAME): one unit hydrolyses 1 μmol of TAME per minute at 25 °C, pH 8.1. Bovine chymotrypsin (EC 3.4.21.1) activity is based on N-benzoyl-L-tyrosine ethyl ester (BTEE): one unit hydrolyses 1.0 μmole of BTEE per minute at pH 7.8 at 25 °C. Porcine pancreatic lipase (EC 3.1.1.3) activity is based on tributyrin as a substrate: one unit liberates 1 μmol butyric acid per minute at 37 °C and at pH 8.0. Bile salt concentrations should be measured using a commercial kit (e.g. Sigma-Aldrich, MAK 309 or ECOLINR Acides Biliaires, DiaSys, 122129990313), or alternatively using LC-MS/MS analysis.25
2.3. Simulated digestive fluids
The electrolyte for the simulated fluids for the different phases, namely, electrolyte simulated salivary fluid (eSSF), electrolyte simulated gastric fluid (eSGF) and electrolyte simulated intestinal fluid (eSIF) are prepared by mixing the different electrolyte stock solutions, according to Table 1. The preparation of these electrolyte simulated digestion fluids at 1.25× concentration (400 mL) can be stored at −20 °C for one year or 2–5 °C for approximately one month. The addition of enzymes, Ca2+ solution and water will result in the correct electrolyte concentration in the final solution (1×) to provide the simulated digestive fluids, namely, simulated salivary fluid (SSF), simulated gastric fluid (SGF) and simulated intestinal fluid (SIF).| Salt solution added | Stock concentrations | Oral phase | Gastric phase | Smal intestinal phase | ||||
|---|---|---|---|---|---|---|---|---|
| mL of stock added to prepare 0.4 L (eSSF)b | Final salt conc. in SSF | mL of stock added to prepare 0.4 L (eSGF)b | Final salt conc. in SGF | mL of stock added to prepare 0.4 L (eSIF)b | Final salt conc. in SIF | |||
| pH7 | pH7 | pH7 | ||||||
| g L−1 | mol L−1 | mL | mmol L−1 | mL | mmol L−1 | mL | mmol L−1 | |
| a The use of a carbonate buffer requires that sealed containers are used with limited headspace. In open vessels, CO2 will be released and the pH will increase with time. However, as the pH is being altered anyway during the gastric phase, this might only be a problem during the intestinal phase. b eSSF, eSGF, eSIF are prepared at a concentration of 1.25× in order to add other solutions (e.g. HCl, enzymes) and to adjust the fluids ionic strength. c Addition immediately before the use. | ||||||||
| KCl | 37.3 | 0.5 | 15.1 | 15.1 | 6.9 | 6.9 | 6.8 | 6.8 |
| KH2PO4 | 68 | 0.5 | 3.7 | 3.7 | 0.9 | 0.9 | 0.8 | 0.8 |
| NaHCO3a | 84 | 1 | 6.8 | 13.6 | 12.5 | 25 | 42.5 | 85 |
| NaCl | 117 | 2 | — | — | 11.8 | 47.2 | 9.6 | 38.4 |
| MgCl2(H2O)6 | 30.5 | 0.15 | 0.5 | 0.15 | 0.4 | 0.12 | 1.1 | 0.33 |
| (NH4)2CO3a | 48 | 0.5 | 0.06 | 0.06 | 0.5 | 0.5 | — | — |
| CaCl2(H2O)2c | 44.1 | 0.3 | 0.025 | 1.5 | 0.005 | 0.15 | 0.044 | 0.6 |
2.4. Equipment
The type of equipment needed is listed below and is only meant as a guide to the general requirements and not as a recommendation of specific apparatus. | ||
| Fig. 1 Example of the apparatus of the gastric digestion phase. | ||
An indication of the basic material needed is as follows: Eppendorf tubes (2 mL), centrifuge plastic tubes (15 mL, 50 mL), pipettes (e.g. Gilson P200 and P1000, P5000) and tips, plastic syringes and serological pipettes (hand cut tip with, for instance, a blade to get a diameter of ∼3 mm). The latter is for the simulation of the gastric emptying. Volumetric flasks and glass beakers for preparation of solutions.
3. Preparations prior to the digestion procedure
3.1. Preparation of stock solutions of simulated digestion fluids
The electrolyte simulated fluids, namely, eSSF, eSGF and eSIF are prepared as described in the INFOGEST standardised static in vitro digestion.3 The only difference is that the eSGF is adjusted to pH 7 instead of pH 3 (Table 1) because the pH decrease during gastric phase will be obtained by a gradual addition of HCl.3.2. Determination of enzyme activities
The protocols for the assays of the enzymes to be used during oral, gastric and intestinal phases are described in the recently published Nature Protocols article by Brodkorb, et al.23 It is important to note that RGE contains both gastric lipase and pepsin,24 therefore the activity of both enzymes should be measured. If the pepsin activity in the RGE is not enough in order to reach the final activity required (4000 U mL−1 in SGF), this should be supplemented with pepsin from porcine origin.3.3. Determination of dry weight of food
The sample to be digested referred to as “food” should represent a real meal that can be consumed and be related to a serving of food. Thus, it should contain a quantity of liquid that is typically consumed together with the food.26 Low moisture solids, for instance cheese or bread, are often consumed together with liquids. Therefore, this dilution should be considered when applying this digestion model. The dry weight (or dry matter) of food to be digested should be measured/known since it is needed for the oral phase calculations. The ratio of SSF added to dry weight of the food is defined as 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1.
:1.
3.4. Determination of caloric content of food
GE is based on the caloric content of food to be digested (kcal per gram of food). Therefore, this value needs to be known. The caloric content is found in the package of the commercial foods. Otherwise, this can be determined by direct energy measurement or calculated theoretically. The caloric content can be calculated by knowing the composition of protein, lipid and carbohydrate in the sample and applying the standard Atwater factors (1 g of lipid yields 9 kcal, 1 g of protein yields 4 kcal and 1 g of carbohydrates yields 4 kcal).3.5. pH test tube
This test is performed in order to determine the volume and concentration of HCl needed to decrease the pH of the tested food to pH 2. The method, described in ESI 2,† excludes two opposing factors, i.e. the gastric enzymes and the GE. Adding the enzymes will increase the buffering capacity of the system while the emptying will lower it. By excluding both factors, the aim is to have a simple compromise solution that still gives a useful result.A mock digestion experiment under the conditions of the actual digestion is recommended to check the final pH, which should reach pH 2 at the end of gastric phase as accurately as possible.
3.6. Practical considerations and calculations for simulation of gastric dynamics
Some aspects need to be considered before digestion. (i) The amount and type of food; this will be subject to the capacity of the reaction vessel and the amount of food available. In addition, the amount of food at the last points of gastric phase should be enough in order to keep the pH probe, located inside the vessel, covered and working properly. For practicality in the calculations, the amount of food selected is scaled down from a standard meal volume of 500 mL. This value is taken as reference but it can be changed in the spreadsheet provided (ESI 3†) if required. It needs to be realistic for the digested food, e.g. a yogurt could be 150 mL and milk could be 250 mL. (ii) Number of GE aliquots; emptying refers to the food that is delivered from the stomach to the intestine through the pylorus, which acts as a sieve allowing liquid and particles of less than approximately 3 mm to pass through. We recommend a minimum of three aliquots to achieve results showing the kinetics of nutrient digestion. (iii) The initial food and the collected aliquots can be considered by weight if food undergoes large changes in consistency during the gastric phase. The simulated fluids and other solutions used during the digestion are considered to have a density of 1 g m−3. (iv) It is important to note that the pH values that can be recorded inside of the vessel might be different to the emptied aliquots due to the intra-gastric heterogeneity. (v) The emptying is preferably performed in a stepwise manner using a pipette with a modified tip with an end inner diameter of ∼3 mm. Another option is a syringe with attached tubing with the same end size. Alternatively, this can be achieved continuously using a peristaltic pump. Emptying should be from the bottom part of the vessel. It should be noted that sampling (different from emptying) can also be performed during gastric phase as considered appropriate. (vi) The mixing during the gastric phase depends on the consistency of the sample during the gastric phase. The use of an orbital shaker at 35 rpm should be adequate in samples that do not present significant changes to provide enough mixing with the fluids. However, there are systems with complex nature and/or samples in which the consistency changes drastically during the gastric phase, which makes the emptying and sampling difficult. In these cases, we recommend the use of an overhead stirrer with a paddle type stirrer blade placed in the lowest possible part of the vessel and using low speed (e.g. 15 rpm). (vii) In the fasted state, the stomach retains a certain basal level of secretion. We propose to use 10% of the total gastric secretion to provide that basal volume (taken as equal to the volume of the total oral mixture). In this way, the pH of the food will drop as soon as the food is added to the simulated stomach and the pH of the gastric content will be higher than the gastric secretion.Once the parameters needed (initial amount of food, dry weight, HCl volume and concentration, caloric content per gram of food, enzyme activities, number of GE points) have been decided/measured, these can be entered in the spread sheet provided (ESI 3†). This will automatically calculate the amount of the remaining solutions needed as well as the volume and time of emptying. The calculations for the gastric emptying dynamics are based on the delivery of 2 kcal min−1 of a food volume of 500 mL based on in vivo considerations. This means that the digestion time is proportional to the energy content of the food but it does not scale with the in vitro volume of food only with the “in vivo” volume of the food. Table 2 shows an example of the calculations used in that dynamic step.
| Food sample (example) | |
| Food volume (mL) | 20 |
| Energy content (kcal mL−1) | 0.80 |
| Total solids (g) | 3.0 |
| Gastric emptying and digestion time | ||
|---|---|---|
| In vitro | In vivo | |
| Food volume (mL) | 20.0 | 500.0 |
| Oral volume (mL) | 3.0 | 75.0 |
| Basal volume (mL) | 2.3 | 57.5 |
| Gastric volume (Food + oral + basal) at t = 0 (mL) | 25.3 | 632.5 |
| Energy content of food (kcal) | 16.0 | 401 |
| Energy emptying rate (kcal min−1) | 0.08 | 2.00 |
| Volume emptying rate (mL min−1) (emptied in 5 steps of 9.2 mL every 40 min) | 0.13 | 3.15 |
| Gastric halftime, t1/2 (min) | 100.3 | 100.3 |
| Total digestion time (min) | 200.5 | |
4. Semi-dynamic digestion protocol
Fig. 2 shows an overview of the semi-dynamic digestion protocol, which builds on the previous static version.3,23 The details regarding the choice and concentration of chemicals, inclusion or omission of certain steps and their justification are discussed in the first paper3 and some issues are corrected and complemented in the recently published paper by Brodkorb, et al.23 A detailed example of the parameters used in the three digestion phases of the semi-dynamic digestion model is illustrated in Table 3.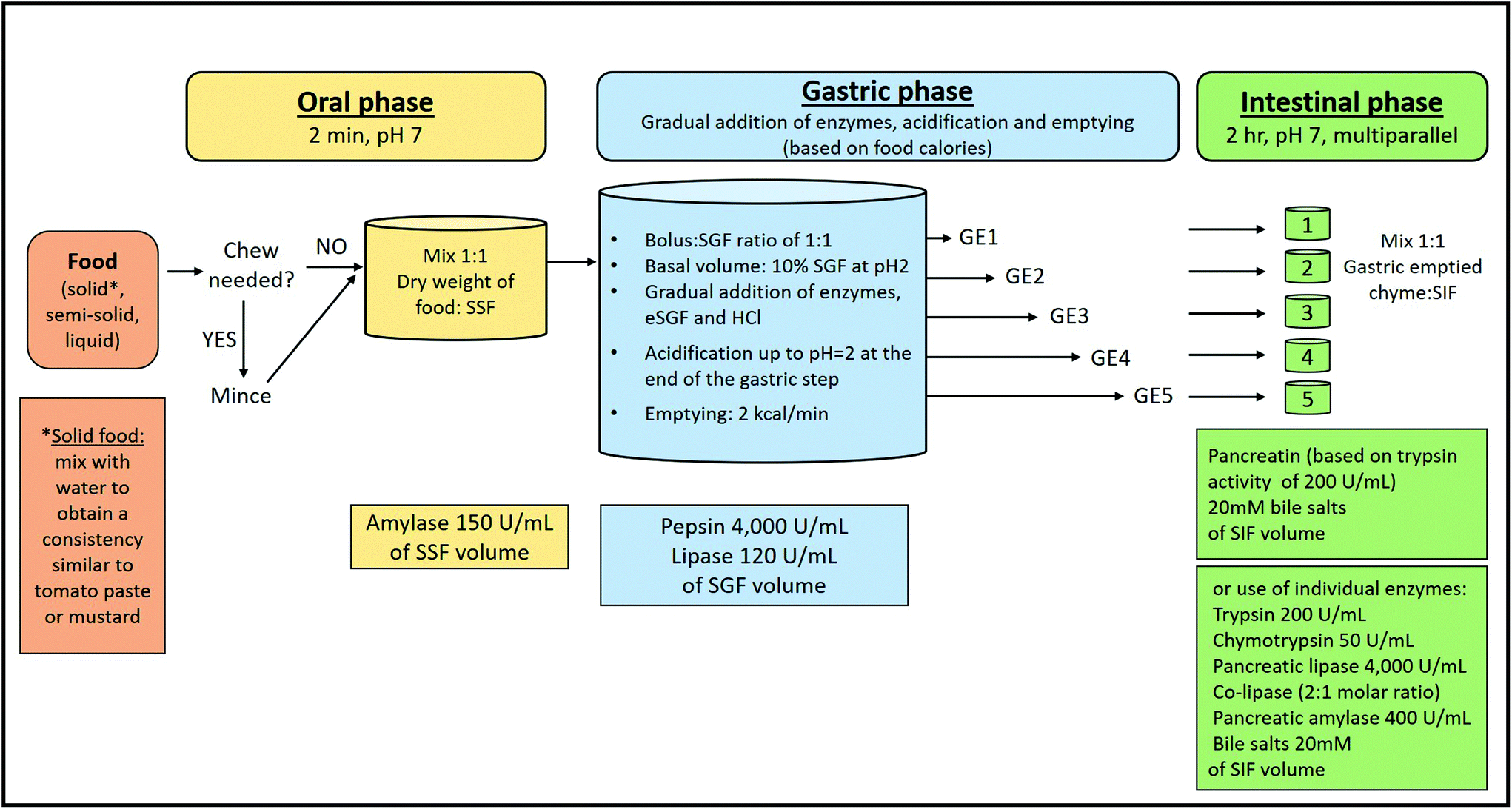 | ||
| Fig. 2 Overview and flow diagram of the simulated semi-dynamic in vitro digestion method. Simulated salivary fluid (SSF), simulated gastric fluid (SGF), simulated intestinal fluid (SIF) and gastric emptying (GE). Enzyme activities are in units per mL of the simulated digestive solution added at each digestion phase (SSF, SGF or SIF). | ||
| Food sample (example) | |
| Food volume (mL) | 20 |
| Energy content (kcal mL−1) | 0.80 |
| Total solids (g) | 3.0 |
| 1. Oral static digestion (37 °C, high shear, 2 min) | ||
|---|---|---|
| Compound | SSF (mL) | SSF (%) |
| eSSF | 2.40 | 80 |
| CaCl2(H2O)2 (1.5 mmol L−1 in SSF) | 0.015 | 0.5 |
| α-Amylase (150 U mL−1 in SSF) | 0.15 | 5.0 |
| MilliQ® water | 0.435 | 14.5 |
| Total | 3.00 | 100 |
| 2. Gastric dynamic digestion (37 °C, low/medium shear, duration depends on food calories) | |||||
|---|---|---|---|---|---|
| Compound | SGF (mL) | SGF (%) | Basal (mL) | Simulated gastric electrolyte mixture (mL). Rate 0.093 mL min−1 | Enzyme solution (mL). Rate 0.010 mL min−1 |
| eSGF | 16.1 | 70.0 | 1.61 | 14.49 | — |
| CaCl2(H2O)2 (0.15 mmol L−1 in SGF) | 0.01 | 0.04 | 0.001 | 0.009 | — |
| MilliQ® water | 2.59 | 7.78 | 0.259 | 2.331 | — |
| HCl (1.5 mol L−1) | 2 | 8.7 | 0.20 | 1.80 | — |
| Pepsin 4000 U mL−1 and gastric lipase 120 U mL−1 in SGF (enzyme solutions made in eSGF) | 2.3 | 10 | 0.23 | — | 2.07 |
| Total | 23 | 100 | 2.3 | 18.63 | 2.07 |
| 3. Small intestinal static digestion for individual GE aliquot (e.g. 9.2 mL) (37 °C, high shear, 2 hours) | ||
|---|---|---|
| Compound | SIF (mL) | SIF (%) |
| eSIF | 5.75 | 62.5 |
| CaCl2(H2O)2 (0.6 mmol L−1 in SIF) | 0.018 | 0.2 |
| Bile salt solution (20 mmol L−1 in SIF) | 1.15 | 12.5 |
| NaOH (2 mol L−1) | 0.5 | 5.43 |
| Pancreatin solution (200 U mL−1 in eSIF, based on trypsin) (enzyme solution made in eSIF) | 0.46 | 5.0 |
| MilliQ® water | 1.322 | 14.37 |
| Total | 9.2 | 100 |
4.1. Oral phase
The inclusion of an oral phase is recommended for all foods, solid or liquid, in order to be consistent with dilution factors. The amount of salivary secretion added corresponds to a final ratio of dry weight of food to SSF of 1:1 (w/v). The addition of amylase will not be required in the absence of starch. For starch containing food, amylase is added to achieve 150 U mL−1 in the SSF, followed by CaCl2(H2O)2 to achieve 1.5 mmol L−1 in SSF and the necessary amount of water to obtain the required concentration of SSF. Users of pancreatic amylase should be aware that it may retain some protease activity and this should be checked. The recommended time of contact with the enzyme is two minutes at 37 °C, which requires the pre-warming of all reagents to 37 °C. A detailed step by step procedure can be found in ESI 4.†
Solid foods need to be chewed, which is simulated by mincing an appropriate amount of food using a commercially available manual or electric mincer, commonly used in kitchens to mince meat. If necessary, water can also be added during mincing.
4.2. Gastric phase
The gastric phase is intended to be dynamic and thus the simulated secretions are added gradually. The fasting level is 10% of the total amount of gastric secretion to be added. The justification of this value is outlined below. As with the static protocol the aim is to obtain a final ratio of oral phase content to SGF of 1:1 (v/v) after addition of all other components.
In addition to changes in volume, the pH also needs to decrease from an initially high level determined by the buffering capacity of the food to pH 2. This is achieved by adding the 10% of SGF volume in the gastric reaction vessel at the start of the gastric phase. The remaining 90% of the simulated gastric electrolyte mixture, which contains eSGF, CaCl2(H2O)2 and water without enzymes, is gradually delivered and has a pH defined by the volume of HCl added to reach pH 2 at the end of the gastric digestion. It is recommended that the amount of acid required is determined in a test experiment prior to digestion. The remaining 90% of enzyme volume is also gradually delivered in a separate device(s). We recommend the addition of gastric lipase using rabbit gastric extract (RGE) to provide 120 U mL−1 of lipase in the final SGF solution, which corresponds to the average gastric lipase activity in the human gastric juice.27 Pepsin should be added to provide 2000 U mL−1 in SGF solution. The enzymes should be dissolved in eSGF. The reaction vessel needs to be held at 37 °C and a small water jacketed vessel is recommended. Very limited mixing during digestion is recommended, especially in the top part of the reaction vessel. This is in order to mimic the in vivo situation where only the antrum experiences significant shear. This can be achieved by using an overhead stirrer with a stirrer head turned at a rate of 15 rpm. The template for this design is given in the attachments of the ESI 1.†
The gastric phase also includes emptying into the intestinal phase and in order to keep things simple we suggest a constant emptying rate based on the energy content of the food (scaled from 2 kcal min−1 in what would be fed in vivo, see Table 2). The emptying is performed in a step-wise manner by manually taking the selected aliquots at calculated times from the bottom of the vessel using a selected laboratory tool with an end diameter of ∼3 mm.
A detailed step by step procedure can be found in ESI 4.†
4.3. Intestinal phase
This part of the protocol remains similar to the static version published previously and is performed individually with each aliquot emptied from the gastric phase (Fig. 2). The aim is to obtain a final ratio of emptied digesta from gastric phase to SIF of 1:1 (v/v) after addition of all other components, which includes eSIF, enzyme solution, bile, water, CaCl2(H2O)2 and base. Addition of base (e.g. 1 mol L−1 NaOH) will be required to neutralise the mixture to pH 7.0 in case it was not added to stop the pepsin activity in the gastric phase. Digestive enzymes can be added as either pancreatin from porcine pancreas or individual enzymes. The amount of pancreatin added is based on the trypsin activity (200 U mL−1 TAME activity in the SIF). If the food contains lipid and lipid digestion is at the centre of the study, pancreatin concentration should be either based on the lipase activity and if necessary, additional porcine pancreatic lipase and colipase should be added to achieve 2000 U mL−1 lipase activity in the final mixture. Alternatively, individual enzymes can be added to the digestion mixture to achieve the following activities in the final mixture: porcine trypsin (200 U mL−1 in SIF), bovine chymotrypsin (50 U mL−1 in SIF), porcine pancreatic α-amylase (400 U mL−1 in SIF), porcine pancreatic lipase (4000 U mL−1 in SIF) and porcine pancreatic colipase (2:1 colipase to lipase molar excess, equivalent to a mass ratio of roughly 1:2 co-lipase to lipase as the mass of human pancreatic lipase is 51.2 kDa and the mass of human co-lipase is 10 kDa). Bile is added to give a final concentration of 20 mmol L−1 of bile salts of the SIF volume. CaCl2(H2O)2 is added to reach 0.6 mmol L−1 in the SIF. The pH may need re-adjustment before finally adding water to the mixture to obtain the required concentration of SIF. The pH may need re-adjustment during digestion. This can be achieved either manually or by automated laboratory titrator. The recommended time of intestinal digestion is two hours at 37 °C.
A detailed step by step procedure can be found in ESI 4.†
4.4. Sampling and stopping reaction
Sample conservation depends on the study focus (food structure, bioaccessibility, enzymatic digestion product, etc.), and should be carefully considered for each study. It may be advisable to perform the intestinal phase in the tube where the emptied aliquot from the gastric phase is collected. Sampling during gastric phase can be performed and should take into consideration that the sample in the gastric compartment might experience structural changes and might not be homogenous.In order to stop the reaction after the gastric digestion the pH can be raised to 7.0 with NaOH or NaHCO3 (1 mol L−1 or higher concentration), which is similar to the in vivo conditions of the small intestine. An alternative is to stop pepsin activity by adding pepstatin A (7.2 μmol L−1 final concentration in the digesta). To stop gastric lipase activity, the use of Orlistat® (tetrahydrolipstatin) is recommended at a final concentration of 1 mmol L−1. After intestinal digestion, Pefabloc® SC (4-(2-aminoethyl)-benzolsulfonylfluorid-hydrochloride) can be added for serine protease (trypsin and chymotrypsin) inhibition (5 mmol L−1 final concentration). The addition of a lipase inhibitor such as 4-bromophenylboronic acid (5 mmol L−1 final concentration) is recommended to inhibit lipolysis by pancreatic lipase. After the addition of the inhibitor, or the pH drift, it is recommended to snap freeze in liquid nitrogen. Alternatively, both gastric and pancreatic lipases can be irreversibly inactivated by using immediately the samples for lipid extraction and analysis, e.g. by sample acidification with HCl and addition of organic solvents. Finally, when starch digestion is being studied, amylase activity should also be stopped at both the gastric and intestinal stages. To do so, it is recommended to heat the samples at 100 °C for 5 min immediately after their collection.28
5. Anticipated results
The semi-dynamic digestion model allows the study of structural changes of foods during simulated gastric digestion, evaluating the effect of the matrix on food disintegration and nutrient delivery to the small intestine. Mulet-Cabero, et al.19 used the semi-dynamic model and assessed several physico-chemical parameters in the emptied aliquots to follow the digestion kinetics of nutrients. Moreover, an adaptation of this model, using in vivo data, provided a close simulation of the structural changes in the gastric phase of two dairy foods when compared with the human stomach.29 In this section, we provide some indicative examples of results that can be obtained using the semi-dynamic model.5.1. Structural changes in milk proteins during semi-dynamic gastric digestion
The following results are based on two milk protein solutions at 8% (w/w) made of whey protein isolated (WPI), BiPRO, purchased from Davisco (Foods international NC, USA) and milk protein concentrate (MPC), Solmiko MPC 80, obtained from Glanbia Ingredients, Ireland.The gastric behaviour of tested foods was visually followed (Fig. 3). The WPI sample presented some precipitation in early stage of gastric digestion but it was dissolved in the course of the digestion, resulting in a clear solution. In contrast, the MPC sample presented strong aggregation and, then, the formation of a tight coagulum that was persistent until the end of the digestion.
 | ||
| Fig. 3 Images of the gastric behaviour of the WPI and MPC solutions at the time points corresponding to different gastric emptying (GE) points: GE1 (16 min), GE3 (48 min) and GE5 (80 min). Note that these gastric emptying time points are specific for these samples according to their caloric content (8% protein). The images were taken immediately before the emptying. | ||
Fig. 4 shows the pH profile obtained during gastric digestion. The initial pH was around 1 simulating the basal stage of the stomach. Following the addition of food, the pH increased rapidly due to the buffering capacity of the food, after which the pH decreased gradually back to the basal conditions.
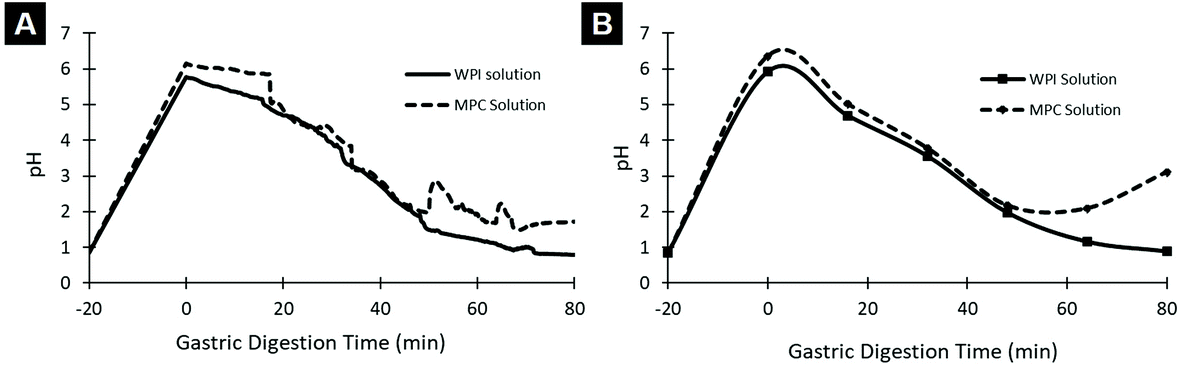 | ||
| Fig. 4 Example of the pH profile of WPI and MPC solutions. The pH curves were obtained using (A) the pH probe inside the vessel, which measures the digesta remaining in the simulated stomach and (B) pH values in each emptied aliquot. | ||
The nutrient composition in each emptied aliquot can be determined, providing the extent and rate at which the nutrients are released from the simulated stomach to the small intestine. As seen in Fig. 5, the WPI sample presented high protein delivery in the first points of gastric digestion with a decreasing pattern, showing the lowest protein delivery at the last GE point. This contrasts to the pattern observed in the MPC sample, in which there was a relatively low and constant delivery of protein at the beginning of the digestion ending with a significant increase at the GE5 point. These trends are linked with the structural changes in the gastric compartment observed in Fig. 3; the formation of that firm coagulum in the MPC sample delayed the emptying of proteins in comparison to the more homogenous digesta obtained in the WPI sample.
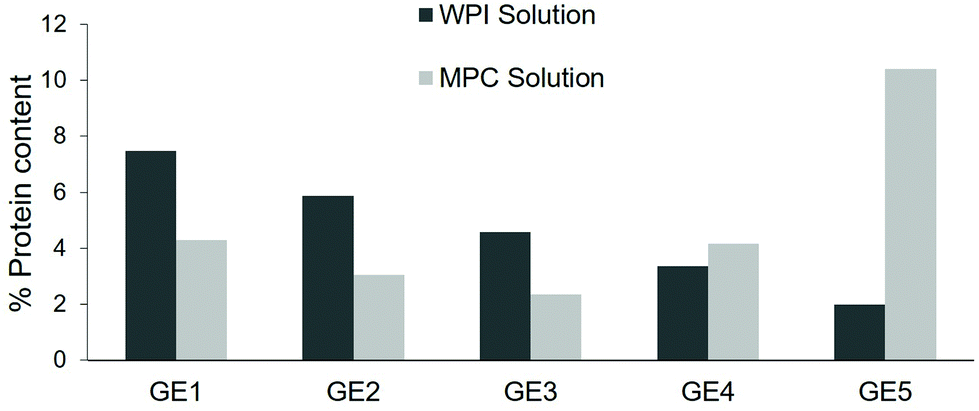 | ||
| Fig. 5 Protein content (%, w/w) in each gastric emptying (GE) aliquot selected to simulate the gastric emptying. GE1, GE2, GE3, GE4 and GE5 correspond to the gastric times of 16, 32, 48, 64 and 80 min, respectively. The protein content was determined by the Dumas method as previously described in Mulet-Cabero, et al.19 A conversion factor of 6.38 was used to obtain the protein content from the nitrogen content. Data represent one replicate as indicative of the results that can be obtained. | ||
5.2. Evolution of protein hydrolysis of skim milk powder during semi-dynamic digestion
Gradual protein hydrolysis in skim milk powder (SMP) was followed during semi-dynamic digestion from whole proteins to peptides and total free amino acids using sodium dodecyl sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), liquid chromatography-mass spectrophotometry (LC-MS) and the o-phthaldialdehyde (OPA) method, as previously described elsewhere.30Electrophoretic separation in samples emptied (from the bottom of the vessel, using a tip of ∼3 mm internal diameter) after 8 and 16 min of gastric digestion showed major bands from intact caseins (Fig. 6). A clear evolution in casein hydrolysis was observed in samples emptied between 24 and 49 min and in later samples no intact casein bands were detected. In contrast to this, intact β-lactoglobulin was visible until the end of gastric digestion, although with a decrease in intensity (possibly through dilution of the sample due to addition of simulated gastric fluid). On the other hand, no bands from intact milk proteins could be observed in the emptied gastric aliquots after intestinal digestion, indicating that they were rapidly hydrolysed at the beginning of the intestinal phase independently of the duration of the gastric phase.
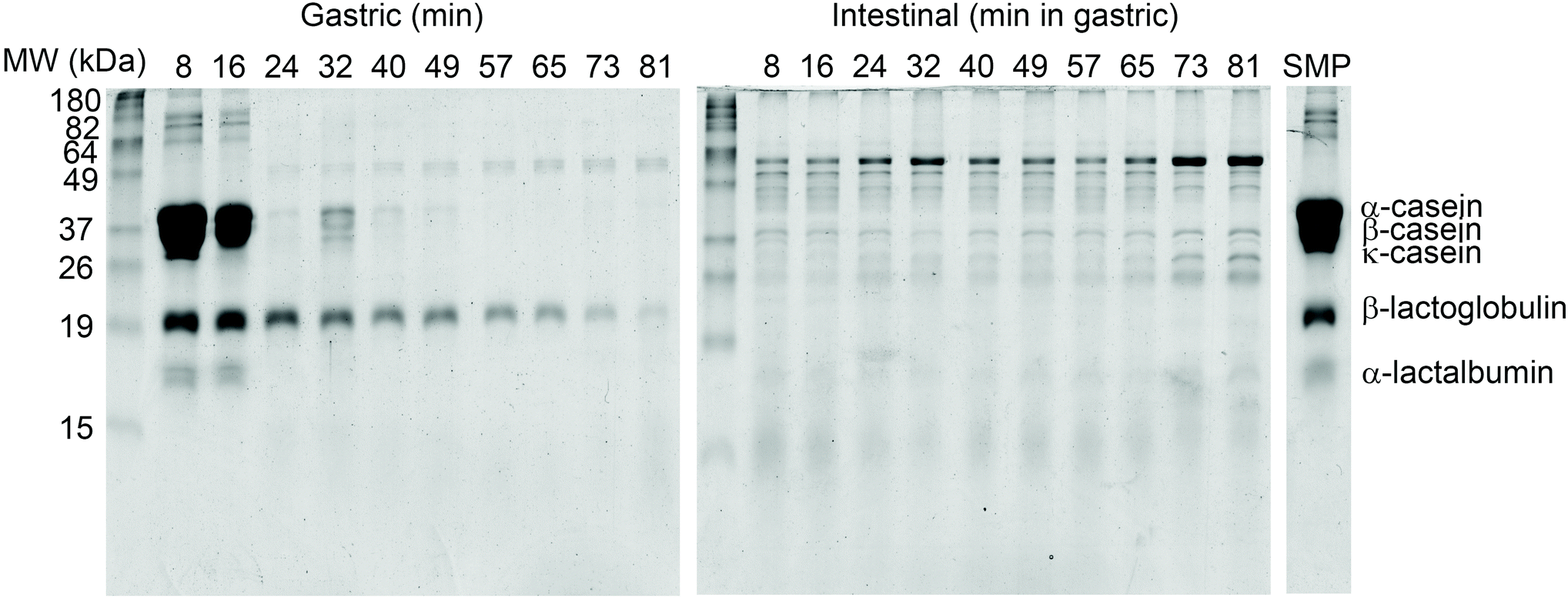 | ||
| Fig. 6 SDS-PAGE of protein hydrolysis during semi-dynamic digestion of SMP. For each of the gastric emptying aliquots (gastric) an individual static intestinal digestion for 120 min was performed (intestinal) and separated on gel. Undigested SMP is shown as control. | ||
All samples were analysed by LC-MS and the evolution during the gastric phase was clearly visible for αs1-casein, shown as low peptide abundance (blue) for the earlier gastric emptying points and increasing at later time points (Fig. 7). Only a few casein peptides were visible in all intestinal samples. In contrast, few β-lactoglobulin peptides were detected in the gastric samples and increasing intensities were observed in the intestinal phase, confirming previously published data on the resistance of this protein to gastric digestion. The individual gastric emptying points were performed without prior homogenization. Thus, depending on the solubility of the food and the formation of a coagulum during the gastric phase, the collected samples had a higher variability. For example, in the sample emptied after 32 min of gastric digestion, caseins were less hydrolysed compared to the previous sample emptied after 24 min. This variability was visible at the level of whole proteins (Fig. 6) and peptides (Fig. 7), but no longer at the level of free amino acids (Fig. 8).
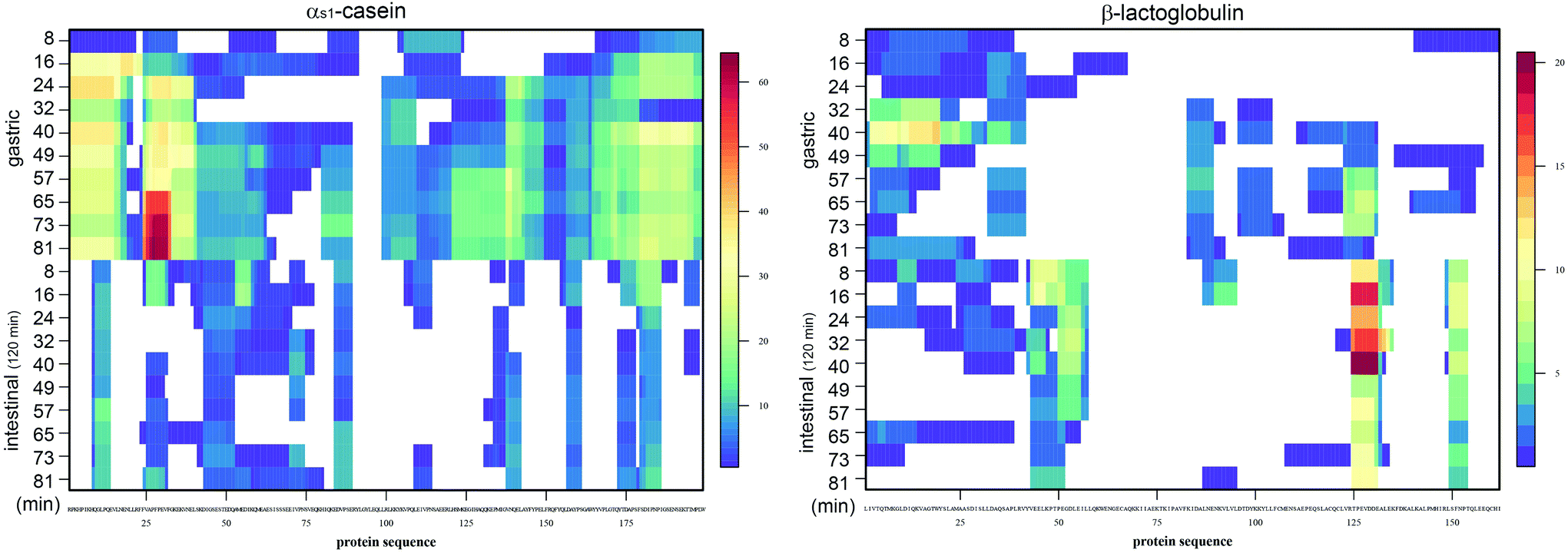 | ||
| Fig. 7 Peptide patterns after LC-MS analysis as previously described 27 for αs1-casein and β-lactoglobulin after different times of semi-dynamic gastric digestions (gastric), followed by 120 min of static intestinal digestion (intestinal) individually for each gastric sample. The colour index shown for both proteins indicates blue colour for low abundance and red colour for high abundance of amino acids identified within the protein sequence. White stretches indicate no identified protein regions. | ||
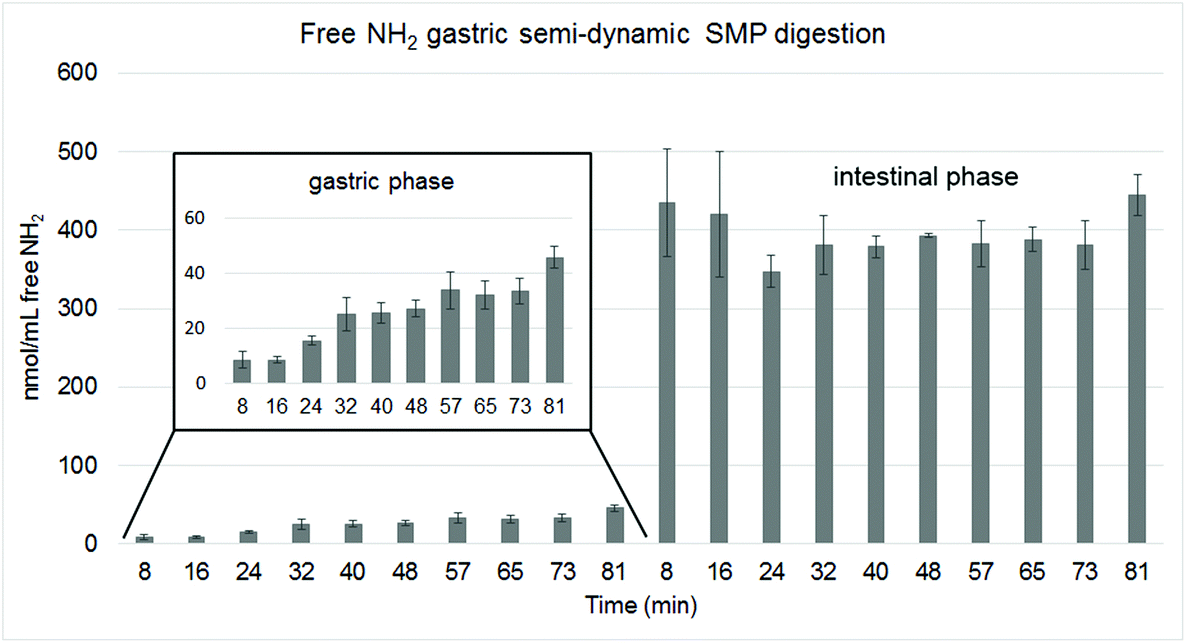 | ||
| Fig. 8 Release of total free NH2 groups analysed with the OPA method in mmol per mL of SMP. The different gastric emptying time points were individually subjected to intestinal digestion during 120 min. The inlet graph shows the evolution in NH2 release during gastric digestion using a smaller scale. | ||
The analysis of total free NH2 groups with the OPA method showed that only a small amount of free amino groups was released during the gastric phase with a constant slight increase during this phase (Fig. 8, inlet graph). All intestinal samples had a similar high content of free NH2 at the end (Fig. 8), indicating that protein hydrolysis was equal also for samples with shorter gastric digestion times.
5.3. Evolution of lipid hydrolysis in presence of gastric lipase
The effects of gastric lipase on lipid and protein hydrolysis were investigated during semi-dynamic digestion of whole homogenised ultra-high temperature processed milk. Time resolved digestions were either performed in the presence or absence of gastric lipase, using rabbit gastric extract (RGE) as source for gastric lipase. Total free fatty acids (FFA, non-esterified fatty acid kit, Fujifilm Wako Diagnostics, US) were quantified with a colorimetric enzymatic method for non-esterified fatty acids.31 As expected, a gradual liberation of free fatty acids was only observed in gastric samples taken at different time points (16, 32, 48, 64, 80, 96, 112, 128, 144, 160 min) where gastric lipase was present, whereas no free fatty acids were detected in samples without lipase taken at the same time points (Fig. 9, inlet graph). In the intestinal phase which was individually performed for 120 min for each gastric endpoint, this higher free fatty acid release persisted in the samples containing gastric lipase until the end of intestinal digestion, leading to a higher fatty acid hydrolysis in total of these samples (Fig. 9). The high increase observed in the intestinal phase from the last gastric aliquot was due to the formation of a cream layer in the gastric phase leading to the emptying of a higher amount of lipid at the end of the digestion.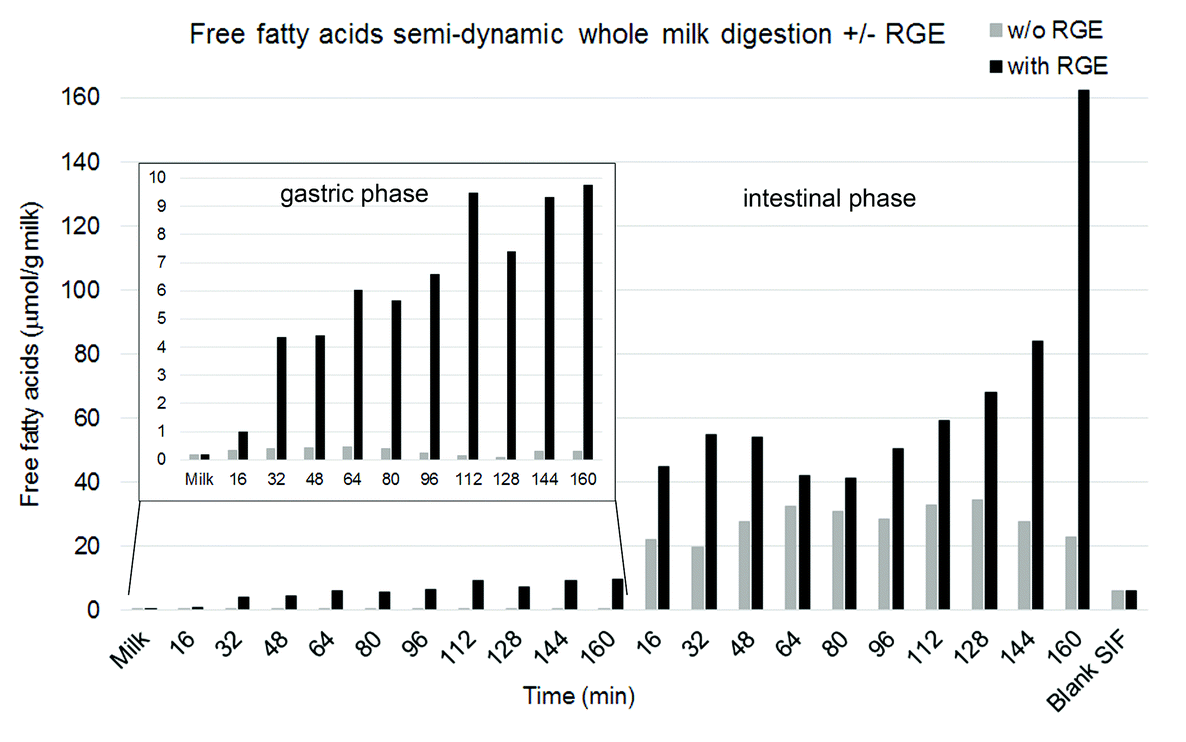 | ||
| Fig. 9 Release of total free fatty acids (FFA, non-esterified fatty acid kit, Fujifilm Wako Diagnostics, US) analysed with a colorimetric method.30 The different gastric emptying time points were individually subjected to intestinal digestion during 120 min. The inlet graph shows the evolution in FFA release during gastric digestion using a smaller scale. The Blank sample shows the FFA content in the intestinal fluid. Data represent one replicate as indicative of the results that can be obtained. | ||
6. Discussion
6.1. Oral phase
The main functions of the oral phase related to food structure are: (a) breaking foods into small pieces by mastication, (b) mixing pieces with secreted saliva for bolus formation and enzyme impregnation. The latter mainly initiates starch hydrolysis by salivary α-amylase.The pH of saliva in resting conditions is often reported to be close to 7. However, pH in the mouth is known to decrease locally due to food fermentation, as represented by the plaque pH.43 This pH usually reaches a minimum 5 to 20 min after the food consumption, and goes back to the resting pH of saliva within 30 to 60 min. As this protocol is intended to follow the passage of food through the GI tract, this effect is not relevant and we recommend a constant pH of 7 for the oral phase.
6.2. Gastric phase
The main functions of the stomach are to provide a reservoir for food and to perform mechanical and chemical disintegration of the contents in order to deliver digested material into the duodenum in a regulated manner. In the stomach, the meal is mixed and digested with gastric secretions including enzymes and hydrochloric acid, ground by antral movements, and gradually emptied into the duodenum. The significance of physiological and simulated gastric parameters will be discussed.The volume of gastric secretion in the fasted state has a high degree of inter individual variability but is typically 4–65 mL.53 In the study by Koziolek et al.,53 the average was actually 31.4 ± 19.7 mL (n = 12). The initial addition of 10% of SFG at low pH in the vessel simulates the basal content in the stomach,54 which lowers the pH of food coming from the mouth. The amount of acid added in the protocol aims to reach a final pH close to 2 and will depend on the properties of the food being studied.
The stomach is a major site of food disintegration and is also responsible of pumping the gastric content into the duodenum in a controlled manner. Gastric motility is characterised by two types of motion. There is a slow motor activity at the upper part of the stomach, fundus and body, by which gastric contents are pushed into the antrum. Therefore, the body part mainly acts as a storage place with negligible mixing. In contrast, the strongest fluid motions are found in the antral part and are responsible for the major food grinding and mixing with gastric fluids.56–58 These peristaltic contraction waves originate in the proximal stomach and their strength increases as they travel toward the pylorus in a sequential manner.57 The pylorus acts as a sieve in which liquid and particles smaller than approximately 3 mm can pass through from the stomach into the duodenum.59,60 Particles greater in size are subjected to a process called retropulsion for further degradation.61
However, the simulation of these mechanical forces is rather difficult due to their complexity in amplitude, frequency and intensity. The present model does not allow an accurate simulation of the gastric motility in particular the mechanical forces of the antrum. We propose a weak mixing during the gastric period simulating the upper part of the stomach, fundus and body, by the use of low frequency orbital shaking or an overhead stirrer using a head of the design outlined in ESI 1.† The latter system may be preferred for more structured gastric content. This allows the mixing of the fluids added with the food, in particular to disperse the acid solution in the vessel. We do not recommend the use of a magnetic stirrer inside the vessel since the type of shear would be different from the stomach physiology and this might disrupt the initial matrix or/and any possible structure formed during digestion. For instance, studies have shown phase separation in the stomach, highlighting the low mixing in the body62,63 contradicting the idea of intragastric homogenisation of the food. In this regard, the present protocol is realistic as it leads to a poor mixing of the gastric contents.
The gastric emptying rate is affected by different physicochemical properties of a meal. It has been shown that high viscosity, osmolarity and, high lipid content delay the gastric emptying.64–66 Large volumes have been seen to increase the rate of emptying and its duration (in the case that there is much more volume to empty).67,68 However, the caloric density of the food has been seen as the major factor controlling gastric emptying, a high caloric density inducing a slower/longer gastric emptying (again in the case that there are more calories to empty).68–71 For example, an increasing caloric density of the food results in a decrease in gastric emptying rate in terms of mL min−1, but an increase in term of kcal min−1.72 In addition, increasing meal volume can also increase the gastric emptying rate but in this case we assume a fixed food volume of 500 mL. A linear relationship has been mostly reported between the caloric content and the intraduodenal calorie delivery rate,69,71,73 in a range from 0.5 to 8 kcal min−1, with a mean value around 2 kcal min−1 (but this depends on the time range over which this rate is calculated and the physical state of the gastric content).69,71,73 The delivery of nutrients initiates feedback mechanisms mediated by duodenal receptors which control the gastric volume through the propulsive forces and pyloric opening to provide regulated intraduodenal nutrient loads.74
The physical state of meal (solid or liquid) has been shown to influence the gastric emptying behaviour.16,75 The emptying from the stomach is slower for solids compared to liquids. Liquid foods empty from the stomach following first order exponential kinetics.16,70,75,76 The emptying behaviour for solids has been shown to be more complicated and is still controversial. Some studies showed a biphasic emptying behaviour of semi-solid foods, where the solids empty linearly after a lag period (zero emptying).54,70,75,77 This lag phase can be explained by the action of the antrum to achieve a suitable particle size, during which no duodenum activity has been seen. The variations among studies of type, caloric content and weight of solid, antral motility as well as the methodology used for gastric emptying measurement, make it difficult to study the lag phase. Also, the different colloidal behaviour in the stomach has been shown to induce changes in emptying profiles.63,78
A simple model cannot take all these variables into account. Thus, we recommend the use of a linear rate of emptying regardless of the physical state of the sample which seems a good approximation due to the ability of the antrum and pylorus to maintain a relatively constant emptied volume. The rates of emptying of the three major macronutrients (lipid, carbohydrate and protein) are indeed regulated so that equal numbers of calories are delivered to the duodenum with an average rate of 2 kcal min−1.71
There is some good evidence showing the relevant contribution of the gastric lipase in the overall digestive process. For this reason, the inclusion of gastric lipase is suggested even though pancreatic lipase is the main enzyme responsible for the digestion of triglycerides. Gastric lipolysis accounts for 10–25% of the overall lipid digestion7,79–81 mainly by hydrolysing triglycerides into diglycerides and fatty acids and contributes up to 7.5% to the duodenal lipolysis.7 Also, gastric lipase is responsible to enhance the pancreatic lipase action.6 Human gastric lipase (HGL) concentration has been seen to increase in the gastric content as function of the time and gastric emptying, with lipase activities ranging from 10 U mL−1 after meal intake to 120 U mL−1 at the end of the meal gastric emptying.7 Since 120 U mL−1 corresponds to the gastric lipase activity in gastric juice,7 we suggest using this activity level in SGF. It is worth noticing that the same activity has been proposed for the new version of the INFOGEST static model.23 The pH for optimal gastric lipase activity has been described in the range from 4 to 5.4.8,10 Since obtaining HGL is limited and subjected to ethical restrictions, we suggest the use of RGL which has been found to have similar activities and specificities to those of HGL.82,83 A recent review compared gastric lipases to microbial lipases and showed the unique characteristics of human and animal gastric lipases.8 Therefore, in case of inaccessibility, we advise not to include any other alternative source of lipase such as microbial lipases which exhibit different activities and specificities.84
With regards to salivary amylase, it should be noticed that this enzyme remains active until gastric pH reaches ∼3.5.11,85,86 Data from human studies showing that salivary amylase can remain active in the stomach long after the oral phase, and might even reach the small intestine without becoming inactive, indicate that this enzyme can be responsible for hydrolysing an important fraction of starch.87 As summarized by Davenport,88 a number of studies around the beginning of the 20th century indeed reached that conclusion. The exact contribution of salivary amylase to starch digestion at the gastric level remain controversial but recent in vitro studies reported that up to 70–80% of bread starch and 15–30% of pasta starch can be hydrolysed, with about half thereof into the form of small oligosaccharides.11,89 This correlates well with the human study of Bergeim90 who reported that up to 76% of the starch in mashed potatoes, and 59% of that in bread, was extensively hydrolysed by salivary amylase in the stomach. The semi-dynamic protocol proposed in here might therefore be useful to gain knowledge on this phenomenon.
6.3. Small intestinal phase
Absorption of most nutrients and micronutrients takes place in the duodenum and jejunum, although some micronutrients are only absorbed in the ileum.93 The physiological conditions we want to model should thus be relevant to the whole small intestine. The pH is known to vary from about 7 to between 6 and 5 in the duodenum due to acidic gastric emptying.13 In contrast, it is more stable in the jejunum and ileum, with a value around 7 in healthy adults.94,95 As the duodenum length is small compared to the other parts of the small intestine, we recommend a constant pH of 7 for the intestinal phase. Concerning the transit time, it should represent the passage through the whole small intestine. This has been characterized in many studies as the small intestine (or small bowel) transit time, with an average value of 240 ± 60 min.96 It is suggested that the duration of the intestinal phase is 2 hours, as recommended in the static INFOGEST protocol,23 with a maximal duration of 4 hours to study specific cases of lower intestinal absorption (ileum).6.4. Differences from the static model
As stated above, the oral and intestinal phase remains essentially the same with the same static approach. However, the chyme emptied from the gastric phase is digested separately in parallel. The rationale for this approach as opposed to simply passing gastric samples into a single intestinal vessel is that segmentation within the small intestine means that the content can in some senses be treated as a plug flow97 and thus digested separately from samples emptied at other time points.The static protocol of the COST Action INFOGEST has been seen as useful for the assessment of the end values but it cannot provide data about the kinetics of nutrient digestion. This has been overcome in the present model by the inclusion of the main dynamics of gastric physiology including gradual pH decrease, gastric fluid secretion and gastric emptying. The kinetics of nutrient digestion are very much linked with the possible structural changes of the food matrix in the gastric compartment, which can be simulated using this semi-dynamic model. For instance, caseins have been denoted as slow proteins in relation to the amino acid absorption, which was suggested to be due to the formation of solid coagula in the stomach. This coagulation can be achieved using the semi-dynamic model, which was not possible with the static model.
6.5. Advantages and limitations of model
The main advantage of this semi-dynamic protocol is the simulation of the transient nature of gastric secretions, gradual acidification and gastric emptying. This allows the evaluation of the changes occurring in food structure and disintegration. For instance, this semi-dynamic setup allows photographic/video record of digesta appearance and visual observation of the changes during gastric digestion at macroscopic level. In addition, the dynamic pH profile can be recorded in situ as well as at each gastric emptying point. Different physicochemical parameters might be assessed in the emptied aliquots to follow the digestion kinetics of nutrients. These analyses include particle size, microscopy and extent of protein, starch and lipid hydrolysis among others.In this model, gastric emptying is simplified and based on the caloric content of food to be digested and set at 2 kcal min−1. This parameter has been found as one of the major factors controlling gastric emptying and is easy to calculate by knowing the composition of protein, lipid and carbohydrate of the food. The provided spreadsheet (see ESI 3†) helps with calculations of this parameter. In addition, the model takes into account the particles that pass through the pylorus by sampling with a tube of a diameter end of ∼3 mm. For those foods that coagulate during gastric residence, like casein, and this results in a delayed gastric emptying, the limited size of the emptying simulates the sieving effect of the pylorus.
Therefore, the semi-dynamic gastric model can be valuable for the assessment of the rate and extent of nutrient bioaccessibility which might give indication of the level of nutrient bioavailability in a more accurate way than static protocols. As occurs for static protocols, this semi-dynamic setup can be used with different volumes and therefore does not consume high amounts of expensive enzymes. Similarly, it is easy to operate and does not require expensive or sophisticated equipment. These advantages make it suitable for a wide range of foods and it can be easily applied in laboratories across the world.
However, as with any in vitro protocol, this semi-dynamic setup has also some limitations. Some parameters, such as, the amount of food, number of gastric emptying aliquots, mixing speed, have to be fixed for each digestion assay which could bring variability between different laboratories. Some dynamic gastric models, such as the Human Gastric Simulator, use more physiologically relevant mechanical forces that simulates the contractions waves of the stomach that might provide more physiologically relevant results in terms of the disintegration of food in the stomach, in particular for complex structured foods.98 Nevertheless, Mulet-Cabero, et al.,19 using the semi-dynamic model in processed milks, obtained similar results to those of Ye, et al.99 using the Human Gastric Simulator in terms of coagulation behaviour, timing, consistency of the gastric digesta and protein digestion.
Another critical point of variability is the manual and stepwise emptying. It is important to note there could be losses of the sample through the emptying process, which will largely depend on the complexity of the studied sample, and the properties of any structures that they form. In that case, it might be advisable to consider the weight of the initial food and the collected aliquots instead of the volume for further calculations. Nevertheless, previous studies using this semi-dynamic model showed suitable reproducibility considering the complexity of structures tested.19,29 Mulet-Cabero, et al.19 quantified the nutrient content in terms of proteins and lipid in each of the five gastric emptying points taken during the gastric digestion of the six milk samples differing in dairy treatment. The average of coefficient of variation was 17.5% and 28.3% for the protein and lipid data respectively.
Oral and intestinal phases are performed static, and it is recommended that the intestinal phase is performed individually with each gastric emptying aliquot. This contrasts with the semi-dynamic two step gastrointestinal model used by Luiking, et al.,100 which consists of computer-controlled reactors. However, this uses more complex and expensive equipment that is less available for most laboratories. It is important to highlight that the semi-dynamic model is a simple model and will never be as consistent as this type of dynamic computerised versions. Moreover, it remains to be elucidated if significant differences will be found at the end of the intestinal phase starting from different gastric points. Preliminary results for SMP digestion have shown that the peptide patterns were similar for all gastric endpoints after the completion of the intestinal phase (Fig. 7). These differences can vary with the food or the nutrient considered and therefore, the application of this semi-dynamic model to different food matrices merits further research.
Author contributions
All authors contributed in the harmonisation of the model. A-I.M-C, LE, OM, SF, AS, M.M.L.G, IR and AM wrote the manuscript. A-I.M-C, LE and RP carried out experiments and analysed data. AB did the 3D files. SM, FC, DD and AM critically revised manuscript.Conflicts of interest
There are no conflicts to declare.Acknowledgements
The authors would like to acknowledge the INFOGEST COST Action FA1005 and subsequently the INFOGEST network funded by INRA for enabling the networking required to develop the model described in this article.References
- K. Venema, R. Havenaar and M. Minekus, Improving in vitro simulation of the stomach and intestines, in Designing Functional Foods: Measuring and Controlling Food Structure Breakdown and Nutrient Absorption, ed. D. J. McClements, E. A. Decker, Woodhead Publishing, Cambridge, UK, 2009, pp. 314–339 Search PubMed.
- A. Guerra, L. Etienne-Mesmin, V. Livrelli, S. Denis, S. Blanquet-Diot and M. Alric, Relevance and challenges in modeling human gastric and small intestinal digestion, Trends Biotechnol., 2012, 30(11), 591–600 CrossRef CAS PubMed.
- M. Minekus, M. Alminger, P. Alvito, S. Ballance, T. Bohn, C. Bourlieu, F. Carriere, R. Boutrou, M. Corredig, D. Dupont, C. Dufour, L. Egger, M. Golding, S. Karakaya, B. Kirkhus, S. Le Feunteun, U. Lesmes, A. Macierzanka, A. Mackie, S. Marze, D. J. McClements, O. Menard, I. Recio, C. N. Santos, R. P. Singh, G. E. Vegarud, M. S. Wickham, W. Weitschies and A. Brodkorb, A standardised static in vitro digestion method suitable for food - an international consensus, Food Funct., 2014, 5(6), 1113–1124 RSC.
- A. Brodkorb, L. Egger, M. Alminger, P. Alvito, R. Assuncao, S. Ballance, T. Bohn, C. Bourlieu-Lacanal, R. Boutrou, F. Carriere, A. Clemente, M. Corredig, D. Dupont, C. Dufour, C. Edwards, M. Golding, S. Karakaya, B. Kirkhus, S. Le Feunteun, U. Lesmes, A. Macierzanka, A. R. Mackie, C. Martins, S. Marze, D. J. McClements, O. Menard, M. Minekus, R. Portmann, C. N. Santos, I. Souchon, R. P. Singh, G. E. Vegarud, M. S. J. Wickham, W. Weitschies and I. Recio, INFOGEST static in vitro simulation of gastrointestinal food digestion, Nat. Protoc., 2019, 14(4), 991–1014 CrossRef CAS PubMed.
- J. C. Bakala N'Goma, S. Amara, K. Dridi, V. Jannin and F. Carriere, Understanding the lipid-digestion processes in the GI tract before designing lipid-based drug-delivery systems, Ther. Delivery, 2012, 3(1), 105–124 CrossRef PubMed.
- M. Armand, Lipases and lipolysis in the human digestive tract: where do we stand?, Curr. Opin. Clin. Nutr. Metab. Care, 2007, 10(2), 156–164 CrossRef CAS PubMed.
- F. Carriere, J. A. Barrowman, R. Verger and L. René, Secretion and contribution to lipolysis of gastric and pancreatic lipases during a test meal in humans, Gastroenterology, 1993, 105(3), 876–888 CrossRef CAS.
- L. Sams, J. Paume, J. Giallo and F. Carriere, Relevant pH and lipase for in vitro models of gastric digestion, Food Funct., 2016, 7(1), 30–45 RSC.
- D. W. Piper and B. H. Fenton, pH stability and activity curves of pepsin with special reference to their clinical importance, Gut, 1965, 6(5), 506–508 CrossRef CAS PubMed.
- F. Carriere, H. Moreau, V. Raphel, R. Laugier, C. Benicourt, J. L. Junien and R. Verger, Purification and biochemical characterization of dog gastric lipase, Eur. J. Biochem., 1991, 202(1), 75–83 CrossRef CAS PubMed.
- D. Freitas, S. Le Feunteun, M. Panouillé and I. Souchon, The important role of salivary α-amylase in the gastric digestion of wheat bread starch, Food Funct., 2018, 9(1), 200–208 RSC.
- B. L. Dekkers, E. Kolodziejczyk, S. Acquistapace, J. Engmann and T. J. Wooster, Impact of gastric pH profiles on the proteolytic digestion of mixed betalg-Xanthan biopolymer gels, Food Funct., 2016, 7(1), 58–68 RSC.
- J. B. Dressman, R. R. Berardi, L. C. Dermentzoglou, T. L. Russell, S. P. Schmaltz, J. L. Barnett and K. M. Jarvenpaa, Upper gastrointestinal (GI) pH in young, healthy men and women, Pharm. Res., 1990, 7(7), 756–761 CrossRef CAS PubMed.
- L. Kalantzi, K. Goumas, V. Kalioras, B. Abrahamsson, J. B. Dressman and C. Reppas, Characterization of the human upper gastrointestinal contents under conditions simulating bioavailability/bioequivalence studies, Pharm. Res., 2006, 23(1), 165–176 CrossRef CAS PubMed.
- H. Singh and A. Sarkar, Behaviour of protein-stabilised emulsions under various physiological conditions, Adv. Colloid Interface Sci., 2011, 165(1), 47–57 CrossRef CAS PubMed.
- J. R. Malagelada, V. L. Go and W. H. Summerskill, Different gastric, pancreatic, and biliary responses to solid-liquid or homogenized meals, Dig. Dis. Sci., 1979, 24(2), 101–110 CrossRef CAS PubMed.
- A. Sarkar, K. K. T. Goh, R. P. Singh and H. Singh, Behaviour of an oil-in-water emulsion stabilized by β-lactoglobulin in an in vitro gastric model, Food Hydrocolloids, 2009, 23(6), 1563–1569 CrossRef CAS.
- Y. Boirie, M. Dangin, P. Gachon, M.-P. Vasson, J.-L. Maubois and B. Beaufrère, Slow and fast dietary proteins differently modulate postprandial protein accretion, Proc. Natl. Acad. Sci. U. S. A., 1997, 94(26), 14930–14935 CrossRef CAS PubMed.
- A.-I. Mulet-Cabero, A. R. Mackie, P. J. Wilde, M. A. Fenelon and A. Brodkorb, Structural mechanism and kinetics of in vitro gastric digestion are affected by process-induced changes in bovine milk, Food Hydrocolloids, 2019, 86, 172–183 CrossRef CAS.
- O. Menard, T. Cattenoz, H. Guillemin, I. Souchon, A. Deglaire, D. Dupont and D. Picque, Validation of a new in vitro dynamic system to simulate infant digestion, Food Chem., 2014, 145, 1039–1045 CrossRef CAS PubMed.
- M. Minekus, M. Smeets-Peeters, A. Bernalier, S. Marol-Bonnin, R. Havenaar, P. Marteau, M. Alric, G. Fonty and J. H. Huis in't Veld, A computer-controlled system to simulate conditions of the large intestine with peristaltic mixing, water absorption and absorption of fermentation products, Appl. Microbiol. Biotechnol., 1999, 53(1), 108–114 CrossRef CAS PubMed.
- M. Wickham, R. Faulks and C. Mills, In vitro digestion methods for assessing the effect of food structure on allergen breakdown, Mol. Nutr. Food Res., 2009, 53(8), 952–958 CrossRef CAS PubMed.
- A. Brodkorb, L. Egger, M. Alminger, P. Alvito, R. Assunção, S. Ballance, T. Bohn, C. Bourlieu-Lacanal, R. Boutrou, F. Carrière, A. Clemente, M. Corredig, D. Dupont, C. Dufour, C. Edwards, M. Golding, S. Karakaya, B. Kirkhus, S. Le Feunteun, U. Lesmes, A. Macierzanka, A. R. Mackie, C. Matins, S. Marze, D. J. McClements, O. Ménard, M. Minekus, R. Portmann, C. N. Santos, I. Souchon, R. P. Singh, G. E. Vegarud, M. S. J. Wickham, W. Weitschies and I. Recio, INFOGEST static in vitro simulation of gastrointestinal food digestion, Nat. Protoc., 2019, 14, 991–1014 CrossRef CAS PubMed.
- L. Sams, S. Amara, P. Mansuelle, R. Puppo, R. Lebrun, J. Paume, J. Giallo and F. Carrière, Characterization of pepsin from rabbit gastric extract, its action on β-casein and the effects of lipids on proteolysis, Food Funct., 2018, 9(11), 5975–5988 RSC.
- L. Humbert, M. A. Maubert, C. Wolf, H. Duboc, M. Mahé, D. Farabos, P. Seksik, J. M. Mallet, G. Trugnan and J. Masliah, Bile acid profiling in human biological samples: comparison of extraction procedures and application to normal and cholestatic patients, J. Chromatogr. B: Anal. Technol. Biomed. Life Sci., 2012, 899, 135–145 CrossRef CAS PubMed.
- F. Bellisle and J. Le Magnen, The structure of meals in humans: eating and drinking patterns in lean and obese subjects, Physiol. Behav., 1981, 27(4), 649–658 CrossRef CAS PubMed.
- F. Carriere, J. A. Barrowman, R. Verger and R. Laugier, Secretion and contribution to lipolysis of gastric and pancreatic lipases during a test meal in humans, Gastroenterology, 1993, 105(3), 876–888 CrossRef CAS.
- I. Goñi, A. Garcia-Alonso and F. Saura-Calixto, A starch hydrolysis procedure to estimate glycemic index, Nutr. Res., 1997, 17(3), 427–437 CrossRef.
- A.-I. Mulet-Cabero, N. M. Rigby, A. Brodkorb and A. R. Mackie, Dairy food structures influence the rates of nutrient digestion through different in vitro gastric behaviour, Food Hydrocolloids, 2017, 67, 63–73 CrossRef CAS.
- L. Egger, et al., Digestion of milk proteins: Comparing static and dynamic in vitro digestion systems with in vivo data, Food Res. Int., 2019, 118, 32–39 CrossRef CAS PubMed.
- W. Duncombe, The colorimetric micro-determination of non-esterified fatty acids in plasma, Clin. Chim. Acta, 1964, 9, 122–125 CrossRef CAS.
- Q. Guo, A. Ye, M. Lad, D. Dalgleish and H. Singh, The breakdown properties of heat-set whey protein emulsion gels in the human mouth, Food Hydrocolloids, 2013, 33(2), 215–224 CrossRef CAS.
- C. Hoebler, M. F. Devaux, A. Karinthi, C. Belleville and J. L. Barry, Particle size of solid food after human mastication and in vitro simulation of oral breakdown, Int. J. Food Sci. Nutr., 2000, 51(5), 353–366 CrossRef CAS PubMed.
- M.-L. Jalabert-Malbos, A. Mishellany-Dutour, A. Woda and M.-A. Peyron, Particle size distribution in the food bolus after mastication of natural foods, Food Qual. Prefer., 2007, 18(5), 803–812 CrossRef.
- F. Le Bleis, L. Chaunier, P. Montigaud and G. Della Valle, Destructuration mechanisms of bread enriched with fibers during mastication, Food Res. Int., 2016, 80, 1–11 CrossRef.
- M. M. Grundy, T. Grassby, G. Mandalari, K. W. Waldron, P. J. Butterworth, S. E. Berry and P. R. Ellis, Effect of mastication on lipid bioaccessibility of almonds in a randomized human study and its implications for digestion kinetics, metabolizable energy, and postprandial lipemia, Am. J. Clin. Nutr., 2014, 101(1), 25–33 CrossRef PubMed.
- C. Hoebler, A. Karinthi, M. F. Devaux, F. Guillon, D. J. Gallant, B. Bouchet, C. Melegari and J. L. Barry, Physical and chemical transformations of cereal food during oral digestion in human subjects, Br. J. Nutr., 1998, 80(5), 429–436 CrossRef CAS PubMed.
- K. D. Foster, A. Woda and M. A. Peyron, Effect of texture of plastic and elastic model foods on the parameters of mastication, J. Neurophysiol., 2006, 95(6), 3469–3479 CrossRef CAS PubMed.
- K. Kohyama and L. Mioche, Chewing behavior observed at different stages of mastication for six foods, studied by electromyography and jaw kinematics in young and elderly subjects, J. Texture Stud., 2004, 35(4), 395–414 CrossRef.
- L. Laguna and A. Sarkar, Influence of mixed gel structuring with different degrees of matrix inhomogeneity on oral residence time, Food Hydrocolloids, 2016, 61, 286–299 CrossRef CAS.
- M. A. Peyron, C. Lassauzay and A. Woda, Effects of increased hardness on jaw movement and muscle activity during chewing of visco-elastic model foods, Exp. Brain Res., 2002, 142(1), 41–51 CrossRef CAS PubMed.
- S. R. Drago, M. Panouillé, A. Saint-Eve, E. Neyraud, G. Feron and I. Souchon, Relationships between saliva and food bolus properties from model dairy products, Food Hydrocolloids, 2011, 25(4), 659–667 CrossRef CAS.
- I. Kleinberg and G. N. Jenkins, The pH of dental plaques in the different areas of the mouth before and after meals and their relationship to the pH and rate of flow of restin saliva, Arch. Oral Biol., 1964, 9, 493–516 CrossRef CAS PubMed.
- C. Tournier, M. Grass, C. Septier, D. Bertrand and C. Salles, The impact of mastication, salivation and food bolus formation on salt release during bread consumption, Food Funct., 2014, 5(11), 2969–2980 RSC.
- S. Watanabe and C. Dawes, The effects of different foods and concentrations of citric acid on the flow rate of whole saliva in man, Arch. Oral Biol., 1988, 33(1), 1–5 CrossRef CAS PubMed.
- G. M. Bornhorst and R. P. Singh, Bolus formation and disintegration during digestion of food carbohydrates, Compr. Rev. Food Sci. Food Saf., 2012, 11(2), 101–118 CrossRef CAS.
- E. Neyraud, J. Bult and E. Dransfield, Continuous analysis of parotid saliva during resting and short-duration simulated chewing, Arch. Oral Biol., 2009, 54(5), 449–456 CrossRef CAS PubMed.
- C. Méjean, M. Morzel, E. Neyraud, S. Issanchou, C. Martin, S. Bozonnet, C. Urbano, P. Schlich, S. Hercberg and S. Péneau, Salivary composition is associated with liking and usual nutrient intake, PLoS One, 2015, 10(9), e0137473 CrossRef PubMed.
- G. K. Aji, F. J. Warren and E. Roura, Salivary α-Amylase Activity and Starch-Related Sweet Taste Perception in Humans, Chem. Senses, 2019, 44(4), 249–256 CrossRef PubMed.
- L. Marciani, P. A. Gowland, R. C. Spiller, P. Manoj, R. J. Moore, P. Young and A. J. Fillery-Travis, Effect of meal viscosity and nutrients on satiety, intragastric dilution, and emptying assessed by MRI, Am. J. Physiol.: Gastrointest. Liver Physiol., 2001, 280(6), G1227–G1233 CrossRef CAS PubMed.
- D. D. Burton, H. J. Kim, M. Camilleri, D. A. Stephens, B. P. Mullan, M. K. O'Connor and N. J. Talley, Relationship of gastric emptying and volume changes after a solid meal in humans, Am. J. Physiol.: Gastrointest. Liver Physiol., 2005, 289(2), G261–G266 CrossRef CAS PubMed.
- O. Wisen and C. Johansson, Gastrointestinal function in obesity: motility, secretion, and absorption following a liquid test meal, Metabolism, 1992, 41(4), 390–395 CrossRef CAS PubMed.
- M. Koziolek, M. Grimm, G. Garbacz, J. P. Kuhn and W. Weitschies, Intragastric volume changes after intake of a high-caloric, high-fat standard breakfast in healthy human subjects investigated by MRI, Mol. Pharm., 2014, 11(5), 1632–1639 CrossRef CAS PubMed.
- J. R. Malagelada, G. F. Longstreth, W. H. Summerskill and V. L. Go, Measurement of gastric functions during digestion of ordinary solid meals in man, Gastroenterology, 1976, 70(2), 203–210 CrossRef CAS.
- V. Kitty, C. Paul, L.-E. Iván, K. Charlotte, L. Tor, M. Alan, R. Teresa, S. Dominika and W. Harry, The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models, 2015, p. 338 Search PubMed.
- M. J. Ferrua and R. P. Singh, Modeling the fluid dynamics in a human stomach to gain insight of food digestion, J. Food Sci., 2010, 75(7), R151–R162 CrossRef CAS PubMed.
- R. Berry, T. Miyagawa, N. Paskaranandavadivel, P. Du, T. R. Angeli, M. L. Trew, J. A. Windsor, Y. Imai, G. O'Grady and L. K. Cheng, Functional physiology of the human terminal antrum defined by high-resolution electrical mapping and computational modeling, Am. J. Physiol.: Gastrointest. Liver Physiol., 2016, 311(5), G895–G902 CrossRef PubMed.
- S. M. Harrison, P. W. Cleary and M. D. Sinnott, Investigating mixing and emptying for aqueous liquid content from the stomach using a coupled biomechanical-SPH model, Food Funct., 2018, 9(6), 3202–3219 RSC.
- J. H. Meyer, J. Elashoff, V. Porter-Fink, J. Dressman and G. L. Amidon, Human postprandial gastric emptying of 1–3-millimeter spheres, Gastroenterology, 1988, 94(6), 1315–1325 CrossRef CAS.
- J. K. Rhie, Y. Hayashi, L. S. Welage, J. Frens, R. J. Wald, J. L. Barnett, G. E. Amidon, L. Putcha and G. L. Amidon, Drug marker absorption in relation to pellet size, gastric motility and viscous meals in humans, Pharm. Res., 1998, 15(2), 233–238 CrossRef CAS PubMed.
- F. Kong and R. P. Singh, Disintegration of solid foods in human stomach, J. Food Sci., 2008, 73(5), R67–R80 CrossRef CAS PubMed.
- A. R. Mackie, H. Rafiee, P. Malcolm, L. Salt and G. van Aken, Specific food structures supress appetite through reduced gastric emptying rate, Am. J. Physiol.: Gastrointest. Liver Physiol., 2013, 304(11), G1038–G1043 CrossRef CAS PubMed.
- L. Marciani, M. Wickham, G. Singh, D. Bush, B. Pick, E. Cox, A. Fillery-Travis, R. Faulks, C. Marsden, P. A. Gowland and R. C. Spiller, Enhancement of intragastric acid stability of a fat emulsion meal delays gastric emptying and increases cholecystokinin release and gallbladder contraction, Am. J. Physiol.: Gastrointest. Liver Physiol., 2007, 292(6), G1607–G1613 CrossRef CAS PubMed.
- B. Flourie, N. Vidon, J. A. Chayvialle, R. Palma, C. Franchisseur and J. J. Bernier, Effect of increased amounts of pectin on a solid-liquid meal digestion in healthy man, Am. J. Clin. Nutr., 1985, 42(3), 495–503 CrossRef CAS PubMed.
- S. C. Larsson, L. Bergkvist and A. Wolk, High-fat dairy food and conjugated linoleic acid intakes in relation to colorectal cancer incidence in the Swedish Mammography Cohort, Am. J. Clin. Nutr., 2005, 82(4), 894–900 CrossRef CAS PubMed.
- N. W. Read, J. Cammack, C. Edwards, A. M. Holgate, P. A. Cann and C. Brown, Is the transit time of a meal through the small intestine related to the rate at which it leaves the stomach?, Gut, 1982, 23(10), 824–828 CrossRef CAS PubMed.
- J. B. Mitchell and K. W. Voss, The influence of volume on gastric emptying and fluid balance during prolonged exercise, Med. Sci. Sports Exercise, 1991, 23(3), 314–319 CrossRef CAS.
- T. H. Moran, J. B. Wirth, G. J. Schwartz and P. R. McHugh, Interactions between gastric volume and duodenal nutrients in the control of liquid gastric emptying, Am. J. Physiol.: Regul., Integr. Comp. Physiol., 1999, 276(4), R997–R1002 CrossRef CAS PubMed.
- J. A. Calbet and D. A. MacLean, Role of caloric content on gastric emptying in humans, J. Physiol., 1997, 498(Pt 2), 553–559 CrossRef CAS PubMed.
- P. J. Collins, M. Horowitz, D. J. Cook, P. E. Harding and D. J. Shearman, Gastric emptying in normal subjects–a reproducible technique using a single scintillation camera and computer system, Gut, 1983, 24(12), 1117–1125 CrossRef CAS PubMed.
- J. N. Hunt and D. F. Stubbs, The volume and energy content of meals as determinants of gastric emptying, J. Physiol., 1975, 245(1), 209–225 CrossRef CAS PubMed.
- J. N. Hunt, J. L. Smith and C. L. Jiang, Effect of meal volume and energy density on the gastric emptying of carbohydrates, Gastroenterology, 1985, 89(6), 1326–1330 CrossRef CAS.
- J. G. Moore, P. E. Christian and R. E. Coleman, Gastric emptying of varying meal weight and composition in man. Evaluation by dual liquid- and solid-phase isotopic method, Dig. Dis. Sci., 1981, 26(1), 16–22 CrossRef CAS PubMed.
- P. Janssen, P. Vanden Berghe, S. Verschueren, A. Lehmann, I. Depoortere and J. Tack, Review article: the role of gastric motility in the control of food intake, Aliment. Pharmacol. Ther., 2011, 33(8), 880–894 CrossRef CAS PubMed.
- M. Camilleri, J. R. Malagelada, M. L. Brown, G. Becker and A. R. Zinsmeister, Relation between antral motility and gastric emptying of solids and liquids in humans, Am. J. Physiol., 1985, 249(5 Pt 1), G580–G585 CAS.
- P. M. Hellstrom, P. Gryback and H. Jacobsson, The physiology of gastric emptying, Best Pract. Res., Clin. Anaesthesiol., 2006, 20(3), 397–407 CrossRef PubMed.
- J. A. Siegel, J. L. Urbain, L. P. Adler, N. D. Charkes, A. H. Maurer, B. Krevsky, L. C. Knight, R. S. Fisher and L. S. Malmud, Biphasic nature of gastric emptying, Gut, 1988, 29(1), 85–89 CrossRef CAS PubMed.
- A. Steingoetter, T. Radovic, S. Buetikofer, J. Curcic, D. Menne, M. Fried, W. Schwizer and T. J. Wooster, Imaging gastric structuring of lipid emulsions and its effect on gastrointestinal function: a randomized trial in healthy subjects, Am. J. Clin. Nutr., 2015, 101(4), 714–724 CrossRef CAS PubMed.
- M. Armand, B. Pasquier, M. Andre, P. Borel, M. Senft, J. Peyrot, J. Salducci, H. Portugal, V. Jaussan and D. Lairon, Digestion and absorption of 2 fat emulsions with different droplet sizes in the human digestive tract, Am. J. Clin. Nutr., 1999, 70(6), 1096–1106 CrossRef CAS PubMed.
- M. Armand, B. Pasquier, M. André, M. Senft, J. Peyrot, J. Salducci and D. Lairon, Émulsions et absorption des lipides : importance des propriétés physico-chimiques, 1997, vol. 4, pp. 178–185 Search PubMed.
- F. Carrière, C. Renou, S. Ransac, V. Lopez, J. De Caro, F. Ferrato, A. De Caro, A. Fleury, P. Sanwald-Ducray and H. Lengsfeld, Inhibition of gastrointestinal lipolysis by Orlistat during digestion of test meals in healthy volunteers, Am. J. Physiol.: Gastrointest. Liver Physiol., 2001, 281(1), G16–G28 CrossRef PubMed.
- H. Moreau, Y. Gargouri, D. Lecat, J.-L. Junien and R. Verger, Purification, characterization and kinetic properties of the rabbit gastric lipase, Biochim. Biophys. Acta, Lipids Lipid Metab., 1988, 960(3), 286–293 CrossRef CAS.
- P. Capolino, C. Guérin, J. Paume, J. Giallo, J.-M. Ballester, J.-F. Cavalier and F. Carrière, In Vitro Gastrointestinal Lipolysis: Replacement of Human Digestive Lipases by a Combination of Rabbit Gastric and Porcine Pancreatic Extracts, Food Dig., 2011, 2(1), 43–51 CrossRef CAS.
- P. L. Zentler-Munro, B. A. Assoufi, K. Balasubramanian, S. Cornell, D. Benoliel, T. C. Northfield and M. E. Hodson, Therapeutic potential and clinical efficacy of acid-resistant fungal lipase in the treatment of pancreatic steatorrhoea due to cystic fibrosis, Pancreas, 1992, 7(3), 311–319 CrossRef CAS PubMed.
- P. Bernfeld, A. Staub and E. H. Fischer, Sur les enzymes amylolytiques XI. Propriétés de l’α-amylase de salive humaine cristallisée, Helv. Chim. Acta, 1948, 31(7), 2165–2172 CrossRef CAS PubMed.
- G. J. Walker and W. J. Whelan, The mechanism of carbohydrase action. 7. Stages in the salivary α-amylolysis of amylose, amylopectin and glycogen, Biochem. J., 1960, 76(2), 257 CrossRef CAS PubMed.
- M. Fried, S. Abramson and J. Meyer, Passage of salivary amylase through the stomach in humans, Dig. Dis. Sci., 1987, 32(10), 1097–1103 CrossRef CAS PubMed.
- H. W. Davenport, The gastric mucosal barrier, in A History of Gastric Secretion and Digestion, Springer, 1992, pp. 258–276 Search PubMed.
- D. Freitas and S. Le Feunteun, Oro-gastro-intestinal digestion of starch in white bread, wheat-based and gluten-free pasta: unveiling the contribution of human salivary α-amylase, Food Chem., 2019, 274, 566–573 CrossRef CAS PubMed.
- O. Bergeim, Intestinal chemistry: iii. Salivary digestion in the human stomach and intestines, Arch. Intern. Med., 1926, 37(1), 110–117 CrossRef.
- R. Gotthard, G. Bodemar, M. Tjadermo, P. Tobiasson and A. Walan, High gastric bile acid concentration in prepyloric ulcer patients, Scand. J. Gastroenterol., 1985, 20(4), 439–446 CrossRef CAS PubMed.
- J. Wenner, T. Gunnarsson, H. Graffner and G. Lindell, Influence of smoking and Helicobacter pylori on gastric phospholipids, Dig. Dis. Sci., 2000, 45(8), 1648–1652 CrossRef CAS PubMed.
- L. R. Johnson, in Gastrointestinal Physiology, Elsevier, Mosby, 8th edn, 2013 Search PubMed.
- D. F. Evans, G. Pye, R. Bramley, A. G. Clark, T. J. Dyson and J. D. Hardcastle, Measurement of gastrointestinal pH profiles in normal ambulant human subjects, Gut, 1988, 29(8), 1035–1041 CrossRef CAS PubMed.
- A. E. Mikolajczyk, S. Watson, B. L. Surma and D. T. Rubin, Assessment of Tandem Measurements of pH and Total Gut Transit Time in Healthy Volunteers, Clin. Transl. Gastroenterol., 2015, 6, e100 CrossRef PubMed.
- S. Marze, Modeling of Food Digestion, in Modeling of Microscale Transport in Biological Processes, Elsevier, 2017, pp. 353–374 Search PubMed.
- M. R. Jaime-Fonseca, O. Gouseti, P. J. Fryer, M. S. J. Wickham and S. Bakalis, Digestion of starch in a dynamic small intestinal model, Eur. J. Nutr., 2016, 55(8), 2377–2388 CrossRef CAS PubMed.
- F. Kong and R. P. Singh, A Human Gastric Simulator (HGS) to Study Food Digestion in Human Stomach, J. Food Sci., 2010, 75(9), E627–E635 CrossRef CAS PubMed.
- A. Ye, J. Cui, D. Dalgleish and H. Singh, Effect of homogenization and heat treatment on the behavior of protein and fat globules during gastric digestion of milk, J. Dairy Sci., 2017, 100(1), 36–47 CrossRef CAS PubMed.
- Y. C. Luiking, E. Abrahamse, T. Ludwig, Y. Boirie and S. Verlaan, Protein type and caloric density of protein supplements modulate postprandial amino acid profile through changes in gastrointestinal behaviour: A randomized trial, Clin. Nutr., 2016, 35(1), 48–58 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c9fo01293a |
| This journal is © The Royal Society of Chemistry 2020 |