DOI:
10.1039/C9RA05181C
(Paper)
RSC Adv., 2019,
9, 33863-33871
The rapid effects of eicosapentaenoic acid (EPA) enriched phospholipids on alleviating exercise fatigue in mice
Received
8th July 2019
, Accepted 5th October 2019
First published on 22nd October 2019
Abstract
It has been reported that docosahexaenoic acid/eicosapentaenoic acid (DHA/EPA) and phospholipids (PLs) play an important role in alleviating exercise fatigue. However, the difference of DHA and EPA in ameliorating exercise fatigue is still unclear. Furthermore, the comparative study about DHA/EPA-PLs and nonpolar DHA/EPA on exercise fatigue has not been reported. In the present study, the effects of DHA and EPA on exercise fatigue was firstly compared by conducting an exhaustion test, and the results showed that triglyceride (TG) with high ratio of EPA had a more significant effect on alleviating exercise fatigue than TG with a low ratio of EPA in mice. Therefore, eicosapentaenoic acid–ethyl ester (EPA–EE) and EPA–PL were then selected to compare the rapid effects of polar and nonpolar DHA/EPA on exercise fatigue in mice by a weight-loaded swimming exhaustion test. A single intake of EPA–PL but not EPA–EE significantly alleviated exercise fatigue in mice by increasing the lactic acid recycling rate as well as inhibiting glycogen consumption and muscle injury, suggesting that EPA–PL exhibited a rapid effect on alleviating exercise fatigue. The study might represent a potential novel candidate or targeted dietary pattern for alleviating exercise fatigue.
1. Introduction
Exercise fatigue is the transient inability to maintain optimal exercise performance caused by intense exercise loading and inadequate rest, and results in a negative impact on physiological function, such as immune, endocrine, antioxidant system, nervous system and so on.1–4 Epidemiological data suggested a high prevalence of fatigue induced by physical exertion in laborers that disturbs health and life quality.5 However, most people who suffer from fatigue symptoms induced by physical exertion would not take the initiative to lighten the burden of work. Therefore, it is an urgent mission to find a targeted dietary patterns or natural bio-activator for alleviating exercise fatigue.
Increasing evidence has indicated the anti-fatigue effect of ω-3 long chain polyunsaturated fatty acids (ω-3 LCPUFA), including docosahexaenoic acid (DHA; 22![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :6 ω-3) and eicosapentaenoic acid (EPA; 20:5 ω-3). Double-blind studies suggested that fish oil supplementation inhibited muscle damage and alleviated delayed onset muscle soreness (DOMS) induced by exercise overloading in untrained individuals.6,7 Animal studies reported that ω-3 LCPUFA could elicit muscle membrane DHA enrichment, alleviate DOMS, and improve regenerative capacity of skeletal muscle cells, thereby benefiting exercise performance8,9 Moreover, studies have shown that there was a highly significant relevance between dietary and/or muscle levels of ω-3 LCPUFA and exercise performance.10 Notably, several studies found that DHA and EPA exhibited different efficacy in alleviating cardiopathy, mental health disorders, etc.11,12 However, the difference of DHA and EPA in ameliorating exercise fatigue is still unclear. Furthermore, the rapid effect of DHA/EPA on exercise fatigue has not been reported yet.
:6 ω-3) and eicosapentaenoic acid (EPA; 20:5 ω-3). Double-blind studies suggested that fish oil supplementation inhibited muscle damage and alleviated delayed onset muscle soreness (DOMS) induced by exercise overloading in untrained individuals.6,7 Animal studies reported that ω-3 LCPUFA could elicit muscle membrane DHA enrichment, alleviate DOMS, and improve regenerative capacity of skeletal muscle cells, thereby benefiting exercise performance8,9 Moreover, studies have shown that there was a highly significant relevance between dietary and/or muscle levels of ω-3 LCPUFA and exercise performance.10 Notably, several studies found that DHA and EPA exhibited different efficacy in alleviating cardiopathy, mental health disorders, etc.11,12 However, the difference of DHA and EPA in ameliorating exercise fatigue is still unclear. Furthermore, the rapid effect of DHA/EPA on exercise fatigue has not been reported yet.
Additionally, emerging evidence suggested that phospholipids (PLs), such as phosphatidylcholine (PC) and phosphatidylserine (PS), played an important role in alleviating exercise fatigue.13 Double-blind studies showed that PLs could reduce the diastolic blood pressure and cardio-ankle vascular index, suppress the severity of stress responses to exercise, thereby enhancing the exercise capacity.14 Interestingly, marine-derived DHA/EPA esterified to phospholipids (DHA/EPA–PLs) exhibit the structural characteristics of DHA/EPA and PLs.15 It has been reported that DHA/EPA–PLs had better effects than terrestrial PLs or nonpolar DHA/EPA enriched fish oil, such as triacylglycerol (TG) and ethyl ester (EE) forms, on improving neurodegenerative diseases, metabolic syndrome and so on.16,17 Le et al. reported that a 9 week DHA–PL enriched supplementation improved endurance exercise capacity and skeletal muscle mitochondrial function in adult rats.18 However, the comparative study about DHA/EPA–PLs and nonpolar DHA/EPA on exercise fatigue has not been reported.
In the present study, running exhaustion test were firstly used to compare the effects of DHA and EPA on exercise fatigue and the results showed that TG with high ratio of EPA had better effects on alleviating exercise fatigue than TG with low ratio of EPA in mice. Therefore, EPA-EE and EPA-PL were selected to evaluate the rapid effects of polar and nonpolar DHA/EPA on exercise fatigue in mice by weight-loaded swimming exhaustion test.
2. Materials and methods
2.1 Preparation and determination of DHA/EPA
DHA enriched phospholipids and EPA enriched phospholipids were prepared from squid (Sthenoteuthis oualaniensis) roe and sea cucumber according to the previous method, respectively.16 In brief, total lipids were extracted by the modified method of Folch,19 and then phospholipids were separated by the previous method.16 The composition of DHA/EPA–PLs was confirmed by high-performance liquid chromatography-evaporative light scattering detector (HPLC-ELSD) analysis, and the purity of phospholipids was more than 90%. EPA–EE (purity > 90%) was purchased from Sinomega Biotechnology Co., Ltd. (Zhoushan, China), and fish oil (DHA/EPA–TG) and algae oil (DHA–TG) were purchased from Weihai Boow Foods Co., Ltd. (Weihai, China). Fatty acid composition of each sample was determined and the result was shown in Table 1.
Table 1 Main fatty acid compositions of different DHA/EPA supplementsa
| Fatty acids (%) |
DHA/EPA–TG |
DHA–TG |
DHA/EPA–PL |
EPA–EE |
EPA–PL |
| Note: “—”, none detected. |
| C16:0 |
3.14 |
25.4 |
23.5 |
— |
4.32 |
| C16:1 |
1.00 |
— |
— |
4.52 |
5.39 |
| C18:0 |
4.12 |
— |
3.42 |
4.32 |
15.8 |
| C18:1 |
12.9 |
6.22 |
11.5 |
1.78 |
7.95 |
| C18:2n − 6 |
1.03 |
10.2 |
0.68 |
— |
— |
| C18:3n − 3 |
0.643 |
4.52 |
2.21 |
— |
— |
| C20:0 |
1.13 |
— |
5.56 |
— |
— |
| C20:1 |
0.234 |
— |
— |
1.55 |
10.6 |
| C20:2 |
— |
— |
— |
— |
— |
| C20:3 |
— |
— |
— |
— |
— |
| C20:4n − 6 |
2.27 |
— |
0.95 |
3.56 |
5.33 |
| C20:5n − 3 |
34.5 |
— |
10.6 |
72.3 |
49.2 |
| C22:1 |
0.932 |
9.71 |
1.82 |
— |
— |
| C22:6n − 3 |
24.7 |
42.9 |
32.6 |
9.59 |
0.68 |
2.2 Animals and treatments
Mice were purchased from Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China), and provided with food and water ad libitum in a room with a 12 h/12 h light/dark cycle, 23 ± 2 °C constant temperature and 60 ± 10% relative humidity. All protocols and procedures were in accordance with the guidelines of the ethical committee of experimental animal care at College of Food Science and Engineering, Ocean University of China (Qingdao, China). After acclimatization, the experiment was performed as follows.
2.2.1 Effects of DHA and EPA on the time to exhaustion in mice by consecutive administration. Male mice (18–22 g) were randomly divided into four groups, including control, DHA/EPA–TG, DHA–TG and DHA–PL, and fed with modified diet by AIN-93G for 10 days. The ingredients of the experimental diets were summarized in Table 2. The running exhaustion test was performed on the tenth day.
Table 2 Ingredients of the experimental dietsa
| Ingredients (g kg−1) |
Control |
DHA/EPA–TG |
DHA–TG |
DHA–PL |
| Note: “—”, none added. |
| Casein |
200 |
200 |
200 |
200 |
| Sucrose |
100 |
100 |
100 |
100 |
| Cornstarch |
399 |
399 |
399 |
399 |
| Corn oil |
50 |
45.8 |
44.2 |
44.2 |
| Lard |
150 |
137.3 |
132.5 |
132.7 |
| Cellulose |
50 |
50 |
50 |
50 |
| Mineral-salt mix |
35 |
35 |
35 |
35 |
| Vitamin mix |
10 |
10 |
10 |
10 |
| L-methionine |
3 |
3 |
3 |
3 |
| Choline bitartrate |
3 |
3 |
3 |
3 |
| DHA/EPA–TG |
— |
16.9 |
— |
— |
| DHA–TG |
— |
— |
23.3 |
— |
| DHA–PL |
— |
— |
— |
23.1 |
2.2.2 Rapid effects of EPA–EE and EPA–PL on the time to exhaustion in mice by single administration. Male mice (18–22 g) were employed in the experiment. Swim training without load was performed during the acclimatization period. According to body weight and swimming ability, the mice were divided into three groups, including control, EPA–EE and EPA–PL. After fasting for 10 hours (denoted by 0 hour), the mice in EPA–EE and EPA–PL groups were gavaged with EPA–EE and EPA–PL emulsions (1000 mg kg−1 body weight, standardized by EPA) and the control group was given equal corn oil emulsion. Weight-loaded swimming exhaustion test was performed at 2nd, 4th and 8th hour, respectively. After the last test, the mice were sacrificed rapidly or 5 minutes after rest, and the muscles and livers were collected and frozen with liquid nitrogen.
2.3 Running exhaustion test
A motor-driven wheel-track treadmill (YLS-10B, Shandong Academy of Medical Sciences, Jinan, China) was used in this study. All groups were suffered with an exhaustion test (25 rpm, medium difficulty coefficient). During the test, the mice stopping running were rapidly shocked by electricity (3 s, 1.5 mA) and the electric shock times were automatically recorded. When the mice rested for 30 seconds or 5 times in 5 minutes after a long time of electric shock, they were judged to be exhausted, and the time to exhaustion was recorded.
2.4 Weight-loaded swimming exhaustion test
The mice loading lead wire on tails (9% body weight) were requested to swim in the water (50 cm × 50 cm × 40 cm, 25 ± 1 °C). The exhaustion time was recorded from the beginning of swimming to the exhaustion time when the head of the mice could not float out of the water for 7 seconds.
2.5 Biochemical analysis
The concentration of glucose, lactic acid, transforming growth factor beta (TGF-β) in serum and glycogen in muscle and liver, as well as the activity of creatine kinase (CK) in serum were measured using assay kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer's instructions. The amounts of hemoglobin, erythrocyte and leucocyte were measured by routine blood test.
2.6 Statistical analysis
All data were expressed as mean ± SEM (standard error of the mean, indicated by error bars). Differences among groups were assessed by one-way analysis of variance (ANOVA), followed by Tukey's post hoc test. Difference between 0 min and 5 min were assessed by Student's t test. Different letters indicated significant differences among groups when p < 0.05.
3. Results and discussion
3.1 Effects of DHA and EPA on the time to exhaustion in mice by consecutive administration
In order to compare the effects of different DHA/EPA formulations on exercise fatigue, the running exhaustion test were performed and the results were shown in Fig. 1. Administration of DHA/EPA–TG and DHA–PL rather than DHA–TG for 10 days significantly extended the time and distance to running exhaustion in mice. Several studies showed that administration of DHA/EPA enriched fish oil for 8–9 weeks could significantly reduce muscle fatigue with the changed fatty acid composition of membrane phospholipids in muscle.20 Importantly, our previous study suggested that administration of different form of DHA/EPA for 4–14 days could significantly increase the level of hepatic and testicular DHA/EPA in mice.21 In the present study, administration of DHA/EPA in different forms for 10 days did alleviate exercise fatigue in mice, which might be attributed to the changes of membrane composition and structure after ω-3 supplementation for 10 days.
 |
| | Fig. 1 Effects of different DHA/EPA formulations on the running time and distance to exhaustion in mice. (A) The time to exhaustion in running test. (B) The distance to exhaustion in running test. All data was presented as mean ± SEM (n = 10). Different letters indicated significant difference at p < 0.05 among groups determined by ANOVA (Tukey's test). | |
Importantly, we found that TG with higher ratio of EPA (EPA:DHA = 34.5:24.7) had better effects on alleviating exercise fatigue than TG with lower ratio of EPA (EPA:DHA = none detected: 42.9) in mice. It seems unreasonable and contradictory that EPA showed a superior effect than DHA on alleviating exercise fatigue as it has been reported that EPA exhibited lower concentration than DHA in vivo.21,22 Several investigations revealed that it was an important reason for biological activity of EPA that not only EPA involved in the conversion to DHA, but also inhibiting inflammation, improving lipid metabolism and so on.23–25 It might imply that EPA could play an important role in the process of substance and energy metabolism, thereby mitigating exercise fatigue, which need to be further confirmed.
Additionally, PL with lower ratio of EPA (EPA:DHA = 10.6:32.6) had a similar effect to TG with higher ratio of EPA (EPA:DHA = 34.5:24.7) on mitigating exercise fatigue in the present study. It might indicate that EPA–PL had a superior effect than EPA–EE. Le et al. reported that a 9 week DHA-PL enriched supplementation improved endurance exercise capacity and skeletal muscle mitochondrial function in adult rats.18 Various investigations have shown that DHA–PL exhibits distinct effects in comparison with DHA–TG/EE. Our previous study found that DHA/EPA–PL was more effective than DHA/EPA–TG/EE on suppressing neuronal apoptosis in mice with Parkinson's diseases.16,26 The efficiency of DHA/EPA–PL is more significant than that of the TG form in alleviating exercise fatigue, which might be attributed to the differences in absorption, distribution and incorporation of the different DHA/EPA formulations in mice. Phospholipids were efficient carriers for ω-3 PUFA in circulation and ω-3 PUFA enriched phospholipids had more preferential incorporation into cell membrane than ω-3 PUFA enriched triglyceride.27,28
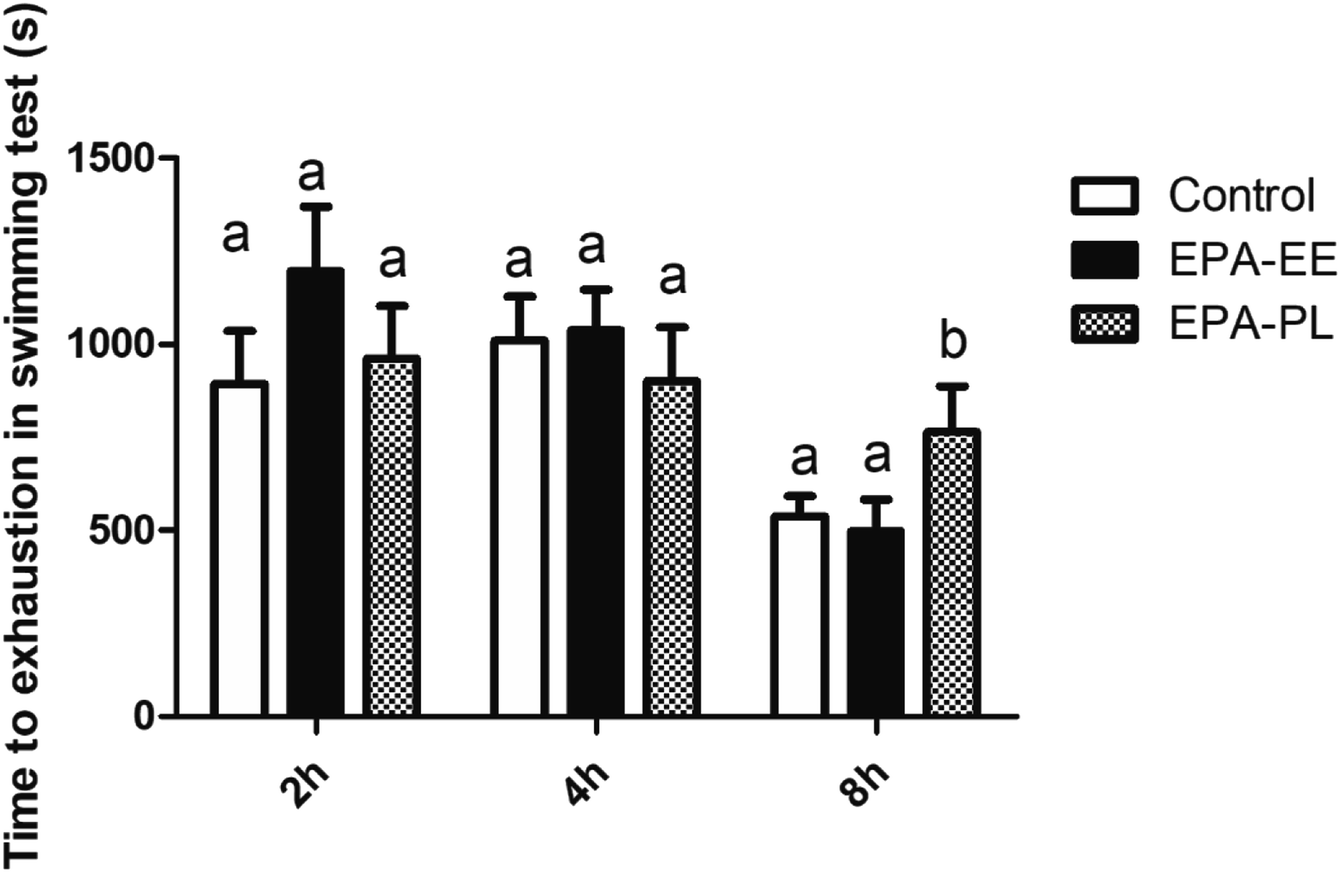
3.2 Rapid effects of EPA–EE and EPA–PL on the time to exhaustion in mice by single administration
Weight-loaded swimming test with a short duration (for minutes, far less than 2 hours) was considered as a common and classical evaluation for exercise endurance. To test the hypothesis that EPA plays a role in the process of substance and energy metabolism, the weight-loaded swimming test was performed to evaluate the rapid effects of EPA-EE and EPA-PL on mitigating exercise fatigue by single administration before changing the fatty acid composition of membrane in muscle. The time to exhaustion of mice was shown in Fig. 2. Although there was no significant difference among the groups at 2nd hour and 4th hour, EPA–PL significantly increased the time to weight-loaded swimming exhaustion at 8th hour, compared with the control and EPA-EE groups. The above results showed that EPA–PL but not EPA–EE had a significant rapid effect on ameliorating exercise fatigue in mice. Jakeman et al. reported that adult male volunteers were immediately supplied with fish oil (750 mg EPA + 50 mg DHA or 150 mg EPA + 100 mg DHA) after muscle injury caused by jumping, and their jumping ability was significantly improved at the 48th hour after intake of fish oil.29 However, the results of this study showed that intake of EPA–EE (1000 mg kg−1 weight) for 8 h did not increase the time to weight-loaded swimming exhaustion in mice (the time at 48 h was not measured), which might be attributed to not only incompletely consistent exercise mode, but also the longer onset time of EPA–EE than EPA–PL. The onset time of EPA–EE/PL might be associated with the rate of digestion and absorption. Dietary lipids are mainly digested in the intestine to form fatty acids and other residual substances, such as monoglycerides or lyso-phospholipids.30 Phospholipids are hydrolyzed into lyso-phospholipids and non-esterified fatty acid (NEFA) by pancreatic lipase during digestion and absorption, while ethyl esters are hydrolyzed into NEFA and ethanol by pancreatic lipase and carboxylesterase. After absorption by intestinal cells, the fatty acid and other substances are re-synthesized into TG and PLs. Importantly, experiments in vitro showed that the rate of hydrolyzing glycerol ester by pancreatic lipase was 10–50 times higher than that of ethyl ester catalyzed by pancreatic lipase and the activity of carboxylesterase was much lower than that of pancreatic lipase.31 For the ethyl ester form, this is a less efficient process than the direct absorption of TG, leading to a worse bioavailability of the ethyl ester form compared with that of the TG form. Notably, the amphiphilic properties of PLs may affect the surface composition of fat droplets to promote the binding of hydrolases before absorption, thus PL form is more easily absorbed than the TG form.
 |
| | Fig. 2 Effects of single intake of different EPA on the weight-loaded swimming time to exhaustion in mice. All data was presented as mean ± SEM (n = 10). Different letters indicated significant difference at p < 0.05 among groups determined by ANOVA (Tukey's test). | |
3.3 Effects of EPA–EE and EPA–PL on energy utilization in mice by single administration
The mice were sacrificed rapidly or 5 minutes after the last weight-loaded swimming exhaustion test. The concentrations of glucose and lactic acid in serum, hepatic glycogen and muscle glycogen were measured and the results were shown in Fig. 3. Compared with the control group, EPA–PL inhibited the decrease of blood glucose and the increase of lactate acid caused by exercise fatigue after weight-loaded swimming exhaustion test, while EPA–EE had no significant effect on glucose content and the accumulation of lactate acid in serum (Fig. 3A and B). EPA–PL significantly increased the levels of glycogen in liver and muscle after weight-loaded swimming exhaustion test but EPA–EE had no beneficial effect (Fig. 3C and D). There was no significant difference between 0 minute and 5 minute after weight-loaded swimming exhaustion test in above parameters. Several results showed that terrestrial phospholipids and nonpolar DHA/EPA significantly inhibited the decrease of blood glucose, glycogen consumption and lactic acid accumulation, and significantly improved exercise fatigue in mice.13,20 The data of biochemical analysis in the present study showed that EPA–PL significantly inhibited the decrease of blood glucose, lactate accumulation and glycogen consumption in liver and muscle, thereby reducing exercise fatigue, which was consistent with the previously reported results. During strenuous exercise, muscle consumes muscle glycogen by glycolysis and produces lactic acid that enters the liver through circulation. Although the majority of lactic acid synthesizes glycogen again in recovery by gluconeogenesis in liver, but the fraction of lactic acid can recycle by gluconeogenesis in liver during exercise. Excessive consumption of muscle glycogen causes excessive accumulation of lactic acid, which inhibit glycolysis and induce acid poisoning in muscle, thereby leading to muscle fatigue. Therefore, promoting Cori cycle (lactic acid circulates in muscle-liver-muscle) is an effective way to eliminate the accumulation of lactic acid in the muscle. The present study showed that single administration of EPA–PL more significantly improved lactic acid recycling rate than EE form. In addition, it might be the reason for reducing glycogen consumption that EPA–PL activated other energy metabolism pathway, such as promoting tricarboxylic acid cycle and lipid metabolism, which could also decrease the production of lactic acid.
 |
| | Fig. 3 Effects of single intake of different EPA on energy utilization in mice weight-loaded swimming test. (A) Glucose, (B) lactic acid, (C) muscle glycogen, (D) liver glycogen. All data was presented as mean ± SEM (n = 10). Different letters indicated significant difference at p < 0.05 among groups determined by ANOVA (Tukey's test). *p < 0.05, significant difference between 0 min and 5 min assessed by Student's t test. | |
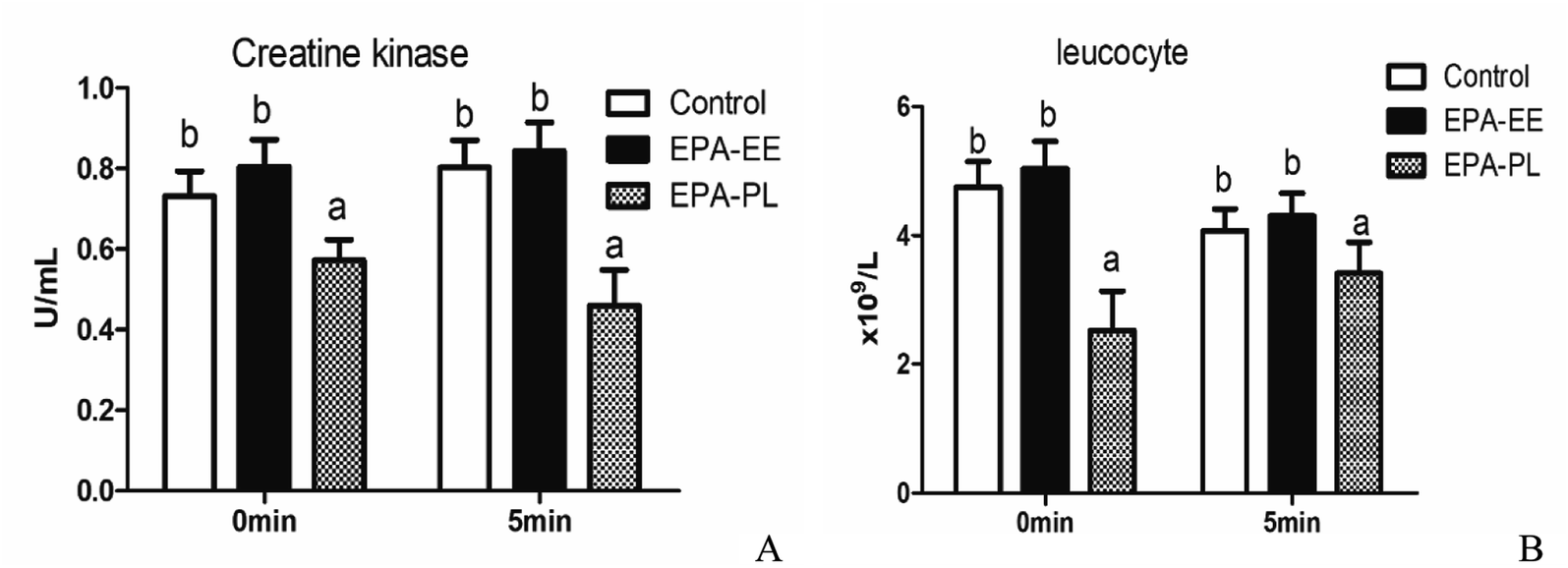
3.4 Effects of EPA–EE and EPA–PL on muscle injury in mice by single administration
The number of leucocyte and the activity of CK in blood were measured to evaluate muscle injury and the results were shown in Fig. 4. Compared with the control group, EPA–PL but not EPA–EE significantly decreased the activity of CK and the amount of leucocyte after weight-loaded swimming exhaustion test. There was no significant difference for CK and leucocyte between 0 minute and 5 minute after weight-loaded swimming exhaustion test. Tinsley et al. showed that fish oil supplementation (6 g day−1, EPA:DHA = 5:1) for one week significantly improved muscle soreness and muscle injury caused by resistance training in adult women.32 Peoples et al. suggested that intake of fish oil for 8 weeks significantly increased muscle strength in male rat using a fatigue model by perfusing the hind limbs of rats with continuous hypoxia and stimulating gastrocnemius–metatarsal–soleus muscle bundle.8 The present study showed that EPA–PL significantly inhibited the activity of CK and the number of leucocytes, suggesting that EPA–PL could alleviate muscle injury induced by weight-loaded swimming exhaustion in mice, which was consistent with the reported results.
 |
| | Fig. 4 Effects of single intake of different EPA on muscle injury in mice weight-loaded swimming test. (A) Creatine kinase, (B) leucocyte. All data was presented as mean ± SEM (n = 10). Different letters indicated significant difference at p < 0.05 among groups determined by ANOVA (Tukey's test). *p < 0.05, significant difference between 0 min and 5 min assessed by Student's t test. | |
3.5 Effects of EPA–EE and EPA–PL on TGF-β in mice by single administration
Increased levels of transforming growth factor β (TGF-β) in circulation and brain lead to the perception of fatigue and reduce autonomous activity. It has been reported that the significantly high level of TGF-β was found in the brain of animals or patients with chronic fatigue syndrome.33 Besides, TGF-β is associated with inflammation. The concentration of TGF-β in serum was measured in mice (Fig. 5). However, exhaustion and single intake of EPA could not affect TGF-β level in the present study. Cao et al. reported that DHA–PL and EPA–PL could suppress the expressions of TGF-β activated kinase 1 in mice with chronic stress.34 Shimojo et al. showed that EPA suppressed the expression of TGF-β in vitro.35 In the present study, short-term exercise fatigue could not affect the level of TGF-β.
 |
| | Fig. 5 Effects of single intake of different EPA on TGF-β in mice weight-loaded swimming test. All data was presented as mean ± SEM (n = 10). Different letters indicated significant difference at p < 0.05 among groups determined by ANOVA (Tukey's test). *p < 0.05, significant difference between 0 min and 5 min assessed by Student's t test. | |
3.6 Effects of EPA–EE and EPA–PL on hemoglobin in mice by single administration
During exercise, oxygen is transported from the respiratory system to muscles and other organs by erythrocyte, and the concentration of hemoglobin reflects carrying oxygen capacity of erythrocyte. The number of erythrocyte and the concentration of hemoglobin were measured and the results were shown in Fig. 6. Unexpectedly, no significant difference among the groups was found in present study, which was presumably due to the short intervention time. Moyers et al. reported that ω-3 fatty acid levels in whole blood were not related to the concentration of hemoglobin in patients with stable coronary artery disease, which could revealed the invalidity of EPA to the concentration of hemoglobin.36
 |
| | Fig. 6 Effects of single intake of different EPA on hemoglobin in mice weight-loaded swimming test. (A) Erythrocyte, (B) hemoglobin. All data was presented as mean ± SEM (n = 10). Different letters indicated significant difference at p < 0.05 among groups determined by ANOVA (Tukey's test). *p < 0.05, significant difference between 0 min and 5 min assessed by Student's t test. | |
4. Conclusions
In the present study, running exhaustion test was used to compare the effects of DHA and EPA on exercise fatigue, and results showed that administration of DHA/EPA–TG and DHA–PL rather than DHA–TG significantly extended the time and distance to running exhaustion in mice. Interestingly, TG with higher ratio of EPA had better effects on alleviating exercise fatigue than TG with lower ratio of EPA in mice. Single administration of EPA–PL but not EPA–EE significantly alleviated exercise fatigue in mice by increasing lactic acid recycling rate as well as inhibiting glycogen consumption and muscle injury, suggesting that EPA–PL exhibited rapid effect on alleviating exercise fatigue (Fig. 7). The study might represent a potential novel candidate or targeted dietary patterns for alleviating exercise fatigue.
 |
| | Fig. 7 The effects of EPA–EE and EPA–PL on alleviating physical fatigue induced by weight-loaded swimming in mice by single administration. | |
Ethics statement
All animal procedures were performed in accordance with the Guidelines for Care and Use of Laboratory Animals of the National Institutes of Health and experiments were approved by the Animal Ethics Committee of experimental animal care at the College of Food Science and Engineering, Ocean University of China (Qingdao, China, approval no. SPXY2015012).
Conflicts of interest
The authors declare no competing financial interest.
Acknowledgements
This work was supported by National Natural Science Foundation of China (31901688), National Key R&D Program of China (2018YFD0901103), Natural Science Youth Foundation of Shandong Province (ZR2019QC004), China Postdoctoral Science Foundation funded project (2017M620294), National Natural Science Foundation of China-Shandong Joint Fund for Marine Science Research Centers (U1606403), the Fundamental Research Funds for the Central Universities (No. 201762028 and 201812017), Laboratory for Marine Drugs and Bioproducts of Pilot National Laboratory for Marine Science and Technology (Qingdao, LMDBKF201807), and Qingdao Postdoctoral Application Foundation funded project, Grant-in-Aid for Scientific Research (No. 16K00890).
References
- G. K. Adler, Exercise and Fatigue—Is Neuroendocrinology an Important Factor?1, J. Clin. Endocrinol. Metab., 2000, 85, 2167–2169 CAS.
- M. Nagai, Y. Morikawa, K. Kitaoka, K. Nakamura, M. Sakurai, M. Nishijo, Y. Hamazaki, S. Maruzeni and H. Nakagawa, Effects of Fatigue on Immune Function in Nurses Performing Shift Work, J. Occup. Health, 2011, 53, 312–319 CrossRef.
- M. B. Reid, Free radicals and muscle fatigue: of ROS, canaries, and the IOC, Free Radicals Biol. Med., 2008, 44, 169–179 CrossRef CAS.
- D. J. McNamara, T. J. Gabbett, G. Naughton, P. Farhart and P. Chapman, Training and competition workloads and fatigue responses of elite junior cricket players, Int. J. Sports Physiol. Perform., 2013, 8, 517–526 Search PubMed.
- L. Lu, F. M. Megahed, R. F. Sesek and L. A. Cavuoto, A survey of the prevalence of fatigue, its precursors and individual coping mechanisms among U.S. manufacturing workers, Appl. Ergon., 2017, 65, 139–151 CrossRef.
- E. Ochi, K. Yanagimoto, T. Morishima and Y. Tsuchiya, Eicosapentaenoic Acid-Rich Fish Oil Supplementation Inhibits the Decrease in Concentric Work Output and Muscle Swelling of the Elbow Flexors, J. Am. Coll. Nutr., 2019, 38, 125–131 CrossRef CAS.
- L. Peter, C. Jillian, H. Kathleen and S. Thomas, Influence of omega-3 (n3) index on performance and wellbeing in young adults after heavy eccentric exercise, J. Sport. Sci. Med., 2014, 13, 151 Search PubMed.
- G. E. Peoples and P. L. McLennan, Dietary fish oil delays hypoxic skeletal muscle fatigue and enhances caffeine-stimulated contractile recovery in the rat in vivo hindlimb, Appl. Physiol., Nutr., Metab., 2017, 42, 613–620 CrossRef CAS.
- A. Saini, A. P. Sharples, N. Al-Shanti and C. E. Stewart, Omega-3 fatty acid EPA improves regenerative capacity of mouse skeletal muscle cells exposed to saturated fat and inflammation, Biogerontology, 2017, 18, 109–129 CrossRef CAS.
- T. D. Mickleborough, Omega-3 polyunsaturated fatty acids in physical performance optimization, Int. J. Sport Nutr. Exercise Metab., 2013, 23, 83–96 CAS.
- X. F. Guo, A. J. Sinclair, G. Kaur and D. Li, Differential effects of EPA, DPA and DHA on cardio-metabolic risk factors in high-fat diet fed mice, Prostaglandins, Leukotrienes Essent. Fatty Acids, 2017, 136, 47–55 CrossRef.
- J. G. Martins, EPA but not DHA appears to be responsible for the efficacy of omega-3 long chain polyunsaturated fatty acid supplementation in depression: evidence from a meta-analysis of randomized controlled trials, J. Am. Coll. Nutr., 2009, 28, 525–542 CrossRef CAS PubMed.
- R. Jäger, M. Purpura and M. Kingsley, Phospholipids and sports performance, J. Int. Soc. Sports Nutr., 2007, 4, 5 CrossRef.
- A. Hirose, M. Terauchi, Y. Osaka, M. Akiyoshi, K. Kato and N. Miyasaka, Effect of soy lecithin on fatigue and menopausal symptoms in middle-aged women: a randomized, double-blind, placebo-controlled study, Nutr. J., 2018, 17, 4 CrossRef.
- T.-T. Zhang, J. Xu, Y.-M. Wang and C.-H. Xue, Health benefits of dietary marine DHA/EPA-enriched glycerophospholipids, Prog. Lipid Res., 2019, 75, 100997 CrossRef CAS.
- C. Wang, D. Wang, J. Xu, T. Yanagita, C. Xue, T. Zhang and Y. Wang, DHA enriched phospholipids with different polar groups (PC and PS) had different improvements on MPTP-induced mice with Parkinson's disease, J. Funct. Foods, 2018, 45, 417–426 CrossRef CAS.
- C. C. Wang, Y. Guo, M. M. Zhou, C. H. Xue, Y. G. Chang, T. T. Zhang and Y. M. Wang, Comparative studies of DHA-enriched phosphatidylcholine and recombination of DHA-ethyl ester with egg phosphatidylcholine on ameliorating memory and cognitive deficiency in SAMP8 mice, Food Funct., 2019, 10, 938–950 RSC.
- G. M. Le, V. Chaté, I. Hininger-Favier, B. Laillet, B. Morio, G. Pieroni, U. Schlattner, C. Pison and H. Dubouchaud, A 9-wk docosahexaenoic acid-enriched supplementation improves endurance exercise capacity and skeletal muscle mitochondrial function in adult rats, Am. J. Physiol. Endocrinol. Metab., 2016, 310, E213 CrossRef.
- J. Folch, M. Lees and G. H. Sloane Stanley, A simple method for the isolation and purification of total lipides from animal tissues, J. Biol. Chem., 1957, 226, 497–509 CAS.
- G. E. Peoples and P. L. McLennan, Long-chain n-3 DHA reduces the extent of skeletal muscle fatigue in the rat in vivo hindlimb model, Br. J. Nutr., 2014, 111, 996–1003 CrossRef CAS.
- D. D. Wang, F. Wu, M. Wen, L. Ding, L. Du, C. H. Xue, J. Xu and Y. M. Wang, Replenishment of Docosahexaenoic Acid (DHA) in Dietary n-3-Deficient Mice Fed DHA in Triglycerides or Phosphatidylcholines After Weaning, J. Food Sci., 2018, 83, 481–488 CrossRef CAS.
- F. Wu, D. D. Wang, M. Wen, H. X. Che, C. H. Xue, T. Yanagita, T. T. Zhang and Y. M. Wang, Comparative analyses of DHA-Phosphatidylcholine and recombination of DHA-Triglyceride with Egg-Phosphatidylcholine or Glycerylphosphorylcholine on DHA repletion in n-3 deficient mice, Lipids Health Dis., 2017, 16, 234 CrossRef PubMed.
- C. T. Chen and R. P. Bazinet, β-oxidation and rapid metabolism, but not uptake regulate brain eicosapentaenoic acid levels, Prostaglandins, Leukotrienes Essent. Fatty Acids, 2015, 92, 33–40 CrossRef CAS.
- C. Song, A. G. Phillips, B. E. Leonard and D. F. Horrobin, Ethyl-eicosapentaenoic acid ingestion prevents corticosterone-mediated memory impairment induced by central administration of interleukin-1β in rats, Mol. Psychiatry, 2003, 9, 630 CrossRef.
- L. Ding, L. Zhang, M. Wen, H. Che, L. Du, J. Wang, C. Xue, J. Xu and Y. Wang, Eicosapentaenoic acid-enriched phospholipids improve atherosclerosis by mediating cholesterol metabolism, J. Funct. Foods, 2017, 32, 90–97 CrossRef CAS.
- C.-C. Wang, D. Wang, T.-T. Zhang, T. Yanagita, C.-H. Xue, Y.-G. Chang and Y.-M. Wang, A comparative study about EPA-PL and EPA-EE on ameliorating behavioral deficits in MPTP-induced mice with Parkinson's disease by suppressing oxidative stress and apoptosis, J. Funct. Foods, 2018, 50, 8–17 CrossRef CAS.
- M. Lagarde, N. Bernoud, N. Brossard, D. Lemaitre-Delaunay, F. Thies, M. Croset and J. Lecerf, Lysophosphatidylcholine as a preferred carrier form of docosahexaenoic acid to the brain, J. Mol. Neurosci. MN, 2001, 16, 201–204 CrossRef CAS ; discussion 215–221.
- F. Thies, C. Pillon, P. Moliere, M. Lagarde and J. Lecerf, Preferential incorporation of sn-2 lysoPC DHA over unesterified DHA in the young rat brain, Am. J. Physiol., 1994, 267, R1273–R1279 CAS.
- J. R. Jakeman, D. M. Lambrick, B. Wooley, J. A. Babraj and J. A. Faulkner, Effect of an acute dose of omega-3 fish oil following exercise-induced muscle damage, Eur. J. Appl. Physiol., 2017, 117, 575–582 CrossRef CAS.
- G. E. Lidgerwood, S. M. Pitson, C. Bonder and A. Pébay, Roles of lysophosphatidic acid and sphingosine-1-phosphate in stem cell biology, Prog. Lipid Res., 2018, 72, 42–54 CrossRef CAS.
- L. Y. Yang, A. Kuksis and J. J. Myher, Lipolysis of menhaden oil triacylglycerols and the corresponding fatty acid alkyl esters by pancreatic lipase in vitro: a reexamination, J. Lipid Res., 1990, 31, 137–147 CAS.
- G. M. Tinsley, J. J. Gann, S. R. Huber, T. L. Andre, P. M. La Bounty, R. G. Bowden, P. M. Gordon and P. W. Grandjean, Effects of fish oil supplementation on postresistance exercise muscle soreness, J. Diet. Suppl., 2017, 14, 89–100 CrossRef CAS.
- M. Arai, H. Yamazaki, K. Inoue and T. Fushiki, Effects of intracranial injection of transforming growth factor-β relevant to central fatigue on the waking electroencephalogram of rats: comparison with effects of exercise, Prog. Neuro-Psychopharmacol. Biol.
Psychiatry, 2002, 26, 307–312 CrossRef CAS.
- W. Cao, C. Wang, Y. Chin, X. Chen, Y. Gao, S. Yuan, C. Xue, Y. Wang and Q. Tang, DHA-phospholipids (DHA-PL) and EPA-phospholipids (EPA-PL) prevent intestinal dysfunction induced by chronic stress, Food Funct., 2018, 10, 277–288 RSC.
- N. Shimojo, S. Jesmin, S. Zaedi, S. Maeda, M. Soma, K. Aonuma, I. Yamaguchi and T. Miyauchi, Eicosapentaenoic acid prevents endothelin-1-induced cardiomyocyte hypertrophy in vitro through the suppression of TGF-β1 and phosphorylated JNK, Am. J. Physiol.: Heart Circ. Physiol., 2006, 291, H835–H845 CrossRef CAS.
- B. Moyers, R. Farzaneh-Far, W. S. Harris, S. Garg, B. Na and M. A. Whooley, Relation of Whole Blood n-3 Fatty Acid Levels to Exercise Parameters in Patients With Stable Coronary Artery Disease (From the Heart and Soul Study), Am. J. Cardiol., 2011, 107, 1149–1154 CrossRef CAS.
|
| This journal is © The Royal Society of Chemistry 2019 |
Click here to see how this site uses Cookies. View our privacy policy here. 
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence *a and
Yu-Ming Wang
*a and
Yu-Ming Wang