
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Engineered nanoselenium supplemented fish diet: toxicity comparison with ionic selenium and stability against particle dissolution, aggregation and release†
Fazel
Abdolahpur Monikh
 *a,
Latifeh
Chupani
bc,
Kristyna
Smerkova
de,
Thijs
Bosker
af,
Petr
Cizar
g,
Vladislav
Krzyzanek
h,
Lukas
Richtera
de,
Roman
Franek
c,
Eliska
Zuskova
c,
Radim
Skoupy
h,
Gopala Krishna
Darbha
i,
Martina
Vijver
a,
Eugenia
Valsami-Jones
j and
Willie
Peijnenburg
ak
*a,
Latifeh
Chupani
bc,
Kristyna
Smerkova
de,
Thijs
Bosker
af,
Petr
Cizar
g,
Vladislav
Krzyzanek
h,
Lukas
Richtera
de,
Roman
Franek
c,
Eliska
Zuskova
c,
Radim
Skoupy
h,
Gopala Krishna
Darbha
i,
Martina
Vijver
a,
Eugenia
Valsami-Jones
j and
Willie
Peijnenburg
ak
aInstitute of Environmental Sciences (CML), Van Steenis Building, Leiden University, P.O. Box 9518, Einsteinweg 2, 2333 CC, 2300 RA Leiden, The Netherlands. E-mail: f.a.monikh@cml.leidenuniv.nl
bBiology, Örebro Life Science Center, School of Science and Technology, Örebro University, SE-701 82 Örebro, Sweden
cFaculty of Fisheries and Protection of Waters, South Bohemian Research Centre of Aquaculture and Biodiversity of Hydrocenoses, University of South Bohemia in Ceske Budejovice, Vodňany, Czech Republic
dDepartment of Chemistry and Biochemistry, Mendel University in Brno, Zemedelska 1, CZ-613 00 Brno, Czech Republic
eCentral European Institute of Technology, Brno University of Technology, Purkynova 123, CZ-612 00 Brno, Czech Republic
fLeiden University College The Hague, Leiden University, The Hague, The Netherlands
gInstitute of Complex Systems, University of South Bohemia in Ceske Budejovice, Ceske Budejovice, Czech Republic
hThe Czech Academy of Sciences, Institute of Scientific Instruments, Kralovopolska 147, CZ-612 64 Brno, Czech Republic
iDepartment of Earth Sciences & Centre for Climate and Environmental Studies, Indian Institute of Science Education and Research Kolkata, Mohanpur, West Bengal-741246, India
jSchool of Geography, Earth and Environmental Science, University of Birmingham, Edgbaston, B15 2TT Birmingham, UK
kNational Institute of Public Health and the Environment (RIVM), Center for Safety of substances and Products, Bilthoven, The Netherlands
First published on 19th June 2020
Abstract
Transformation of nutrients to their nano-form, such as selenium (Se) engineered nanonutrients (Se-ENNs), is expected to enhance the absorption of the nutrients into fish and increase the efficiency of the feed. However, dissolution, aggregation, and release of ENNs from the feed matrix may decrease the efficiency of the Se-ENNs. In this study, we provided fish feed supplemented with Se-ENNs which do not aggregate or dissolve and the particles are also not released from the feed matrix. As a proof of principle, we compared the toxicity of a diet containing Se-ENNs of two different sizes (60 nm and 120 nm) with diets containing ionic Se. The adverse effects were measured by monitoring the survival rate, acetylcholinesterase (AChE) levels and swimming behavior of zebrafish over 21 days of feeding with either the Se-ENNs or ionic Se supplemented fish diets. The number size distribution of the 60 nm Se-ENNs in the diet was similar to that in MilliQ water, while the size distribution of the 120 nm Se-ENNs in the diet was slightly wider. Ion and particle release from Se-ENNs containing diets in the exposure media was not observed, indicating the stability of the particles in the feed matrices. To determine toxicity, zebrafish (Danio rerio) were nourished using a control diet (without Se and Se-ENNs), Se (sodium selenite) containing diets (with 2.4 or 240 mg Se per kg feed) and Se-ENNs containing diets (with 2.4 or 240 mg Se-ENNs of 60 or 120 nm per kg feed) for 21 days. Both sizes of Se-ENNs were taken up in the fish, however only the 120 nm Se-ENNs were detected in the brains of fish. Zebrafish fed with Se-ENNs supplemented diets (60 and 120 nm) showed normal swimming behavior compared to the control. No significant alteration was determined in the AChE activity of the fish fed with the Se-ENNs supplemented diet. In contrast, feeding the zebrafish with a diet containing 240 mg kg−1 Se led to lethal effects. These observations clearly depict the potential benefits of using Se-ENNs as nutrients in fish feed.
Environmental significanceNanotechnology offers a novel opportunity for the aquaculture industry to apply nanomaterials as nutrients, so-called engineered nanonutrients (ENNs). For example, the application of selenium (Se)-ENNs in feed has been reported to improve the final weight and antioxidant status of fish. The challenge is that ENNs in the matrices of feed may undergo dissolution, aggregation, and release which decrease the efficiency of the ENNs. Moreover, like the challenge of the application of other nanomaterials in the food industry, the usage of Se-ENNs may be hampered by the unknown toxicity profile of the particles. In this study, we provided a fish diet supplemented with Se-ENNs. The Se-ENNs in the matrix of the feed are stable against dissolution, aggregation, and release over time. Our study for the first time provided a fish diet containing stable Se-ENNs as nutrients with no adverse effects on swimming behavior and on AChE activity in zebrafish, displaying no effects at a dose that is lethal in the case of ionic Se. |
Introduction
Aquaculture is a rapidly growing worldwide industry, which plays an essential role to assure food security by contributing to the total global fish production (47% in 2016).1 Since the late 1980s, wild-caught fishery production reached a relatively steady state, while aquaculture production has continued to grow, thereby keeping pace with the increasing demands for fish consumption.1 Therefore the aquaculture sector holds great promise in supporting FAO's goal of a world without hunger and malnutrition.2 However, the growth of aquaculture is not without various challenges. For instance, to produce fishmeal the aquaculture industry sector is increasingly harvesting oceanic stocks. This practice increases, causing significant pressure on oceanic resources, and potentially hampering the development of a sustainable aquaculture sector.3,4 To this end, there is an urgent need for sustainable and novel feed ingredients to decrease the demand for fishmeal, for example by increasing the absorption and assimilation efficiency of the nutrients in the feed. These novel ingredients could also support the aquaculture industry to contend against nutritional deficiencies in fish caused by the lack, excess or improper balance of (micro)nutrients present in fish feed for example.5Nanotechnology is one potential novel and innovative technology that allows the production and functionalization of nanomaterials and can facilitate the sustainable growth of aquaculture. For example, transforming nutrients to their nano-form, so-called engineered nanonutrients (ENNs), can enhance the absorption of the nutrients into fish, rather than the nutrients passing unused through the fish digestive system,6,7 thereby increasing the uptake efficiency of the nutrients.8 For instance the addition of iron ENNs to a regular fish diet was shown to positively affect the growth of young carp (Labeo rohita), as well as improvements in the hematological and immunological parameters, compared to conventionally added iron.6 Thus, ENNs may boost the efficiency of nutrient uptake and they may be an important resource for a more sustainable aquaculture industry. However, addition of ENNs in feed is a challenging task due to the possible particle dissolution and release from the formulated feeds and/or particle aggregation which can decrease the absorption and efficiency of the ENNs.9
One key nutrient for optimal fish growth is selenium (Se), with fish requiring dietary concentrations of 0.1–0.5 mg of Se per kg of dry weight (dw).10 Importantly, Se has a narrow window of essentiality from being an essential nutrient to a toxic compound.11 A moderate level of this nutrient is essential for immune functioning, for example against oxygen-derived radicals, while an elevated concentration of Se generates oxidative stress in fish.12 When Se levels exceed 3 mg kg−1 dw, storage capacity is overloaded and the excess of Se can cause an adverse response. Toxicity of Se is likely to be related to its prooxidant ability to catalyze the oxidation of thiol groups and prevent the generation of superoxide.13,14 For example, Han et al.,15 reported that exposure to dietary Se significantly decreased the serum superoxide dismutase (SOD) activities of crucian carp (Carassius auratus gibelio).
An increasing number of studies on Se-ENNs show that these ENNs have potential as a nutritional supplement in fish food.16–21 For example, a basal diet supplemented with Se-ENNs improved the final weight, relative weight gain rate, antioxidant status, and muscle Se concentration of crucian carp and common carp (Cyprinus carpio).17,22 Dietary uptake of Se-ENNs also improved the systemic and mucosal immune status as well as stress resistance of red sea bream (Pagrus major).18 However, the previous studies aimed to monitor only the benefit of supplementing Se-ENNs to fish diet without considering the time-resolved stability of the Se-ENNs in the feed with regard to particle aggregation, release and dissolution, as well as the possible adverse effects of the Se-ENNs to target fish. The adverse effects of Se-ENNs are still unknown. This restricts the application of these materials in feed. In addition, there are generic challenges surrounding the application of nanomaterials as a whole in the food industry, mostly due to public fears about their safety, and a feeling that the use of nanomaterials is tampering with nature.23 Amongst others, Se-ENNs may induce a different mode of biodistribution in comparison to ionic Se based on the nano-specific properties of the particles such as small size and large surface area.24 Therefore, it is possible that Se-ENNs reach targets inside fish that cannot be reached by ionic Se. To the best of our knowledge, no study is available that has quantified the bioaccumulation of Se-ENNs in particulate form in fish tissues.
The main objective of this study was to provide a fish diet supplemented with Se-ENNs as nutrients and evaluate the stability of the particles in the feed matrix against dissolution, aggregation and release over time. The toxicity of formulated diets containing Se-ENNs was compared with diets containing ionic Se in zebrafish (Danio rerio). We expect that the adsorption capacity of zebrafish is higher for smaller particles,25 thus we applied two sizes of Se-ENNs, 60 and 120 nm. To determine the adverse effects of Se-ENNs we evaluated the swimming behavior and the acetylcholinesterase (AChE) activity. Acetylcholine functions as an activator in the peripheral nervous system and has a major role in the enhancement of sensory perceptions in the central nervous system. This is thus a sensitive biomarker to determine safe levels versus excessive Se- and Se-ENNs levels in the diet. Fish behavior is an ecologically relevant indicator, widely studied and often becoming evident at concentrations at which other sub-lethal effects are not visible yet.26 Impaired swimming of fish can alter a number of other essential ecological functions such as food acquisition, grouping, predator avoidance and reproduction.27 In the current study two synthesized Se-ENNs with different size were imbedded in fish feed. The stability of the Se-ENNs against dissolution, particle release and aggregation in the diets was monitored. Thereupon, the homogeneity of the Se-ENNs distribution in the diet was confirmed and the formulated feed was used to feed zebrafish for 21 days. The behavior of the treated fish was monitored using an in-house developed 3-dimensional monitoring system. The particle number and size distribution of the particles in the brain tissue28 were measured because the brain is considered as a target location for some nanomaterials.29 Moreover, sub-lethal adverse effects in the brain can easily be reflected in swimming behavior.30
Materials and methods
Chemicals
All used chemicals were purchased from Sigma-Aldrich (St. Louis, MO, Germany) in ACS purity. Sodium selenite was used as a control to supplement the feed with Se. S-Acetylthiocholine-iodide (ASChI, 98%) and 5,5-dithio-bis-(2-nitrobenzoic acid) (DTNB) were applied to determine the AchE activities. Selenous acid (98%) was used for the synthesis of Se-ENNs. Deionized water was prepared using reverse osmosis (Aqual 25: Tisnov, Czech Republic). The MilliQ (MQ) water was provided using a MQ Direct QUV, equipped with a UV lamp (Aqua Osmotic, Tisnov, Czech Republic).Synthesis of Se-ENNs
The 60 nm Se-ENNs were synthesized using a chemical reduction process. Briefly, poly(vinylpyrrolidone) (denoted as PVP, Mw = 10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 Da, 10 wt%) was dissolved in MQ water (49.6 mL) followed by addition of selenous acid (0.4 mL, 1.0 M). The mixture was then vigorously stirred at room temperature (first solution). The second solution was prepared from MQ water (44.2 mL) and ascorbic acid (0.8 mL, 1.0 M), and used as a reducing agent. The second solution was added dropwise at a rate of 0.8 mL min−1 to the first solution. The synthesized Se-ENNs were collected by centrifugation at 10000 RCF (Eppendorf centrifuge 5417R, Hamburg, Germany) at 10 °C for 10 min and subsequently washed three times with MQ water.
000 Da, 10 wt%) was dissolved in MQ water (49.6 mL) followed by addition of selenous acid (0.4 mL, 1.0 M). The mixture was then vigorously stirred at room temperature (first solution). The second solution was prepared from MQ water (44.2 mL) and ascorbic acid (0.8 mL, 1.0 M), and used as a reducing agent. The second solution was added dropwise at a rate of 0.8 mL min−1 to the first solution. The synthesized Se-ENNs were collected by centrifugation at 10000 RCF (Eppendorf centrifuge 5417R, Hamburg, Germany) at 10 °C for 10 min and subsequently washed three times with MQ water.
To synthesize the 120 nm Se-ENNs, PVP (Mw = 10000 Da, 10 wt%) was dissolved in MQ water (49.2 mL) followed by the addition of ascorbic acid (0.8 mL, 1.0 M) as reducing agent. The mixture was then stirred vigorously at room temperature. A second solution was prepared from MQ water (44.6 mL) and selenous acid (0.4 mL, 1.0 M). The second solution was added dropwise at a rate of 0.8 mL min−1 to the first solution. The synthesized ENNs were collected by centrifugation at 10000 RCF for 10 min and washed three times with MQ water.
Characterization of the Se-ENNs dispersions
Dispersions of the Se-ENNs in MQ water were tip sonicated for 5 min using a SONOPULS ultrasonicator (Bandelin Electronic, Berlin, Germany). After sonication, the hydrodynamic size and ζ potential of the ENNs in the dispersion were immediately measured using a Zetasizer Nanodevice (Malvern Panalytical, Netherlands). Scanning electron microscope (SEM) images of the Se-ENNs dispersions in MQ water were obtained using MAIA 3 SEM (TESCAN Company, Brno, Czech Republic) operated at an accelerating voltage 5 kV to determine the size and shape of the particles. The number concentration and the size distribution of the Se-ENNs were measured using single-particle inductively coupled plasma mass spectrometry (spICP-MS). All spICP-MS measurements were performed on a PerkinElmer NexION 300D ICP-MS operating in a single-particle mode, using the conditions in Table S1 (ESI†). Both samples of Se-ENNs in MQ water and internalized in tissues were pictured at room temperature using a SEM Magellan 400L equipped with an energy dispersive spectroscopy (EDS) detector Octane Elect Super (EDAX)31 (S2, ESI†).Diet formulation
Commercial zebrafish feed (GEMMA MICRO zebrafish diet, Nederweert, the Netherlands) was used in the current study. The percentage of the different components in the diets based on information provided by the manufacturer are reported in Table S2 (ESI†). To prepare the experimental diets the feed was oven-dried (30 °C) for 6 h to reach a constant weight. Subsequently, the feed was powdered and oven-dried again under the same conditions. A stock solution (100 mg L−1) of the Se-ENNs was prepared in MQ water followed by sonication in a bath sonicator (35 kHz frequency, DT 255, Bandelin electronic, Sonorex digital, Berlin, Germany) for 10 min. An aliquot of the Se-ENNs dispersion was sprayed on commercial feed pellets to reach final concentrations of 2.4 and 240 mg Se-ENNs per kg feed (Table 1). The first concentration (2.4 mg kg−1) was selected to represent the fish required dietary concentration of Se (optimal level). This concentration is the highest recommended Se concertation in fish diet, although it is different from fish to fish.32,33 The 240 mg kg−1 was chosen because it is a lethal concentration of Se to fish.34 We selected these two concentrations because they represent exposure concentrations at which dietary uptake of Se may have effects on fish. The Se-ENNs supplemented feed was slowly mixed for 1 h and oven-dried for 30 min. The supplemented diets were covered by a 3.3% gelatine solution and sieved through a mesh to obtain <1 mm granules. The granules covered with a thin gelatin layer were oven-dried at 40 °C for 2 h and then kept in airtight containers at −20 °C. The negative and positive control diets were prepared using the same procedure except that the spraying with Se-ENNs was replaced by spraying with MQ water or with a sodium selenite solution as a Se stock. For the positive controls, the final concentration of Se in the feed was 2.4 and 240 mg per kg feed (Table 1).| Treatment | Containing | Used as |
|---|---|---|
| Control | No Se-ENNs and no Se | Control |
| 2.4-Se | 2.4 mg selenite per kg feed | Positive control 1 where we do expect optimal Se levels |
| 240-Se | 240 mg selenite per kg feed | Positive control 2 where we expect toxicity |
| 2.4-Se-ENN-60 | 2.4 mg of 60 nm Se-ENNs per kg feed | Feed-1 60 nm |
| 240-Se-ENN-60 | 240 mg of 60 nm Se-ENNs per kg feed | Feed-2 60 nm |
| 2.4-Se-ENN-120 | 2.4 mg of 120 nm Se-ENNs per kg feed | Feed-3 120 nm |
| 240-Se-ENN-120 | 240 mg of 120 nm Se-ENNs per kg feed | Feed-4 120 nm |
Release experiments
To measure the amount of Se released from the formulated diets, release experiments were performed. Briefly, 1 g of each diet was dispersed in 100 ml of MQ water and stirred for 72 min using a magnet stirrer. Aliquots of the suspension were taken after 6, 12, 24, 36, 48, 60 and 72 min and centrifuged according to the method reported previously35 to separate the dissolved Se from particulate Se. The time of sampling was arbitrarily selected to cover the duration of the feeding process based on personal observations. The total concentration of the Se in the supernatant was measured using ICP-MS (PerkinElmer NexION 300D ICP-MS, Birmingham, UK).Exposure experiments
All zebrafish experiments were performed with the approval of the ethics committee of the Research Center of Aquaculture and Biodiversity of Hydrocenoses, the University of South Bohemia in Ceske Budejovice, Czech Republic (MSMT-6744/2018-4). Zebrafish were obtained from the Research Facility at the Faculty of Fisheries and Protection of Waters, the University of South Bohemia, Vodnany, Czech Republic. Adult zebrafish were acclimated to experimental conditions for 14 days before the start of the experiment. The experiment was performed in 40 L glass aquaria and the fish were fed with a diet of commercial fish feed (the same feed was used to prepare the formulated feed) twice a day (1.5% body weight). Nine fishes were kept in each aquarium, with three replicates per treatment. The aquaria were filled with aged tap water and aerated using air pumps. The physicochemical conditions of the exposure media are reported in ESI† (S4). During the exposure, fish were fed their respective treatment feed and the remaining feed was removed from the aquaria after 30 min. The aquaria were cleaned and the water was changed three times during the exposure period.Characterization and quantification of Se and Se-ENNs in the samples
To measure the total mass of the Se in the samples (feed, feces, exposure media and tissues), the samples were digested for 60 min with HNO3 (65%) at 130 °C followed by 2 h of additional digestion with HClO4 at 170 °C on a heating block. The total mass concentrations of Se in the solutions were measured using ICP-MS (PerkinElmer NexION 300D ICP-MS, Birmingham, UK). The isotope 80Se was measured with a plasma flow of 15 L min−1 and a nebulizer gas flow of 1 L min−1.The extraction of Se-ENNs from tissues was carried out following the method reported in our previous study.25 This allowed us to measure the size and number concentration of the particles using spICP-MS. The operational parameters of the spICP-MS are summarized in Table S1 (ESI†). We used gold nanoparticles (RM-8013, Boulder, CO, USA) consisting of spherical 60 nm particles with a mass concentration of 50 mg L−1 as reference materials to operate the spICP-MS. A dispersion of Se-ENNs in MQ water was used as a control to evaluate whether the particle extraction approached altered the Se-ENNs, according to the method reported in our previous study.25
Analysis of the swimming activity
The exposure experiments were arranged independently in triplicates for each treatment group (Se and Se-ENNs). Swimming behavioral analysis of fish was performed 24 h before the start of the exposure, and at days 10 and 20 of the exposure by measuring the swimming speed of the fish (see S5, ESI†). A non-invasive system for small tank monitoring was applied to measure the swimming distance of fish using a 3D/near infrared camera linked to a computer for real time data recording (see S5, ESI†).Determining the acetylcholinesterase (AChE) activity
The AChE activity was measured using a spectrophotometrical assay and the Ellman's reaction.36 The gills and brain tissue were homogenized in 0.05 M Tris-HCl buffer (pH 8). The homogenate was centrifuged at 10000 RCF for 20 min at 4°C and the supernatant was removed to determine the AChE activity as described by Roex et al.,37 Briefly, the activity of AChE was determined in a suspension containing ASChI and DTNB at a final concentration of 0.3 mM and 1 mM, respectively. The absorbance was determined at 402nm during 4 min using UV-vis spectroscopy (Agilent 8453 UV/vis, Markham, Canada). The AChE activity is presented as the percentage of the average activity as compared to the negative controls.
Data analysis
The statistical analysis was performed using Sigmaplot version 14 and SPSS v 19. For the behavioral data a repeated measures ANOVA was conducted. The Shapiro–Wilk's test was used to test for normality, and Cochran's and Hartley's were used to test the assumption of normality and homoscedasticity, respectively. To compare the differences between adsorption of Se by zebrafish and inhibition of the AChE activity between tissues we applied t-test. To determine the signification differences between treatments and swimming behavior we used ANOVA followed Duncan's multiple range tests. The Kruskal–Wallis test and the Friedman ANOVA were used to analyzing the swimming speed. All data is presented as mean ± standard error (SE).Results and discussion
Characterization of the synthesized Se-ENNs
The Se-ENNs were characterized with regard to their properties and the stability of the particles was monitored over time. The SEM pictures shows that the particles are spherical and no aggregates of particle in MQ water were observed (Fig. 1a). The hydrodynamic size (nm) and the SEM measured size are reported in Table S3 (ESI†). Analysis of the homoaggregation over time showed that the particles are stable over the 24 h of measurement (Fig. 1b). The EDX map and the results of the analysis (spectrum) of the particles in MQ water are reported in Fig. S4 and S5 (ESI†). The dissolution profile of the particles in MQ water is shown in Fig. 1c. The percentage of dissolved Se was less than 1% of the total Se-ENNs for both particles in MQ water over 24 h of incubation time, showing that the particles were stable against dissolution in MQ water. The spICP-MS data for the size distribution of the particles in MQ water showed that the sizes of the particles are normally distributed. This confirmed that very low particle aggregation in MQ water occurred (Fig. 1d). | ||
| Fig. 1 a) Scanning electron microscope images of the Se-ENNs. b) The aggregation profile of the Se-ENNs over 24 h time in MQ water measured using dynamic light scattering. c) The dissolution profile of the particles in MQ water over 24 h measured using spICP-MS. d) Number size distribution of the particles in MQ water measured using spICP-MS. | ||
Characterization of the Se-ENNs in the fish diet
We synthesized 60 nm and 120 Se-ENNs to determine the influence of particle size on their absorption to the gut epithelium in the fish body. We assumed that, due to its smaller size, the 60 nm particles have a higher absorption rate compared to the 120 nm particles.38 To assure a homogenous distribution of the Se-ENNs and Se in the prepared diets, 6 random samples from each diet were analyzed to measure the total mass concentration of the Se (Table 2). The low variance within and across treatments indicates that the Se-ENNs and Se ions were indeed homogeneously distributed in the feed.| Treatment | Nominal mass (mg kg−1 feed) | Measured total mass ± SD (mg kg−1 feed) |
|---|---|---|
| 2.4-Se: 2.4 mg selenite per kg feed, 240-Se: 240 mg selenite per kg feed, 2.4-Se-ENN-60: 2.4 mg of 60 nm Se-ENNs per kg feed, 240-Se-ENN-60: 240 mg of 60 nm Se-ENNs per kg feed, 2.4-Se-ENN-120: 2.4 mg of 120 nm Se-ENNs per kg feed, 240-Se-ENN-120: 240 mg of 120 nm Se-ENNs per kg feed. | ||
| 2.4-Se | 2.4 | 2.7 ± 0.5 |
| 240-Se | 240 | 221 ± 17 |
| 2.4-Se-ENN-60 | 2.4 | 1.6 ± 0.4 |
| 240-Se-ENN-60 | 240 | 197 ± 16 |
| 2.4-Se-ENN-120 | 2.4 | 1.8 ± 0.6 |
| 240-Se-ENN-120 | 240 | 213 ± 19 |
The number size distribution of the particles in the feed was measured using spICP-MS (Fig. 2a). The number concentration of Se particles in the 2.4-Se and 240-Se samples was lower than the detection limit of the instrument (0.01 μg L−1), indicating that no Se particles were formed in these feeds. The number size distribution of Se-ENNs in 2.4-Se-ENN-60 and 240-Se-ENN-60 was similar to the number size distribution of Se-ENNs in MQ water. The size distribution of the Se-ENNs in 2.4-Se-ENN-120 and 240-Se-ENN-120 was wider than the size distribution measured for 120 nm Se-ENNs in MQ water, implying that the particles in these formulations may have slightly aggregated due to the influence of the feed matrices as compared to the same particles in MQ water. The influence of food matrices on the size distribution of ENNs has been previously reported for silica nanomaterials,39,40 which highlights the importance of considering feed matrices on embedded ENNs.
 | ||
| Fig. 2 a) The size distribution of the Se-ENNs in the fish feed measured by spICP-MS. b) Se released from the fish feed in the exposure media over 72 h duration. c) Percentage of Se recovery, showing the adsorption of Se by zebrafish (Danio rerio). 2.4-Se: 2.4 mg selenite per kg feed, 240-Se: 240 mg selenite per kg feed, 2.4-Se-ENN-60: 2.4 mg of 60 nm Se-ENNs per kg feed, 240-Se-ENN-60: 240 mg of 60 nm Se-ENNs per kg feed, 2.4-Se-ENN-120: 2.4 mg of 120 nm Se-ENNs per kg feed, 240-Se-ENN-120: 240 mg of 120 nm Se-ENNs per kg feed. | ||
The release of the Se-ENNs and Se ions from the feed in the exposure media was investigated over 72 h. No particles could be measured in the exposure media in all treatments. The concentration of the dissolved Se in the exposure media over 72 h is illustrated in Fig. 2b. The highest Se release was observed for 2.4-Se and 240-Se. The Se releases from Se-ENNs supplemented diets were low (Fig. 2b) regardless of the size and quantity of the particles in the diet. A previous study using zinc oxide ENNs used the same procedure as we applied in the current study, and they also reported a low zinc release from the feed.41
Fish survival
The survival of the zebrafish in the control group (fish fed with no Se and Se-ENNs) and fish fed with any of the feeds containing ENNs was 100% during the exposure period. As previously reported, PVP is not toxic to zebrafish,42 thus we did not perform a PVP exposure test in this study. In addition, no mortality was observed in fish fed with diet containing 2.4 mg Se. However, there was significantly higher mortality rate (5 fish after 10 days treatment) in fish fed with diet contain 240 Se compared to the control group. Our results are comparable to some previous studies. For example, a concentration of 9.60 mg Se per kg feed (wet weight) modified the behavior of striped bass and caused death to all fish within 78 d.43 Mortality due to the exposure to 26 mg of Se per kg feed (dw) was also observed in Sacramento splittail.44 Our data showed that the application of a high concentration of Se in its nano-form does not cause mortality in fish as it is reported for Se ions.Absorption of Se and Se-ENNs in fish
To determine the amount of Se and Se-ENNs adsorbed by fish, we measured the total mass concentration of the Se (μg kg−1) in the diets and in the feces of the fish. The concentrations of the measured Se in feces compared to the Se concentrations in the diet were calculated and reported as the percentage Se recovery in Fig. 2c. The data showed that no Se was recovered in the feces of fish fed with diets containing 2.4-Se, 2.4-Se-ENN-60, 240-Se-ENN-60 and 2.4-Se-ENN-120. This indicates that the Se-ENNs in the diets might be absorbed by fish because the spICP-MS measurement showed that there are no detectable Se-ENNs in the exposure media 30 min after feeding. The recovery of the Se for 240-Se was 34% and 25% for 240-Se-ENN-120. This indicates that not all of the Se and Se-ENNs in these diets were absorbed by the fish. This also shows that the absorption of the SE-ENNs is significantly (ANOVA, p < 0.05) higher than the absorption of Se. The absorption of the Se-ENNs with the smaller particle size (60 nm) was higher than the absorption of the Se-ENNs with the bigger particle size (120 nm). This finding supports our hypothesis that the size of ENNs influences the absorption of these materials, which is in agreement with previous studies showing that the adsorption capacity of zebrafish is higher for smaller particles.25Determining Se-ENNs in the brain of treated fish
Previous studies reported that some nanomaterials might end up in the brain of fish.25,29,30 Internalization of particles in the brain may cause some long-term behavioral adverse effects, which could appear in time points longer than the duration of the experiment in this study. Using spICP-MS we could only detect and quantify the number of 120 nm Se-ENNs in the brain of fish fed with 240-Se-ENN-120, indicating the penetration of these particles in the brain, while the smaller sized Se-ENNs (60 nm) were not detectable in the brain. The number size distribution of the 120 nm particles in the brain (Fig. 3) shifted toward a smaller particle size in comparison to the number size distribution of the particles in MQ water, with a second peak at around 105 nm. This could be associated with selective uptake of Se-ENNs in the brain and/or dissolution of the particles whether in the brain or in other tissues before reaching the brain. The Se-ENNs are stable in the exposure media for the duration of the treatment. However, they might undergo transformations such as dissolution and interaction with biomolecules after entering the fish body,45 which may lead to their assimilation as reported for other ENNs (the released Se from the particles can be incorporated into existing biological pathways).46,47 Previous studies reported that metallic nanomaterials dissolve to a certain extent in organisms' tissues as a function of particle size.45,48–52 Thus, two explanations can be put forward for our findings. First, the transformation (e.g. dissolution) of the 60 nm Se-ENNs is different compared to the transformation of the 120 nm Se-ENNs in the fish body. As a result, this variation led to the observed difference between the biodistribution of the ENNs in the fish body. Second, the 60 nm particles underwent dissolution and the resulting smaller sized particles entered the brain. However, their size might be lower than the detection limit of the spICP-MS for Se-ENNs (∼30 nm).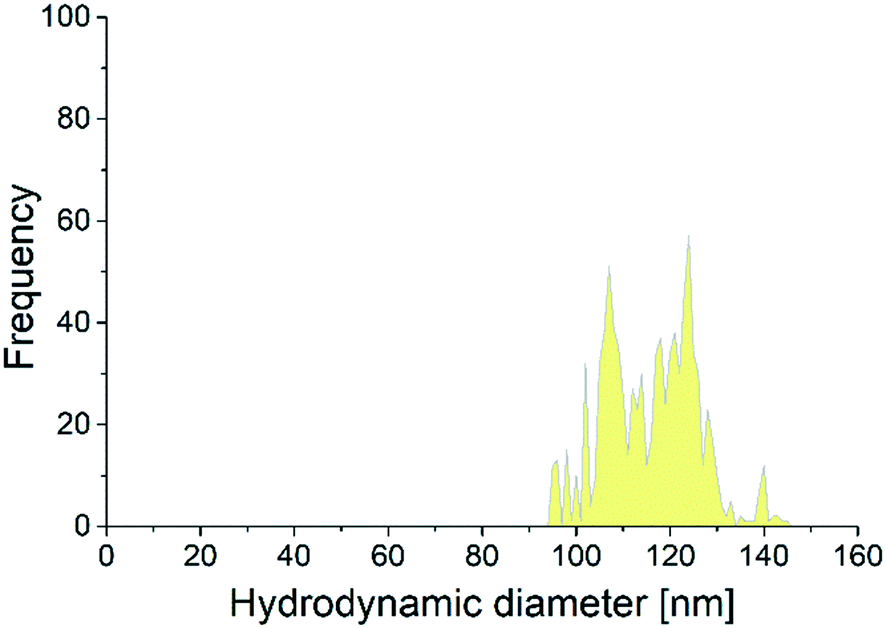 | ||
| Fig. 3 The number size distribution of the Se-ENNs in the brain of zebrafish (Danio rerio) fed with a 120 nm Se-ENNs containing diet. | ||
Behavioral changes of the treated fish
Changes in swimming behavior was used as an endpoint to investigate adverse effects of Se-ENNs. We excluded the fish fed with 240-Se (the diet containing 240 mg kg−1 Se) because of the observed high mortality rate in this treatment. The results are based on the average swimming distance of all replicates per second before the exposure and at days 10 and 20 of the exposure. Zebrafish, which were fed with Se-ENNs supplemented diets or the 2.4-Se diet showed a normal swimming behavior compared to the control (Fig. 4). Chronic dietary Se exposure has been reported to alter the behavioral and physiological responses in adult zebrafish.10 Abnormal swimming behavior was reported in juvenile bluegill after exposure to waterborne Se (0.64 mg Se per L)53 and in white sturgeon54 after exposure to dietary Se (41 mg Se per kg diet) for 8 weeks. Our findings showed that the swimming performance is normal in fish fed with diets containing nano-form of Se (Se-ENNs) at a level which was comparable to the level at which Se was lethal when not provided in a nano-form. This suggests that dietary uptake of Se-ENNs causes no behavioral alteration in fish compared to the dietary uptake of their traditional counterparts. | ||
| Fig. 4 Swimming behavior of zebrafish (Danio rerio) exposed to Se and Se-ENNs within different diets. No significant differences were observed between treatments (P > 0.05). 2.4-Se: 2.4 mg selenite per kg feed, 2.4-Se-ENN-60: 2.4 mg of 60 nm Se-ENNs per kg feed, 240-Se-ENN-60: 240 mg of 60 nm Se-ENNs per kg feed, 2.4-Se-ENN-120: 2.4 mg of 120 nm Se-ENNs per kg feed, 240-Se-ENN-120: 240 mg of 120 nm Se-ENNs per kg feed. | ||
Determining AChE activities in treated fish
In neurotoxicity, AChE activity is used as an indicator of exposure to Se.55 Here, we determined the activity of AChE in the brain and gills, as target organs, to determine the possible oxidative stress induced in fish fed with the formulated diets. The AChE activity in the fish fed with 240-Se was not determined due to the lethal effect observed in this treatment. There was no significant alteration in the AChE activity of the fish (brain and gills) fed with the Se-ENNs supplemented diet. AChE activity in the gills of fish fed with 2.4-Se supplemented diet decreased significantly (t-test, p < 0.05) in comparison to the control (Fig. 5b). A previous study, which investigated the oxidative stress in fish after exposure to waterborne Se for 4 weeks,55 reported that AChE activity was inhibited in the brain and muscle tissues of the exposed fish. The authors of this study suggested that waterborne Se exposure can significantly inhibit AChE activity. | ||
| Fig. 5 a) AChE activity determined in the brain and gills of zebrafish (Danio rerio) in each treatment after 21 days of exposure to Se and Se-ENNs. The letters (a and b) indicate the significant differences (p < 0.05) between the tissue and control (n = 9). | ||
The inhibition of the AChE activity in gills could be due to the release of the Se from the feed followed by the uptake of the waterborne Se through the gills. Fish gills have tight junctions, which make the uptake of particles through gills unlikely to happen even if there were any undetectable freely available Se-ENNs in the exposure media.25,29,30 Our study for the first time indicated that Se in nano-form does not inhibit AChE activity, as observed for the non-particulate form in the gills.
Conclusion
In this study, we provided fish diets containing a nano-form of Se (Se-ENNs) with two different particle sizes (60 and 120 nm) and investigated whether the modified diets cause mortality and adverse effects on swimming behavior and AChE activity to zebrafish after long term feeding. The Se-ENNs were stable against aggregation and dissolution in the exposure media. The release of the Se-ENNs from the formulated diets was low, indicating the stability of the diets in the exposure media. Both the Se-ENNs, 60 and 120 nm were absorbed in the fish body, but the smaller sized particles were found to have a higher extent of absorption than the larger ones. The spICP-MS data showed that the 120 nm Se-ENNs could penetrate the brain barrier and accumulate in the brain. Nevertheless, zebrafish fed with Se-ENNs containing diet, regardless of the particle size of the SE-ENNs, showed no abnormal swimming behavior over 21 days of exposure. On the contrary, fish fed with a diet containing 240 mg Se per kg feed showed mortality only after roughly one week of treatment. Moreover, the inhibition of AChE activity in the brain and gills of fish fed with diets containing 2.4 mg Se compared to a diet containing Se-ENNs indicated no impacts of Se-ENNs on AChE activity, and potentially limited impacts on the nervous system in general. This study presents important findings for designing safer nanomaterials, not only for use within aquaculture, but potentially also for other application in other foods.Author contributions statements
F. A. M. supervised, conceptualized, wrote and reviewed the paper. F. A. M., L. C., R. F. and E. Z. maintained the organisms, designed the experiments and performed the exposure test. F. A. M., K. S. and L. R. synthesized and characterized the nanoparticles. F. A. M. and P. C. performed the behavioral observations using a 3D/near infrared camera. V. K. and R. S. performed the microscope observations. F. A. M. and T. B. performed statistical analysis of the data and contributed in editing the paper. F. A. M., L. C., G. K. D., performed the toxicity and provided the nano-formulation diets. F. A. M., and E. V. J., performed the single particle and total mass analysis of the particles in the samples. M. V., and W. P., contributed in writing and editing the paper as well as conceptualization.Conflicts of interest
There are no conflicts to declare.Acknowledgements
The Nanofeed project has received funding from the European Union's Horizon 2020 research and innovation programme under grant agreement No. 652831 (AQUAEXCEL2020). This output reflects only the author's views and the European Union cannot be held responsible for any use that may be made of the information contained therein. The study was also partially supported by ERDF “Multidisciplinary research to increase application potential of nanomaterials in agricultural practice” (No. CZ.02.1.01/0.0/0.0/16_025/0007314). The electron microscopy investigations were supported by the Czech Science Foundation grant (17-15451S).References
- C. M. Moffitt and L. Cajas-Cano, Blue Growth: The 2014 FAO State of World Fisheries and Aquaculture, Fisheries, 2014, 39, 552–553 CrossRef.
- E. Roth and H. Rosenthal, Fisheries and aquaculture industries involvement to control product health and quality safety to satisfy consumer-driven objectives on retail markets in Europe, Mar. Pollut. Bull., 2006, 53, 599–605 CrossRef CAS PubMed.
- L. Ababouch, D. Alderman, K. Ashida, C. Boyd, S. Chimatiro, E. R. Cruz-Lacierda, A. Dalsgaard, P. Edwards, E. S. Garrett, K. Gopakumar, T. Hajime, L. T. Luu, E. G. Merino, H. J. Rim, T. Q. Son, S. Suwanrangsi, K. Tonguthai, M. Yndestad, J. Yulin, B. Aalvik, M. Boonyaratpalin, S. Suwannatos, N. Wongchinda, P. McClure, S. Adams, R. Khamphet, J. O. Naewbanij, G. Nash, U. Barg, P. K. Benembarek, M. Beveridge, C. L. Dos Santos, R. Fuchs, P. Howgate, F. Kaferstein, H. Kongkeo, M. Phillips, A. Reilly, R. Subasinghe and A. G. J. Tacon, Food safety issues associated with products from aquaculture – Report of a joint FAO/NACA/WHO Study Group, World Health Organ. Tech. Rep. Ser., 1999, 883, 1–56 Search PubMed.
- S. M. Garcia and C. H. Newton, Responsible Fisheries – an Overview of Fao Policy Developments (1945-1994), Mar. Pollut. Bull., 1994, 29, 528–536 CrossRef CAS.
- A. I. S. Luis, E. V. R. Campos, J. L. de Oliveira and L. F. Fraceto, Trends in aquaculture sciences: from now to use of nanotechnology for disease control, Rev. Aquacult., 2019, 11, 119–132 CrossRef.
- T. Behera, P. Swain, P. V. Rangacharulu and M. Samanta, Nano-Fe as feed additive improves the hematological and immunological parameters of fish, Labeo rohita H, Appl. Nanosci., 2014, 4, 687–694 CrossRef CAS.
- M. Gopi, B. Pearlin, R. D. Kumar, M. Shanmathy and G. Prabakar, Role of Nanoparticles in Animal and Poultry Nutrition: Modes of Action and Applications in Formulating Feed Additives and Food Processing, Int. J. Pharmacol., 2017, 13, 724–731 CAS.
- S. Gangadoo, D. Stanley, R. J. Hughes, R. J. Moore and J. Chapman, Nanoparticles in feed: Progress and prospects in poultry research, Trends Food Sci. Technol., 2016, 58, 115–126 CrossRef CAS.
- S. M. Rodrigues, P. Demokritou, N. Dokoozlian, C. O. Hendren, B. Karn, M. S. Mauter, O. A. Sadik, M. Safarpour, J. M. Unrine, J. Viers, P. Welle, J. C. White, M. R. Wiesner and G. V. Lowry, Nanotechnology for sustainable food production: promising opportunities and scientific challenges, Environ. Sci.: Nano, 2017, 4, 767–781 RSC.
- J. K. Thomas and D. M. Janz, Dietary selenomethionine exposure in adult zebrafish alters swimming performance, energetics and the physiological stress response, Aquat. Toxicol., 2011, 102, 79–86 CrossRef CAS.
- J. W. Hilton, P. V. Hodson and S. J. Slinger, The Requirement and Toxicity of Selenium in Rainbow Trout (Salmo Gairdneri), J. Nutr., 1980, 110, 2527–2535 CrossRef CAS PubMed.
- J. R. Arthur, R. C. McKenzie and G. J. Beckett, Selenium in the Immune System, J. Nutr., 2003, 133, 1457S–1459S CrossRef CAS.
- R. L. Davis and J. E. Spallholz, Inhibition of selenite-catalyzed superoxide generation and formation of elemental selenium (Se°) by copper, zinc, and aurintricarboxylic acid (ATA), Biochem. Pharmacol., 1996, 51, 1015–1020 CrossRef CAS.
- J. E. Spallholz, On the nature of selenium toxicity and carcinostatic activity, Free Radical Biol. Med., 1994, 17, 45–64 CrossRef CAS PubMed.
- D. Han, S. Xie, M. Liu, X. Xiao, H. Liu, X. Zhu and Y. Yang, The effects of dietary selenium on growth performances, oxidative stress and tissue selenium concentration of gibel carp (Carassius auratus gibelio), Aquacult. Nutr., 2011, 17, E741–E749 CrossRef.
- S. Skalickova, V. Milosavljevic, K. Cihalova, P. Horky, L. Richtera and V. Adam, Selenium nanoparticles as a nutritional supplement, Nutrition, 2017, 33, 83–90 CrossRef CAS PubMed.
- S. Ashouri, S. Keyvanshokooh, A. P. Salati, S. A. Johari and H. Pasha-Zanoosi, Effects of different levels of dietary selenium nanoparticles on growth performance, muscle composition, blood biochemical profiles and antioxidant status of common carp (Cyprinus carpio), Aquaculture, 2015, 446, 25–29 CrossRef CAS.
- M. A. O. Dawood, S. Koshio, A. I. Zaineldin, H. Van Doan, E. M. Moustafa, M. M. Abdel-Daim, M. Angeles Esteban and M. S. Hassaan, Dietary supplementation of selenium nanoparticles modulated systemic and mucosal immune status and stress resistance of red sea bream (Pagrus major), Fish Physiol. Biochem., 2019, 45, 219–230 CrossRef CAS PubMed.
- M. A. O. Dawood, M. Zommara, N. M. Eweedah, A. I. Helal and M. A. Aboel-Darag, The potential role of nano-selenium and vitamin C on the performances of Nile tilapia (Oreochromis niloticus), Environ. Sci. Pollut. Res., 2020, 27, 9843–9852 CrossRef CAS PubMed.
- M. S. Izquierdo, W. Ghrab, J. Roo, K. Hamre, C. M. Hernández-Cruz, G. Bernardini, G. Terova and R. Saleh, Organic, inorganic and nanoparticles of Se, Zn and Mn in early weaning diets for gilthead seabream (Sparus aurata; Linnaeus, 1758), Aquacult. Res., 2017, 48, 2852–2867 CrossRef CAS.
- P. Swain, R. Das, A. Das, S. K. Padhi, K. C. Das and S. S. Mishra, Effects of dietary zinc oxide and selenium nanoparticles on growth performance, immune responses and enzyme activity in rohu, Labeo rohita (Hamilton), Aquacult. Nutr., 2019, 25, 486–494 CrossRef CAS.
- X. Zhou, Y. Wang, Q. Gu and W. Li, Effects of different dietary selenium sources (selenium nanoparticle and selenomethionine) on growth performance, muscle composition and glutathione peroxidase enzyme activity of crucian carp (Carassius auratus gibelio), Aquaculture, 2009, 291, 78–81 CrossRef CAS.
- T. V. Duncan, The communication challenges presented by nanofoods, Nat. Nanotechnol., 2011, 6, 683–688 CrossRef CAS PubMed.
- J. Mal, W. J. Veneman, Y. V. Nancharaiah, E. D. van Hullebusch, W. J. G. M. Peijnenburg, M. G. Vijver and P. N. L. Lens, A comparison of fate and toxicity of selenite, biogenically, and chemically synthesized selenium nanoparticles to zebrafish (Danio rerio) embryogenesis, Nanotoxicology, 2017, 11, 87–97 CrossRef CAS PubMed.
- F. Abdolahpur Monikh, L. Chupani, E. Zuskova, R. Peters, M. Vancova, M. G. Vijver, P. Porcal and W. J. G. M. Peijnenburg, Method for Extraction and Quantification of Metal-Based Nanoparticles in Biological Media: Number-Based Biodistribution and Bioconcentration, Environ. Sci. Technol., 2019, 53, 946–953 CrossRef CAS PubMed.
- J. Cachat, A. Stewart, L. Grossman, S. Gaikwad, F. Kadri, K. M. Chung, N. Wu, K. Wong, S. Roy, C. Suciu, J. Goodspeed, M. Elegante, B. Bartels, S. Elkhayat, D. Tien, J. Tan, A. Denmark, T. Gilder, E. Kyzar, J. DiLeo, K. Frank, K. Chang, E. Utterback, P. Hart and A. V. Kalueff, Measuring behavioral and endocrine responses to novelty stress in adult zebrafish, Nat. Protoc., 2010, 5, 1786–1799 CrossRef CAS PubMed.
- J. S. Weis and A. Candelmo, Pollutants and fish predator/prey behavior: A review of laboratory and field approaches, Curr. Zool., 2012, 58, 9–20 CrossRef.
- F. Abdolahpur Monikh, L. Chupani, E. Zusková, R. Peters, M. Vancová, M. G. Vijver, P. Porcal and W. J. G. M. Peijnenburg, Method for Extraction and Quantification of Metal-Based Nanoparticles in Biological Media: Number-Based Biodistribution and Bioconcentration, Environ. Sci. Technol., 2019, 53, 946–953 CrossRef CAS.
- R. D. Handy, R. Owen and E. Valsami-Jones, The ecotoxicology of nanoparticles and nanomaterials: current status, knowledge gaps, challenges, and future needs, Ecotoxicology, 2008, 17, 315–325 CrossRef CAS.
- K. Mattsson, E. V. Johnson, A. Malmendal, S. Linse, L.-A. Hansson and T. Cedervall, Brain damage and behavioural disorders in fish induced by plastic nanoparticles delivered through the food chain, Sci. Rep., 2017, 7, 11452 CrossRef PubMed.
- S. Marvanová, P. Kulich, R. Skoupý, F. Hubatka, M. Ciganek, J. Bendl, J. Hovorka and M. Machala, Size-segregated urban aerosol characterization by electron microscopy and dynamic light scattering and influence of sample preparation, Atmos. Environ., 2018, 178, 181–190 CrossRef.
- M. Lorentzen, A. Maage and K. Julshamn, Effects of dietary selenite or selenomethionine on tissue selenium levels of Atlantic salmon (Salmo salar), Aquaculture, 1994, 121, 359–367 CrossRef CAS.
- D. J. Wise, J. R. Tomasso, D. M. Gatlin, S. C. Bai and V. S. Blazer, Effects of Dietary Selenium and Vitamin E on Red Blood Cell Peroxidation, Glutathione Peroxidase Activity, and Macrophage Superoxide Anion Production in Channel Catfish, J. Aquat. Anim. Health, 1993, 5, 177–182 CrossRef.
- P. V. Hodson and J. W. Hilton, The Nutritional-Requirements and Toxicity to Fish of Dietary and Waterborne Selenium, Ecol. Bull., 1983, 335–340 CAS.
- D. Arenas-Lago, F. Abdolahpur Monikh, M. G. Vijver and W. J. G. M. Peijnenburg, Dissolution and aggregation kinetics of zero valent copper nanoparticles in (simulated) natural surface waters: Simultaneous effects of pH, NOM and ionic strength, Chemosphere, 2019, 226, 841–850 CrossRef CAS PubMed.
- G. L. Ellman, K. D. Courtney, V. Andres and R. M. Featherstone, A new and rapid colorimetric determination of acetylcholinesterase activity, Biochem. Pharmacol., 1961, 7, 88–95 CrossRef CAS.
- E. W. M. Roex, R. Keijzers and C. A. M. van Gestel, Acetylcholinesterase inhibition and increased food consumption rate in the zebrafish, Danio rerio, after chronic exposure to parathion, Aquat. Toxicol., 2003, 64, 451–460 CrossRef CAS.
- R. D. Handy, T. B. Henry, T. M. Scown, B. D. Johnston and C. R. Tyler, Manufactured nanoparticles: their uptake and effects on fish—a mechanistic analysis, Ecotoxicology, 2008, 17, 396–409 CrossRef CAS PubMed.
- S. Wagner, S. Legros, K. Loeschner, J. Liu, J. Navratilova, R. Grombe, T. P. J. Linsinger, E. H. Larsen, F. von der Kammer and T. Hofmann, First steps towards a generic sample preparation scheme for inorganic engineered nanoparticles in a complex matrix for detection, characterization, and quantification by asymmetric flow-field flow fractionation coupled to multi-angle light scattering and ICP-MS, J. Anal. At. Spectrom., 2015, 30, 1286–1296 RSC.
- M.-R. Go, S.-H. Bae, H.-J. Kim, J. Yu and S.-J. Choi, Interactions between Food Additive Silica Nanoparticles and Food Matrices, Front. Microbiol., 2017, 8, 1013 CrossRef.
- L. Chupani, H. Niksirat, V. Lünsmann, S.-B. Haange, M. von Bergen, N. Jehmlich and E. Zuskova, Insight into the modulation of intestinal proteome of juvenile common carp (Cyprinus carpio L.) after dietary exposure to ZnO nanoparticles, Sci. Total Environ., 2018, 613–614, 62–71 CrossRef CAS.
- C. M. Powers, T. A. Slotkin, F. J. Seidler, A. R. Badireddy and S. Padilla, Silver nanoparticles alter zebrafish development and larval behavior: Distinct roles for particle size, coating and composition, Neurotoxicol. Teratol., 2011, 33, 708–714 CrossRef CAS.
- D. J. Coughlan and J. S. Velte, Dietary Toxicity of Selenium-Contaminated Red Shiners to Striped Bass, Trans. Am. Fish. Soc., 1989, 118, 400–408 CrossRef CAS.
- S. J. Teh, X. Deng, D.-F. Deng, F.-C. Teh, S. S. O. Hung, T. W. M. Fan, J. Liu and R. M. Higashi, Chronic Effects of Dietary Selenium on Juvenile Sacramento Splittail (Pogonichthys macrolepidotus), Environ. Sci. Technol., 2004, 38, 6085–6093 CrossRef CAS PubMed.
- M. Riediker, D. Zink, W. Kreyling, G. Oberdörster, A. Elder, U. Graham, I. Lynch, A. Duschl, G. Ichihara, S. Ichihara, T. Kobayashi, N. Hisanaga, M. Umezawa, T.-J. Cheng, R. Handy, M. Gulumian, S. Tinkle and F. Cassee, Particle toxicology and health – where are we?, Part. Fibre Toxicol., 2019, 16, 19 CrossRef PubMed.
- F. M. Hilty, M. Arnold, M. Hilbe, A. Teleki, J. T. N. Knijnenburg, F. Ehrensperger, R. F. Hurrell, S. E. Pratsinis, W. Langhans and M. B. Zimmermann, Iron from nanocompounds containing iron and zinc is highly bioavailable in rats without tissue accumulation, Nat. Nanotechnol., 2010, 5, 374–380 CrossRef CAS PubMed.
- F. Rohner, F. O. Ernst, M. Arnold, M. Hilbe, R. Biebinger, F. Ehrensperger, S. E. Pratsinis, W. Langhans, R. F. Hurrell and M. B. Zimmermann, Synthesis, Characterization, and Bioavailability in Rats of Ferric Phosphate Nanoparticles, J. Nutr., 2007, 137, 614–619 CrossRef CAS PubMed.
- J. T. Buchman, N. V. Hudson-Smith, K. M. Landy and C. L. Haynes, Understanding Nanoparticle Toxicity Mechanisms To Inform Redesign Strategies To Reduce Environmental Impact, Acc. Chem. Res., 2019, 52, 1632–1642 CrossRef CAS PubMed.
- D. Drobne, S. Novak, I. Talaber, I. Lynch and A. J. Kokalj, The Biological Fate of Silver Nanoparticles from a Methodological Perspective, Materials, 2018, 11, 12 CrossRef PubMed.
- S. Manzo, A. Rocco, R. Carotenuto, F. De Luca Picione, M. L. Miglietta, G. Rametta and G. Di Francia, Investigation of ZnO nanoparticles' ecotoxicological effects towards different soil organisms, Environ. Sci. Pollut. Res., 2011, 18, 756–763 CrossRef CAS PubMed.
- L. V. Stebounova, E. Guio and V. H. Grassian, Silver nanoparticles in simulated biological media: a study of aggregation, sedimentation, and dissolution, J. Nanopart. Res., 2011, 13, 233–244 CrossRef CAS.
- Z. McKenzie, M. Kendall, R.-M. Mackay, H. Whitwell, C. Elgy, P. Ding, S. Mahajan, C. Morgan, M. Griffiths, H. Clark and J. Madsen, Surfactant protein A (SP-A) inhibits agglomeration and macrophage uptake of toxic amine modified nanoparticles, Nanotoxicology, 2015, 9, 952–962 CrossRef PubMed.
- L. Cleveland, E. E. Little, D. R. Buckler and R. H. Wiedmeyer, Toxicity and bioaccumulation of waterborne and dietary selenium in juvenile bluegill (Lepomis macrochirus), Aquat. Toxicol., 1993, 27, 265–279 CrossRef CAS.
- D. H. Tashjian, S. J. Teh, A. Sogomonyan and S. S. O. Hung, Bioaccumulation and chronic toxicity of dietary l-selenomethionine in juvenile white sturgeon (Acipenser transmontanus), Aquat. Toxicol., 2006, 79, 401–409 CrossRef CAS PubMed.
- J.-H. Kim and J.-C. Kang, Oxidative stress, neurotoxicity, and non-specific immune responses in juvenile red sea bream, Pagrus major, exposed to different waterborne selenium concentrations, Chemosphere, 2015, 135, 46–52 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d0en00240b |
| This journal is © The Royal Society of Chemistry 2020 |