
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Tools and rules for modelling uptake and bioaccumulation of nanomaterials in invertebrate organisms†
Nico W.
van den Brink
 *a,
Anita
Jemec Kokalj
b,
Patricia V.
Silva
c,
Elma
Lahive
d,
Karin
Norrfors
e,
Marta
Baccaro
a,
Zahra
Khodaparast
c,
Susana
Loureiro
c,
Damjana
Drobne
b,
Geert
Cornelis
e,
Steve
Lofts
f,
Richard D.
Handy
g,
Claus
Svendsen
d,
Dave
Spurgeon
d and
Cornelis A. M.
van Gestel
h
*a,
Anita
Jemec Kokalj
b,
Patricia V.
Silva
c,
Elma
Lahive
d,
Karin
Norrfors
e,
Marta
Baccaro
a,
Zahra
Khodaparast
c,
Susana
Loureiro
c,
Damjana
Drobne
b,
Geert
Cornelis
e,
Steve
Lofts
f,
Richard D.
Handy
g,
Claus
Svendsen
d,
Dave
Spurgeon
d and
Cornelis A. M.
van Gestel
h
aDepartment of Toxicology, Wageningen University, Wageningen, The Netherlands. E-mail: nico.vandenbrink@wur.nl
bDepartment of Biology, Biotechnical Faculty, University of Ljubljana, Ljubljana, Slovenia
cDepartment of Biology and CESAM, University of Aveiro, Aveiro, Portugal
dCentre of Ecology and Hydrology (CEH-NERC), Wallingford, UK
eDepartment of Soil and Environment, Swedish University of Agricultural Sciences, Uppsala, Sweden
fCentre for Ecology and Hydrology, Lancaster, UK
gDepartment of Biological and Marine Sciences, Plymouth University, Plymouth, UK
hDepartment of Ecological Science, Faculty of Science, Vrije Universiteit Amsterdam, Amsterdam, The Netherlands
First published on 14th May 2019
Abstract
Quantification of the uptake and elimination of nanomaterials (NMs) by organisms is key in assessing the environmental risks of NMs. For this, uptake models for conventional solutes may be used, although no consensus exists on their applicability for NMs. In this critical review therefore, conventional modelling approaches are scrutinised for their applicability for NMs. Statically derived accumulation factors, like BCF or BAF based on measured concentrations, are considered to be flawed because NMs are thermodynamically not stable, an important assumption for this approach. Dynamically derived accumulation factors, based on kinetic exposure experiments, may be applicable because no equilibrium between the organism and exposure medium is needed. Currently there is no full understanding of the passive uptake of NMs, which hampers assessment of the applicability of biotic ligand models. Passive uptake, however, is generally considered to be very limited, which would imply a limited applicability of BLMs for NMs. Physiologically based pharmacokinetic (PBPK) models, or biodynamic models, have successfully been applied in uptake studies with NMs. Their underlying assumptions can be met in experiments addressing NMs and case studies presented in this review demonstrate their applicability to model NM-form specific kinetics, integrated with environmental fate models, including relevant physiological processes. Their application requires the a priori definition of the major mechanisms driving the uptake kinetics and the quantification of the associated kinetic rate constants. This limits their application to those mechanisms for which the kinetic rate constants can actually be quantified. Within these limitations, PBPK models have been shown to be applicable and provide a promising general approach to improve modelling of NM-accumulation in organisms.
Environmental significanceIn this paper, we critically review approaches in modelling the uptake of nanomaterials by biota under environmentally relevant conditions. Existing modelling approaches, developed for the uptake of solutes by organisms, are evaluated within the context of the underlying assumptions and applicability to nanomaterials. Guidance for the selection of modelling approaches is provided, which will enhance the applicability of uptake modelling in environmental risk assessment of nanomaterials. |
Introduction
Environmental risk assessment requires information both on the level of exposure and the hazards that chemicals pose to organisms. The environmental risk assessment paradigm can be applied to nanomaterials (NMs), but the challenges include uncertainty about actual forms and environmental concentrations of NMs and exposure modelling, as well as the difficulty of determining which facets of the physico-chemical properties of NMs are the most important to the hazard.1 To date, most of the research on environmental risks of NMs has focussed on the characterisation and quantification of their hazards, using standard toxicity assays or slightly adapted procedures to cope with the special properties of NMs.2,3 Dose–response relationships may be derived from nominal exposure concentrations, although the use of measured concentrations is preferred in ecotoxicity testing with NMs. Generally, studies characterise NM-properties in the spiking medium4–7 to at least confirm that the expected dose was present, even if the precise form of the NMs in the exposure could have changed. Quantification of the actual form of a NM taken up by organisms is still in its infancy, and the biological matrices present many challenges to NM detection inside organisms. Most uptake or accumulation studies report either qualitative or semi-quantitative data to infer the presence of NMs in tissues, such as electron microscopy observations8 and synchrotron X-ray fluorescence analysis for fluorescent NMs,9 or use total concentrations, e.g. in the case of metal-based NMs.10–12 Only recently, some studies have appeared in which different forms of NMs were reported in tissues of organisms, providing insights into form-specific uptake of NMs.13–16 Methods to quantify form-specific concentrations of NMs such as single particle inductively coupled plasma mass spectroscopy (sp-ICP-MS) for metal-based NMs are becoming more available. These developments in analytical methods have enabled modelling approaches that could reveal patterns of form-specific uptake and accumulation of NMs in organisms, such as aspects of absorption, distribution, metabolism and excretion (ADME). This also provides the possibility to predict form-specific uptake of NMs in organisms under environmentally relevant conditions.17,18Several authors have applied modelling approaches that are commonly used in studies of conventional chemicals to model NM uptake in different terrestrial and aquatic organisms.19–22 The models used for conventional chemicals were developed specifically to address the uptake of solutes such as metals or lipophilic organic compounds and were not intended for NMs. The solute chemistry assumptions underlying those models may therefore not be met when applying them to NMs.23 For instance, fugacity models, used to quantify the accumulation kinetics of non-polar organic compounds, assume equilibrium partitioning which is disputed for NMs.24 Currently, however, there is no consensus on the pros and cons of different accumulation models and whether or not they are applicable to NMs, or on how to select the most appropriate model for specific cases of NMs.
In light of the demands from the perspective of the environmental risk assessment of NMs, this knowledge gap needs to be addressed, hence, in this review we critically review modelling approaches used for conventional chemicals, which may be applicable for describing the uptake and accumulation of NMs in soil and aquatic invertebrates. Invertebrates were selected because they comprise more than 99% of all animals; they are the most diverse group of organisms present on Earth and are integral to several ecological functions (e.g. soil structure and maintenance, nutrient cycling) which link to ecosystem services.25–27 The review will be restricted to whole body accumulation in invertebrate species and not focus on tissue-specific uptake patterns and internal distribution. This is because the literature on form- and tissue-specific uptake of NM and their internal distribution in environmental organisms is currently too limited for a proper review. First, we will provide an overview of different routes of uptake of NMs and their fate in different invertebrate species. Based on this, different approaches used for modelling of the uptake of conventional chemicals will be discussed with respect to their application to NMs. We will address basic assumptions underlying the different modelling approaches that need to be met and will critically discuss the literature using such approaches for NM accumulation in selected species. Finally, we will provide an outlook for future research on modelling approaches and guidance on the applicability of existing modelling approaches to NMs.
Routes of uptake and elimination of NMs in invertebrates
Invertebrates can potentially accumulate NMs actively via ingestion and consecutive uptake across the epithelium in the body and to a lesser extent by anal uptake, or passively via uptake through body surfaces or body openings (Fig. 1). Feeding strategies of invertebrates vary considerably, but still they can be classified into different functional feeding groups, i.e. shredders (chew conditioned food e.g. litter, plants or wood), filter feeders (suspension feeding), gathering collectors (deposit feeders: they ingest sediment and/or soil), scrapers (graze food attached to surfaces), and predators (engulf prey, ingest body fluids).28–30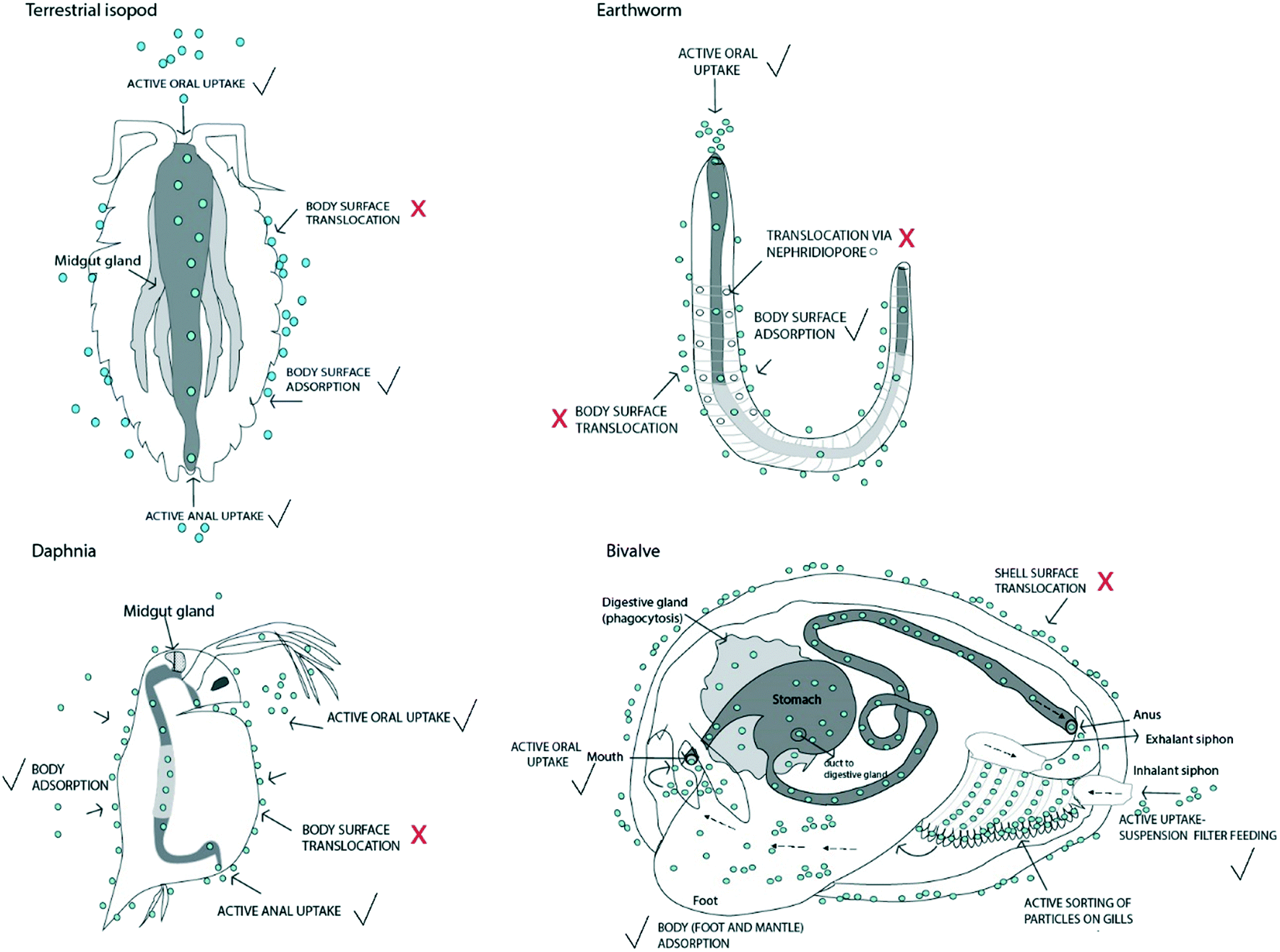 | ||
| Fig. 1 Active and passive uptake of nanomaterials in different representative invertebrate species commonly used in environmental research: terrestrial isopod (Crustacea), earthworm (Annelida), water flea (Crustacea), and bivalve (Mollusca; e.g. Mytilus sp.; only one valve with a removed mantle is shown). The organisms are presented schematically, as extracted from Brusca et al. (2016)30 and Ruppert et al. (2004).25 The gut is shown as a grey tube. Light grey represents the midgut, while the foregut and hindgut regions are dark grey. The position and length of each gut region are not shown at realistic scales. Nanomaterials are shown as blue circles. Routes where NM passage is questionable are marked with a question mark and a red cross. Probable uptake routes are marked with a black tick symbol. | ||
Depending on the environmental fate of NMs, feeding groups may be differentially exposed to NMs. In aquatic environments, suspension feeders for instance will be exposed predominantly to waterborne NMs while deposit feeders will be exposed mainly to NMs following sedimentation, although the potential for waterborne exposure cannot be excluded. Shredders, scrapers and predators can also be exposed to both, depending on whether they are pelagic or benthic invertebrates. In terrestrial environments, deposit feeders and shredders are probably in first contact with NMs followed by predators, although in cases NMs are available to other soil invertebrates these may also directly be exposed. Aging of NMs in the environment, including transformation processes such as aggregation, sulfidation and dissolution into ions, influences the environmental fate of NMs, and as such their availability for the different groups of organisms. For instance, aggregation of NMs will increase sedimentation in aquatic systems, likely increasing the exposure of deposit feeders. There is some evidence that uptake of metals from NMs (e.g. ZnO) is through the dissolution of ions from the NMs to the pore water of soil which is then available for uptake by organisms.21,31 Laycock et al. (2016) found that the dermal uptake rates of ZnO NMs based on pore water were comparable to those based on oral and dermal uptake from soil, and it was concluded that the dominant uptake route of Zn was through the dermis from pore water exposure.31 Other transformations, such as sulfidation (e.g. Ag to Ag2S) usually making NMs less reactive, have also started to be considered in bioavailability kinetic studies in order to reflect more environmentally realistic exposure scenarios.14,32 These studies have concluded that the uptake of Ag from Ag2S NMs is slower compared to pristine Ag NMs which supports the assertion that uptake is largely driven by dissolution of ions from NMs.14 Uptake of ions from metal based NMs may be via the usual uptake pathways for solutes (i.e. facilitated uptake through ion channels or active transporters or even electroneutral diffusion,33 or by endocytosis34,35). However, particulate materials from NMs may also be accumulated, as was shown for coelomocytes from earthworm36 and in snails.34 This endocytosis was shown to be (partly) active, as was indicated by the application of inhibitors of different receptor mediated endocytosis pathways (including clathrin and caveolin mediated uptake). The corona of NMs may change during ageing, depending on the environment they are in, which may affect the interactions of NMs with organisms,37 including the potential receptor mediated endocytosis of the NMs.
A second potential route of active NM uptake is through anal intake of water. This was shown for terrestrial isopods that have a water-conducting system on the dorsal outside part of their body surface that collects the excreted urine that is reabsorbed by the gut to regain the lost water.38–40 Anal uptake of water has also been demonstrated for some other crustaceans, such as Daphnia magna, commonly used in ecotoxicity testing39 and is common in sea cucumbers.30
Some NMs that are lipid soluble, such as pristine C60, have been demonstrated to move across biological membranes by diffusion, similar to some organic chemicals. However, this does not mean such NMs would also cross the integument of invertebrates by passive uptake processes. The integument is a considerable barrier, often with several anatomical layers and many biological ligands. The structure of such invertebrate integuments, however, varies considerably,30 ranging from lacking a cuticle (Platyhelminthes) to being lined with different types of cuticles. Such a lining can be composed of sclerotized proteins without chitin (Annelida, Polychaeta, Mollusca), highly cross-linked collagens and specialised insoluble proteins (“cuticulins”), glycoproteins or lipids (Nematoda), or of multi-layered chitin (Arthropoda).25,30 The cuticle enables the invasion of hostile environments by the organisms, such as dry terrestrial soils and digestive tracts of hosts, because it drastically reduces the permeability of the body wall, also for NMs. Passive translocation of NMs through any of the invertebrate integuments is therefore very unlikely if the barrier is intact. Paracellular diffusion of NMs in between epithelial cells is also unlikely for most species, either because the epithelium is electrically tight (i.e. high resistance, low permeability) or because the presence of bivalent ions (especially Ca2+ and Mg2+) and the protein extracellular matrix in the paracellular space would promote rapid aggregation of NMs. However, in a few cases of soft-bodied invertebrates, dermal paracellular uptake of Ag-NMs has been suggested e.g. in earthworms.10 Regardless, in the case of a damaged integument or erosion of the external cuticle when present, NMs may be able to penetrate the tissues. Additionally, dermal uptake of solutes like metal ions resulting from metal NMs may be important in soft bodied species like for instance earthworms.41
NMs may also enter the body passively via openings. Besides the mouth and anus, involved in active uptake, probable body openings that could act as an entrance point for NMs are those related to excretion and gas exchange. Most of the invertebrates possess gills that enable gas exchange. For example, in bivalves, the water effluent runs via the inhalant siphon and the particles are directed to the mouth via active sorting on the gills (Fig. 1). In the case of terrestrial species, such entrance routes do not exist because gas exchange surfaces are internalised to prevent water loss and are connected to the external environment via openings that may have a special mechanism of closure. However, the respiratory system is regarded as very tight to solutes and much less permeable than the gut of animals. In some insects, such as collembolans, a potential entry point of NMs could be the ventral tube, which is the main site of water and salt exchange. This organ enables the absorption of water from surfaces under relatively dry conditions if no water is available for drinking.42,43
Besides actual uptake (i.e. body internalisation), NMs may also be adsorbed onto the body surfaces of organisms, which has been shown for some invertebrate species, for example the brine shrimp Artemia salina44 and the water flea D. magna.45,46 Whether the structure of the invertebrate body surfaces affects the extent of NM adsorption remains to be investigated. Although surface adsorbed NMs have not crossed epithelia tissues, and are thus not internalised by organisms, they should be considered in case of potential transfer of NMs to higher trophic levels.47 This also accounts for NMs in the gut, which are not crossing the gut epithelium, as has been observed often e.g. for daphnid species.48,49
Storage and elimination
Once taken up by organisms, NMs can be retained in the body or excreted.50 The processes that a species may adopt will depend on the exposure route and concentration, as well as the detoxification strategies available to the animal. The fate of NMs in the body will depend on the material NMs are made of and the transformations they may have undergone while ageing. For metal-containing NMs that dissolve, it is possible for the free metal ion to be taken up and then subsequently incorporated into a metal storage granule inside the organism. However, whether or not internalised particulate NMs can be added to those storage pathways or act as an initiating ‘seed’ to form a new storage granule is unclear. Several studies have reported intracellular compartmentalization as an important mechanism to minimise the toxicity of NMs in aquatic and terrestrial invertebrates. NMs and derived materials may accumulate in phagocytic macrophages, e.g. in digestive tissues (i.e., gut epithelia, digestive glands) and are thus not distributed in the whole body.9,50–52 Numerous electron-dense granules and vacuoles have been observed in the TEM images of the midgut cells of the cutworm Spodoptera litura exposed to ionic zinc.53 The number of dense granules and vacuoles and their size correlated with the accumulation of Zn in the midgut. In isopods, the digestive gland or hepatopancreas has been reported as the storage compartments for the bioaccumulation of Ag from Ag-NMs and AgNO3 (ref. 9) or of Co from CoFe2O4 NMs.54 Earthworms have the ability to store metals in the chloragogenous tissue, which contains phosphate, calcium and sulphur.55 In a toxicokinetics study with silver exposure (as nanoparticles and as ions) of the earthworm Lumbricus rubellus, it was indicated that Ag-NMs may have a specific pathway for uptake, detoxification, and excretion via the gut wall, liver-like chloragogenous tissue, and nephridia.21The rate of elimination of metals and NMs from organisms has been demonstrated to be predominantly influenced by the organism's physiology, while other parameters such as medium concentrations and characteristics of the NMs56 or the exposure route9,21 are assumed not to have major impacts in this regard. Therefore, physiological pathways, such as carrier systems (including the ability to exocytose material by e.g. macrophages), may assist to depurate NMs. Several different processes may be involved in the elimination strategies of nanomaterials among aquatic and terrestrial invertebrates.57 Excretion via faeces may help to discharge and detoxify metal-based nanomaterials by invertebrates, although it is likely that most of the NMs in faeces are transient materials not taken up by the organism. The ability of digestive cell vesicles to store metals and then release them either into the lumen of the alimentary canal or into the midgut gland is another process that chemical compounds can undergo to be finally discharged by faeces.58 However, coprophagic organisms like terrestrial isopods may re-ingest these discharged materials through the uptake of faeces.
The kinetics of uptake and elimination of metal-based NMs, or derived metal ions, vary among organisms and determine their accumulation patterns.59 Several processes of accumulation can be described for ionic forms of trace metals in aquatic invertebrates. For instance, the Zn body concentration is generally levelled by matching the Zn excretion rate with the uptake rate. Barnacles, however, showed a different strategy, as they were able to store Zn in a detoxified form in granules, which is hardly eliminated because these granules do not have access to ducts leading to excretion. Some invertebrates detoxify Cd by binding to metallothionein.60 Other organisms can excrete metals from the metabolic available pool, such as P. elegans which can excrete Cu,61 while other species may excrete metals from the detoxified store. For instance, the amphipod crustacean Orchestia gammarellus has the ability to store Cu taken up from food in Cu-rich detoxified granules of ventral caeca cells and release them into the gut lumen. It should be noted, however, that most of the mentioned studies are based on ionic forms of the metals involved. Little information is available on the exact kinetic processes of NMs in invertebrates, which points to an important gap in understanding these phenomena for NMs.
The uptake and elimination kinetics of metal NMs may also be form-dependent, meaning that the same organism can use different uptake and depuration pathways for e.g. NMs and ions.57 For example, different uptake and elimination strategies have been established for Ag-NMs and ionic Ag in the estuarine polychaete Nereis diversicolor.8 Ag-NMs were mainly associated with inorganic granules, organelles, and heat denatured proteins, while ionic Ag was associated predominantly with metallothionein. N. diversicolor phagocytoses particles in the digestive system; therefore, the fate of particulate materials differs from that of the ionic form. Different accumulation patterns of Ag-NMs and ionic Ag in the estuarine snail Peringia ulvae have also been reported, mainly due to the lower Ag-NM uptake and different effluxes of Ag-NMs and ionic Ag.62 In that study, the elimination of Ag-NMs occurred in two phases: faster efflux of Ag-NMs followed by slower efflux of dissolved Ag. In the earthworm L. rubellus, however, no differences were detected in the fraction to which Ag was associated, the metal-rich granules being the most important for both ionic and particulate forms.13 In Daphnia magna, CuO-NMs were localized in the gut lumen, with no indication of being internalized in the cells, and the NMs were quickly eliminated from the body.63 In another study, Au-NMs were also retained in the gut lumen of D. magna and also no internalization in cells was observed.64 The authors used a two-compartment model to describe Au-NM elimination, since it was revealed to be bi-phasic, with a fast elimination rate in the first hour followed by a slower elimination rate. D. magna exposed to both Ag-NMs and AgNO3 through different exposure routes (contamination through water only, food only and both water and food) showed a generally lower elimination rate for Ag-NMs compared to AgNO3, indicating that Ag from Ag-NM exposure was possibly more difficult to depurate than Ag from AgNO3.65 Additionally, the inert fraction, not excreted from the organism, obtained through kinetics modelling showed that for Ag-NMs this fraction was higher than for ionic Ag.
Biomineralization is the process by which organisms use minerals to support existing tissues, which can work as a sequestration strategy for metals, for instance by incorporating them into exoskeletons and relatively inert shells.57 Zn, Cd and Cu excretion during the larval development of the midge Chironomus riparius was associated with moulting and metamorphosis (exuviae).66 There is, however, a need for more studies on the possibility of elimination of metals in NMs and materials from NMs through biomineralization and storage in shells or via moults of organisms, and on the potential recirculation of materials in organisms feeding on exuviae. Another strategy for excretion of materials was found in Mytilus galloprovincialis exposed to metal oxide NMs, showing the ability to repackage CeO2-NMs in pseudo faeces and excrete them.67
Modelling of uptake and elimination
Here we will review those modelling approaches most widely used to describe and analyse the uptake and accumulation patterns of conventional chemicals in organisms, i.e. biotic ligand models (BLMs), accumulation factors and physiologically based pharmacokinetic models (PBPK models) or biodynamic models. Their applicability for NMs will be scrutinised, after which some case studies will be presented which were successful in applying accumulation models to NMs.Biotic ligand models
Biotic ligand models (BLMs) predict the bioavailability and toxicity for metal exposure via direct, passive uptake, assuming reversible, equilibrium binding between metal ions and receptor sites in the organism (the biotic ligands) (Di Toro et al., 2001 (ref. 81)). Metal cations compete with other cations for binding to biotic ligands, while interactions with solution ligands (such as natural organic matter) result in complexation, decreasing the bioavailability of the metals. Several studies have applied BLM approaches to predict the accumulation patterns and effects of metals in the ionic form, including cadmium, nickel and copper in soil organisms and plants.68,69In assessing whether BLM-type approaches are suitable and/or necessary for modelling passive uptake of NMs, an understanding of why such an approach is needed for ionic metals is useful. The need for a BLM-type approach for ionic metals is derived from the facts that (i) uptake and toxicity are not simple functions of the total concentration of the metal to which the organism is exposed, but rather are functions of the activities of one or more specific metal species (usually the free metal ion), and (ii) uptake and toxicity are not functions of the potential free ion only, but also of competing ions in the exposure medium. Furthermore, the uptake mechanism must meet the mechanistic requirements of the BLM, i.e. rapid, reversible association with receptor sites on the organism as the first step in internalisation leading to toxic effects. Some studies22,70 have applied the Michaelis–Menten approach to quantify the uptake rates of NMs to organisms in short-term exposures. Such modelling derives the receptor concentration and binding affinity parameters and could thus be argued to be a BLM-type approach. A more recent work concluded that BLM approaches only predict the acute toxicity of Ag in case the uptake was predominantly via the ionic dissolved form of Ag rather than the NM forms.20 The potential applicability of the BLM is further confounded by the relative roles of dietary uptake versus direct uptake, as Khan et al., 2014 (ref. 64) showed for D. magna. Overall, the results indicate that BLM approaches currently seem less applicable to model the uptake and toxicity of NMs compared to ionic metals, especially over longer periods of time. The BLM will remain of importance in modelling the uptake and toxicity of metals dissolving from NMs during exposure. More research is needed to understand the mechanisms by which NMs are passively taken up by invertebrates and the extent to which this uptake satisfies the requirements of BLM theory, before making more definitive statements on the applicability of the BLM for NMs. This includes the potential for differential uptake of different NM ‘species’ (e.g. with different extents of a particular coating type) and whether components of the exposure medium can compete with NMs for uptake.
Accumulation factors and equilibrium partitioning models
Accumulation factors are the simplest way of describing the uptake of chemicals in organisms. Accumulation factors are defined as ratios between concentrations in the organism and in the surrounding water in the case of aquatic organisms (bioconcentration factor; BCF), in the surrounding soil/sediment in the case of soil/sediment organisms (biota to soil/sediment accumulation factor; BSAF), or in the food (biomagnification factor; BMF). When calculating BCFs and BSAFs from measured concentrations in different matrices, it is assumed that the concentrations in the organisms and the surrounding media are in equilibrium with each other. In a study on the aquatic uptake of TiO2 NMs in nematodes, it was shown that the ratio between titanium concentrations in organisms and water was clearly dependent on water concentrations.6 For the BCF/BSAF concept to be suitable for risk assessment, BCF/BSAF values should be independent of the exposure concentration. However, also for other metals, it has been shown that BCFs and BSAFs tended to be highest at lower exposure concentrations and decrease with increasing exposure level for different metal-salts,71 hence it was concluded that the BCF/BSAF concept may not always be applicable for metal uptake. Similarly, Praetorius et al. (2014) discussed the misconception of using BCFs based on measured concentrations of NMs in organisms and in water/soil, stating that due to the fact that NMs in the environment are in thermodynamically unstable forms they cannot be in equilibrium between two compartments.24 Based on this, is was concluded that the equilibrium partitioning concept was not valid for NMs and that bioaccumulation factors (BCF or BSAF) for NMs cannot be derived from measured concentrations in organisms and other media. With respect to accumulation and BCFs/BSAFs, it needs to be assured that nanoparticles are actually internalised in the tissues of the organisms. For BMFs, no equilibrium is assumed, since the flow of material is one way from the diet item to the consumer, and feeding may occur in discrete events. Nevertheless, a steady state is assumed, which will only be the case under static test conditions.Accumulation factors (BCF/BSAF) can also be derived dynamically, based on kinetic experiments that aim at quantifying uptake and elimination rate constants (for details see later discussion on PBPK/biodynamic models). Different types of toxicokinetics models or biodynamic models72 can be developed, which incorporate specific processes with respect to accumulation, distribution, metabolism and excretion (ADME) of chemicals,73 which may also be applicable to nanomaterials.70 The simplest model only considers uptake and elimination in a one compartment model. According to this model, the uptake phase can be modelled by eqn (1) including both uptake and elimination when exposed.
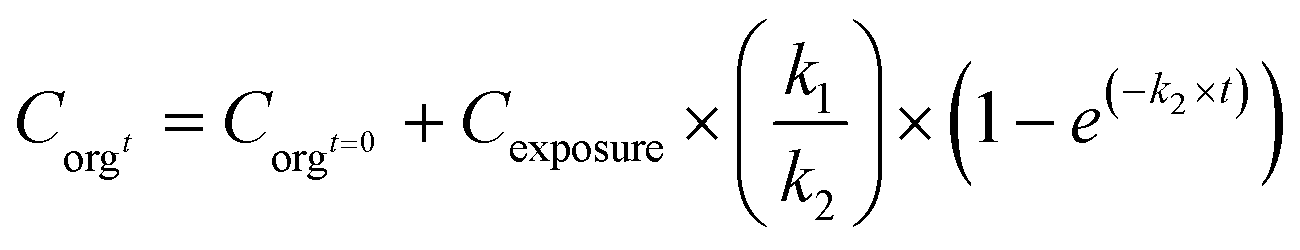 | (1) |
This model assumes a constant exposure concentration. A steady state under stable conditions will be reached after a certain, usually fairly long period of time, resulting in a BCF/BSAF that equals k1/k2 (at longer time periods the parameter e(−k2×t) will reach 0). In this way the BCF/BSAF can be derived dynamically.24 Such dynamic assessment of accumulation factors does not require the establishment of a steady state between compartments (k1 and k2 can be derived before the steady state has been reached), and may provide insight into the potential of NMs to be taken up by organisms. However, dynamically derived accumulation factors can vary quite a lot between studies, although this variation is not necessarily related to exposure concentrations.6
Based on this discussion and underlying references it can be concluded that accumulation factors should be avoided when based on ratios between organisms and media, due to the fact that equilibrium partitioning theory does not apply to chemicals that are not thermodynamically stable, such as nanomaterials. Furthermore, they should be used with care when derived dynamically in time resolved experiments, because ADME processes related to the fate of nanomaterials in organisms may not always be resolved completely. Nanomaterials can also occur in different forms which may interact and as such affect each other's accumulation patterns. This may also hamper the application of simple accumulation or concentrations factors.
Physiologically based pharmacokinetic (PBPK) or biodynamic models
Accumulation of chemicals in organisms, including nanomaterials, depends on their availability in the exposure medium and on the physiological traits of the species involved, driving accumulation, distribution, metabolism and excretion (ADME). Physiologically based pharmacokinetic (PBPK) or biodynamic models can be used to model these processes dynamically.72,73 Different approaches can be applied, depending on routes of uptake and the ADME processes that need to be included (Fig. 2).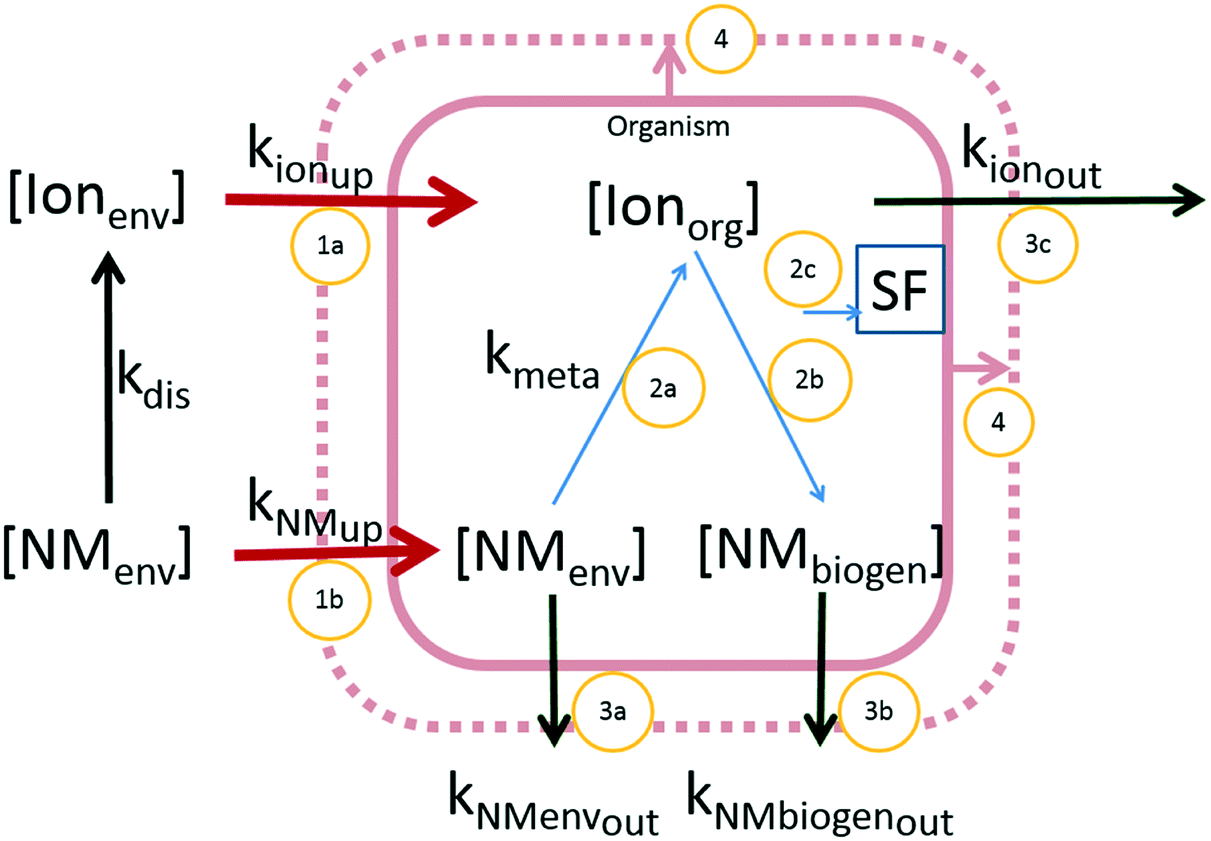 | ||
| Fig. 2 Conceptual overview of different forms of metal-based NMs and kinetic pathways of uptake and elimination. Forms of materials: NMenv: nanomaterial in the original form (can be both in the environment and in the organism), Ionenv: ionic form of the NM-material in the environmental compartment (dissolved), Ionorg: ionic form in the organism (may originate from both Ionenv as well as dissolved from NMenv in the organism), NMbiogen: biogenic particulate form, Fi: inert fraction; kinetic processes displayed by arrows. Red arrows: uptake. 1a: Ionic uptake, 1b: particulate uptake; blue arrows: within organism kinetics. 2a: dissolution of NMs, 2b: biogenic formation of particulate materials, 2c: transport to the inert fraction; dark green arrows: elimination: 3a elimination of original NM, 3b: elimination of the biogenic particulate material, 3c: elimination of the ionic form. Pink arrows: growth dilution (4). | ||
The simplest approach considers the organism as a single compartment, with just uptake and elimination as kinetic processes (only include processes 1a and 1b combined for uptake; 3a, 3b and 3c combined for elimination, Fig. 2). Uptake under these assumptions is described by eqn (1). When the test organisms are transferred to a clean medium, they will eliminate the NMs. The process of elimination can be described by eqn (2):
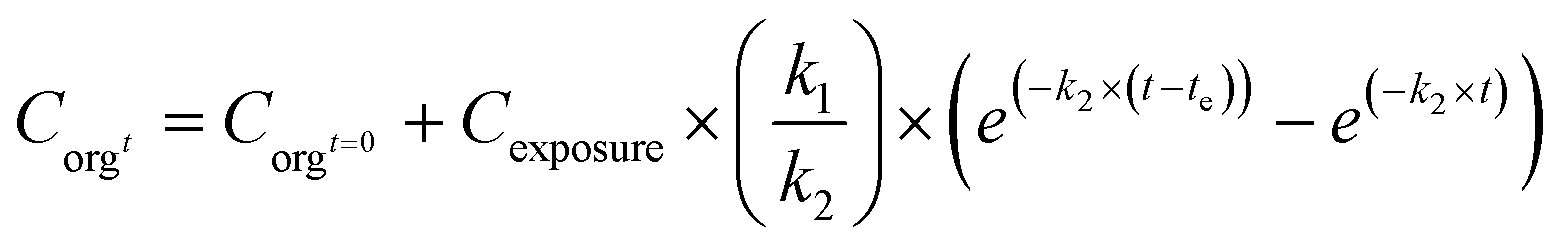 | (2) |
C orgt: concentration in the organism at time t (mg kg−1); Corgt=0: concentration in the organism at t = 0 (mg kg−1); Cexposure: concentration in the exposure medium (mg kg−1 or mg L−1); k1: uptake rate constant (kgmedium kgorganism−1 per day or Lmedium kgorganism−1 per day), k2: elimination rate constant (day−1); t: time (day); te is the time at which the test organisms are transferred from a contaminated to a clean medium (day).
In the case of metals, such a simple one-compartment model may not be fully adequate, as storage in a stored fraction may occur in specialized tissues of organisms (process 2C, Fig. 2)73 as was shown in earthworms in which metals were sequestered in stored forms that were not biologically active.74 Such storage may also be applicable to nanomaterials or the metal ions released from NMs, which was illustrated in studies on the uptake of Ag-NMs in D. magna65 and in isopods.9 In such cases, the model may be extended with a stored fraction (SF no dimension). In other studies an inert fraction has been used in order to account for the fraction stored in organisms, however, that approach only accounted for storage in the elimination phase of their experiments but was not included in the first experimental phase when organisms were exposed.9,65,75 To overcome this, the extended model with a SF can be defined using eqn (3a) and (3b):
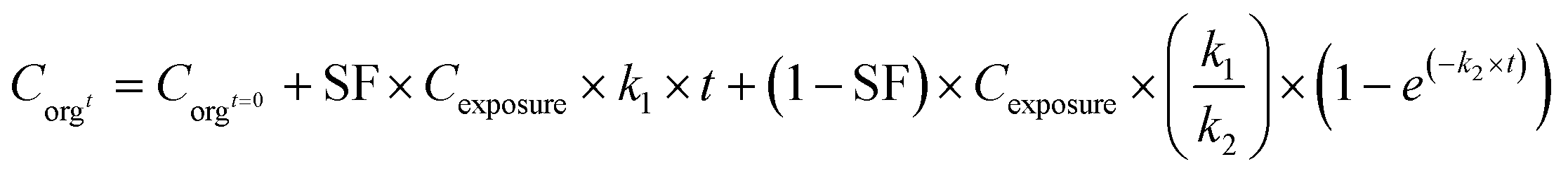 | (3a) |
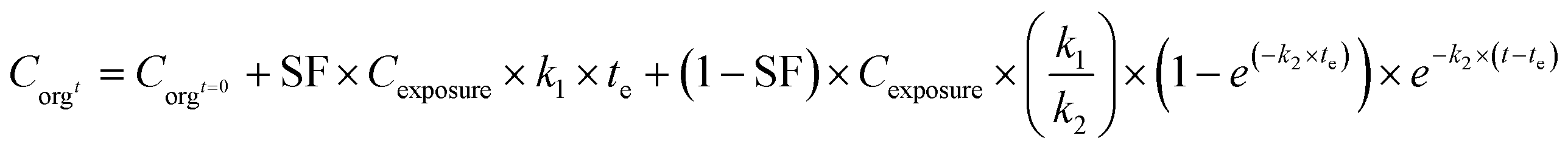 | (3b) |
When applying the modelling with a SF to the work of Ribeiro et al. 2017 (ref. 65) using the total Ag-concentrations in Daphnia magna after exposure to Ag-NMs14 based on eqn (3a) and (3b), and comparing this with the outcomes using the models without a SF (eqn (1) and (2)), both approaches were highly significant (P < 0.001), but the percentage explained variance increased from 61 to 71% of the total variance with the inclusion of the SF. A SF of 0.099 ± 0.033 was derived (average ± standard error). The k1 value increased from 0.190 ± 0.025 to 0.363 ± 0.121, while the k2 value increased from 0.031 ± 0.007 to 0.124 ± 0.059. The SF of 0.099 indicates that 9.9% of all Ag that was internalised by the daphnias during exposure (equals k1 × ConcAg_medium × day) is in the stored fraction. This implies that the percentage stored in the organism is actually higher than, in this case, 9.9% of the accumulated Ag (internalised Ag minus excreted Ag), while this percentage increases over time. This example illustrates that inclusion of potentially relevant storage may affect the quantification of the other rate constants considerably which implies that selection of ADME processes in the modelling is extremely important, and even more that comparison of kinetic parameters between studies can only be done with the total model in mind, including all other kinetic rate constants. This approach to model the stored fraction assumes that there is no limit to the storage capacity of the organisms, be it in e.g. metallothioneins for metal-NMs or fat deposits for organic NMs, and secondly that the stored fraction is not eliminated over time. When including such an stored fraction (SF), the ratio between k1 and k2 no longer yields a correct estimate of accumulation factors (BCF/BSAF). The assumption that the stored fraction is not eliminated over time may be difficult to meet from a physiological point of view. An alternative to meet this is to assume the organism to be composed of two compartments, one that does eliminate the compound fast and one that accumulates the compound but eliminates it (very) slowly. By doing so, the model describing the uptake rate of the compound separately describes uptake in the two different compartments and includes a rate constant for the transfer of the compound from the first to the second compartment. The models describing the uptake phase are shown in eqn (4a) and (4b) (taken from ref. 73).
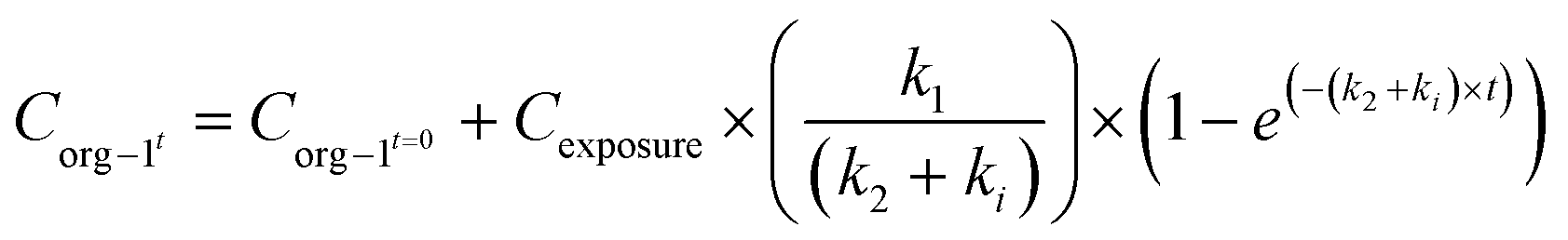 | (4a) |
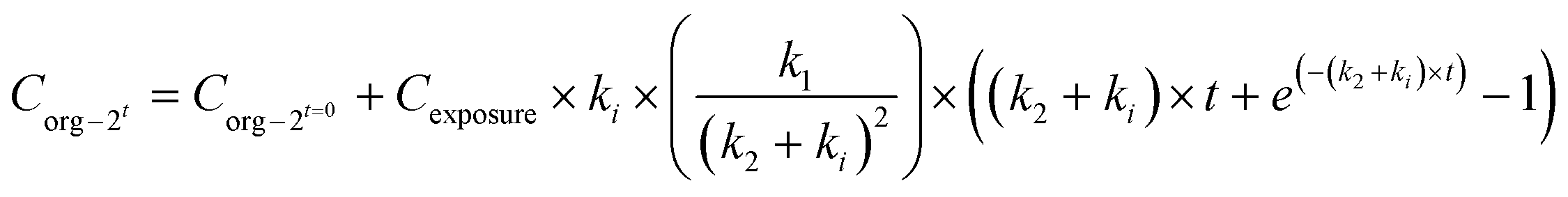 | (4b) |
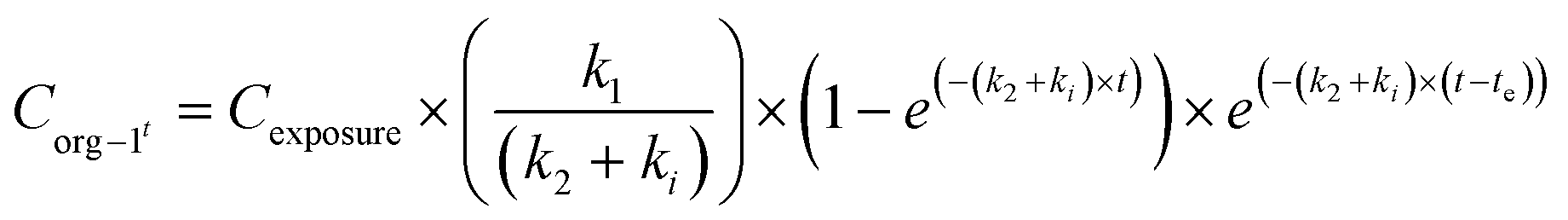 | (5a) |
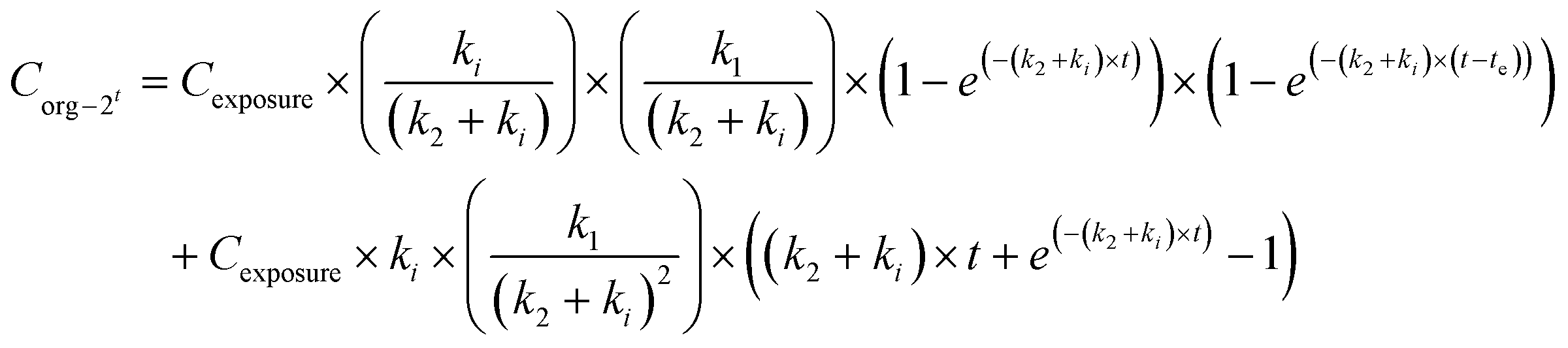 | (5b) |
Quantification of the rate constants k1, k2, and ki for a two compartment model requires measurements of the NM in the storage tissue, which for invertebrates may be difficult and sometimes even impossible because the material may be stored throughout the body. In such a case, application of the model with the stored fraction may be more feasible.
Depending on the complexity of the toxicokinetics of the nanomaterials in the organisms, different model formulations can be integrated in the models, including different excretion pathways with different elimination rate constants, growth dilutions, changes over time of the bioavailable fraction of the chemical in the exposure medium or inclusion of storage in a stored fraction using two compartment models.73 Several studies have used PBPK or biodynamic models to quantify the bioaccumulation of different nanomaterials, with model formulations.5,9,21,22,76 Studies on metal-based nanomaterials are generally based on the total metal content and do not take the different forms of the materials into account, although recent studies have focussed on this.14 A modelling example on the uptake of different forms of Ag in earthworms illustrated the importance of form-specific approaches, and uptake levels and forms of Ag (as particulate or ionic uptake) were found to depend on i.e. the rate of dissolution of the silver nanoparticles and ADME processes.17 These processes thus need to be included in the modelling, using available model formulations and equations.73
Due to transformation and ageing of nanomaterials, stable exposure concentrations as assumed by the simple one-compartment model will hardly be the case. This is best illustrated by a metal-based NM, like Ag NMs. Such NMs are prone to release Ag+ ions into the environment, so organisms exposed to a medium spiked with Ag NMs in fact will be exposed to a mixture of NMs and free Ag ions (although Ag+ may also complex with Cl− or S2−), which might even change in composition over time. In the most simple case of a mixture of Ag NMs and Ag+ ions, the Ag uptake rate in the test organisms might be modelled with a model that includes two uptake rate constants to account for the contribution of both Ag forms. The model could take the form
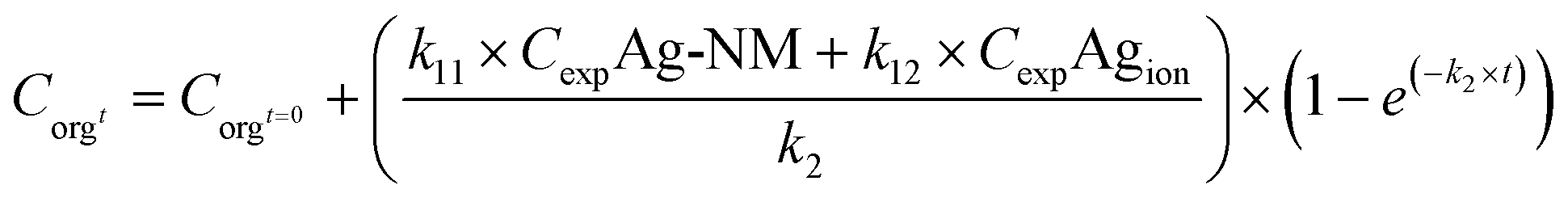 | (6) |
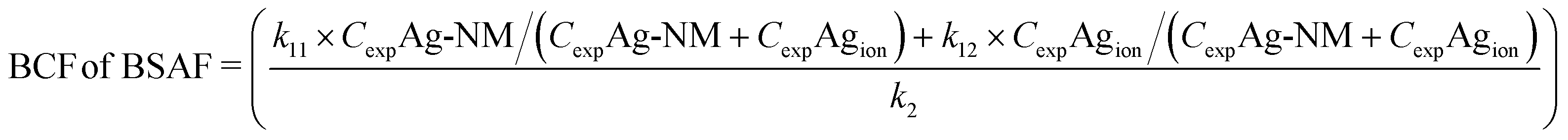 | (7) |
Excretion rates may also be dependent on the form in which the material is present in the organism. In such a case, a different k2 value may be applied, a fast and a slow elimination rate constant (k2fast, k2slow). The fast elimination stops after the fast eliminating pool is depleted (tendk2fast). The uptake can be modelled according to eqn (8):
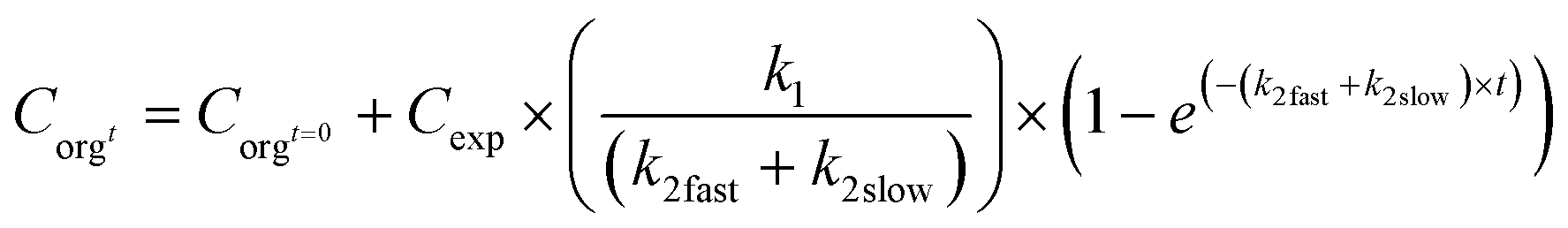 | (8) |
In combination with eqn (6), different uptake rates can also be included for the different forms. Elimination in the phase that fast elimination still takes place (t < tendk2fast) can be modelled as in eqn (9a), and after this, according to eqn (9b). When fitting the model to the data, tendk2fast can be assessed iteratively together with k1, k2fast and k2slow.
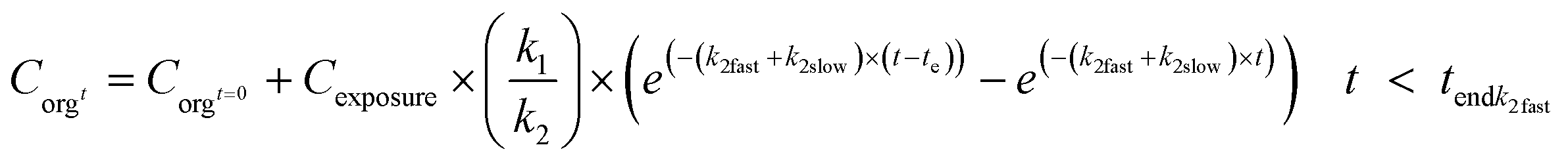 | (9a) |
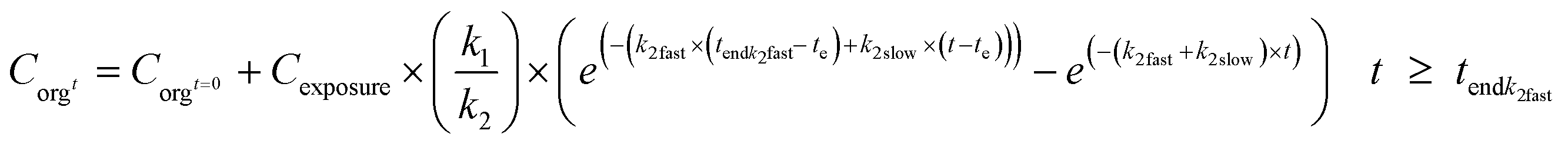 | (9b) |
In addition to the above described models, other processes may need to be covered. It is possible to estimate uptake and elimination rate constants accounting for a steady decline of exposure concentrations. This is done by adding kdeg (day−1) to eqn (1) to yield eqn (10).73
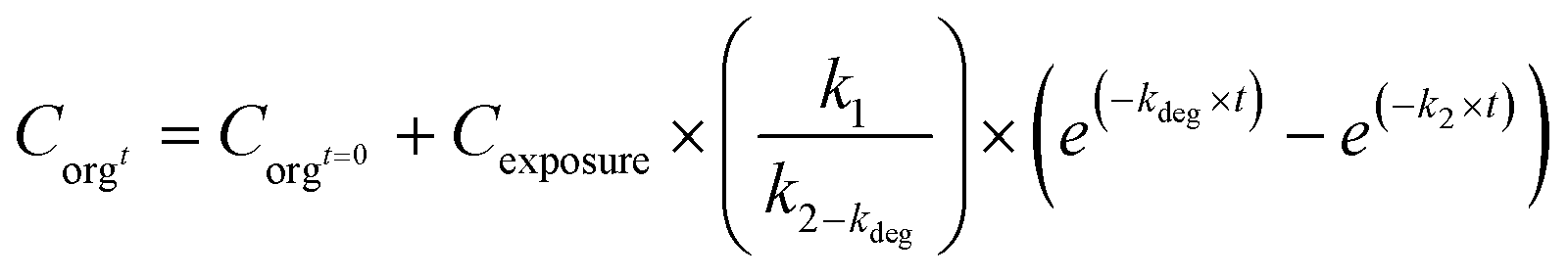 | (10) |
Such models may also be rewritten to account for a steady change in exposure concentration, as may be the case when only the free ions released from a metal-based NM would be taken up. The rate at which the metal ions are released may be included in the equations for uptake and elimination kinetics. For carbon fullerenes, such an approach was followed to model the uptake kinetics with inclusion of declining exposure concentrations due to settling of the fullerenes during the experiment.48 This was done by including the linear regression slope of the natural logarithm of the fullerene concentration in the aqueous phase versus exposure time in the model formulations.
Another factor that may need to be accounted for is the biogenic formation of NMs inside the exposed organisms, a process which has been suggested for Ag to occur in earthworms as part of the detoxification of metals in insoluble granules.14 In such a case, the model may take the form of a two-compartment model (eqn (4a) and (4b)) and ki could be considered the rate at which NMs are formed from ionic metals taken up by the organism.
A final case to mention is when exposed organisms show a considerable change in biomass during exposure to the nanomaterials. A significant increase of biomass may affect the uptake kinetics as it may lead to so-called growth dilution, while in the case of considerable mass loss the compound may become more concentrated in the biological tissues (process 4 in Fig. 2). To account for these situations, a growth rate (kg) has to be calculated, which then can be included in the equations for uptake and elimination by simply replacing k2 in eqn (1) and (2) with k2 + kg.73
In the following cases the applicability of PBPK or biodynamic models will be illustrated, based on existing datasets and references.
Case study i: Ag uptake from enchytraeids exposed to Ag2S NMs with and without correction for mass loss.
In this case, the effect of biomass changes in the organisms on the prediction of rate constants will be illustrated. Adult age-synchronized Enchytraeus crypticus was exposed for 14 days to Lufa 2.2 soil spiked with 20 nm Ag2S NMs at 2.5 mg Ag per kg dry soil (for details, see ESI†). At different time intervals, animals were sampled for the determination of Ag uptake kinetics. After 14 days, the remaining animals were transferred to clean soil to assess Ag elimination for 14 days. Three replicate samples were taken at each sampling time, and animals were allowed to void their guts after, freeze dried, weighed, acid digested and analysed for Ag. Fig. 3 (left) shows the Ag concentrations in the animals and the fit of the one-compartment model (eqn (1) and (2)) to the data. This resulted in k1 = 0.057 gsoil ganimal−1 per day and k2 = 0.370 day−1. Upon analysis, it turned out that the animals gradually lost weight during the experiment. Fig. 3 (right) shows the mass over time of the animals, from which a negative kgrowth value of −0.032 day−1 was derived. Correcting the uptake and elimination kinetics for weight change was done by replacing k2 in eqn (1) and (2) with ‘k2 + kgrowth’. This did not affect the fit of the curve to the data and did not affect k1, but the k2 value increased from 0.370 to 0.402 day−1, so did the value of kgrowth. This case clearly indicates the potential to over- or underestimate kinetic rate constants when not including all relevant processes that may drive the internal concentrations of NMs.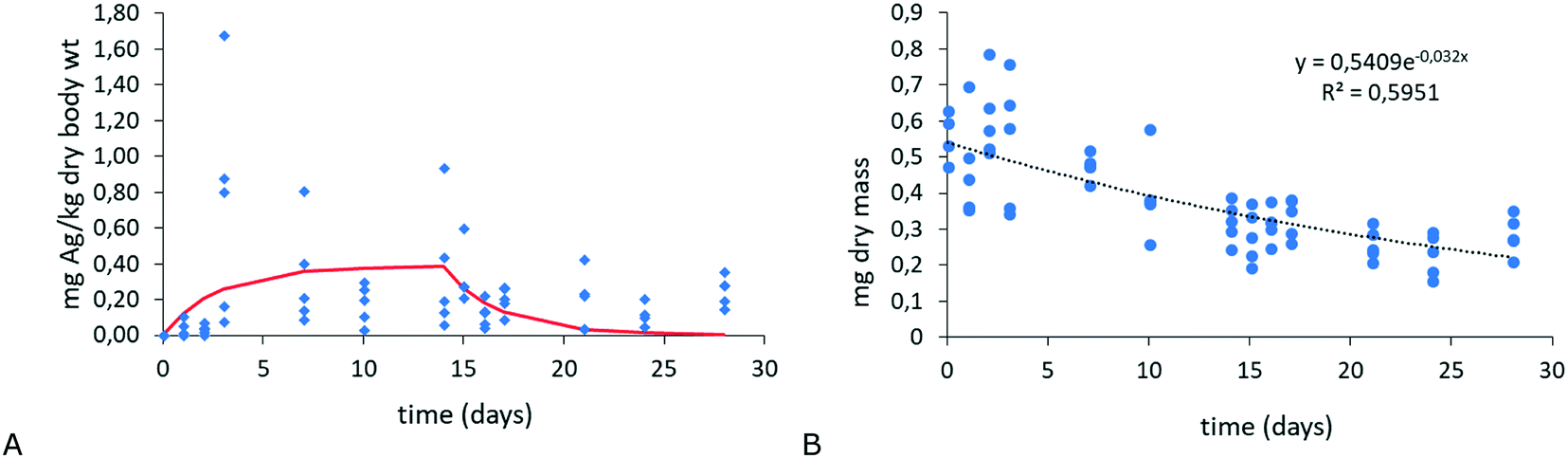 | ||
| Fig. 3 A: Ag uptake and elimination in E. crypticus exposed to Ag2S-NMs (20 nm, 2.5 mg Ag per kg dry soil) (x-axis: days; y-axis: mg kg−1). B: Mass over time of animals during the uptake and elimination kinetics test (x-axis: days; y-axis: mg dry mass). | ||
Case study ii: modelling approaches with different PBPK model definitions
For the different model formulations as described in eqn (1) to (9b), specific kinetic rate constant parameters are needed. To assess potential effects of different model definitions and kinetic parameters on the analysis and interpretation of experimental results, data on the uptake of different forms of Ag in earthworms14 were analysed with different model formulations, including scenarios with either separate uptake rates for different forms of NMs (e.g. ionic and particulate uptake of Ag) or two elimination rates (fast and slow elimination). In Baccaro et al. (2018), different exposure experiments were conducted in which earthworms were exposed to AgNO3, pristine Ag-NMs (which showed dissolution) and Ag2S-NMs.14 The experimental data shown in Fig. 4, and used in this example, were taken from the pristine Ag-NM exposure experiment. When a single compartment was fitted with a single k1 and k2 (eqn (1) and (2)) the model described the data with a significant fit, according to Baccaro et al. (2018)14 (Fig. 4: ‘Predict’ p < 0.01; r2: 0.75; for k1 and k2 see Table 1). The fit (as r2) improved when an additional excretion rate was included in the model (eqn (8), (9a) and (9b)), although the significance of the regression decreased due to the lower degrees of freedom (Fig. 4 ‘Predict separate k2’; p < 0.05; r2: 0.85; for k1, k2slow and k2fast see Table 1). The uptake rate constant was slightly greater when a second elimination rate constant was included, while the fast elimination rate constant was slightly greater than the original one and the slow elimination rate constant was smaller. Directly after the transfer of the worms to clean soil, the Ag-concentrations decreased rather fast (driven by both fast and slow elimination routes77), however, after a longer time the elimination rate decreased. The modelled fast elimination only occurred up to 7 days after placing the worms in clean soil, after which only the slow elimination remained (Endk2fast = 7 days, Table 1).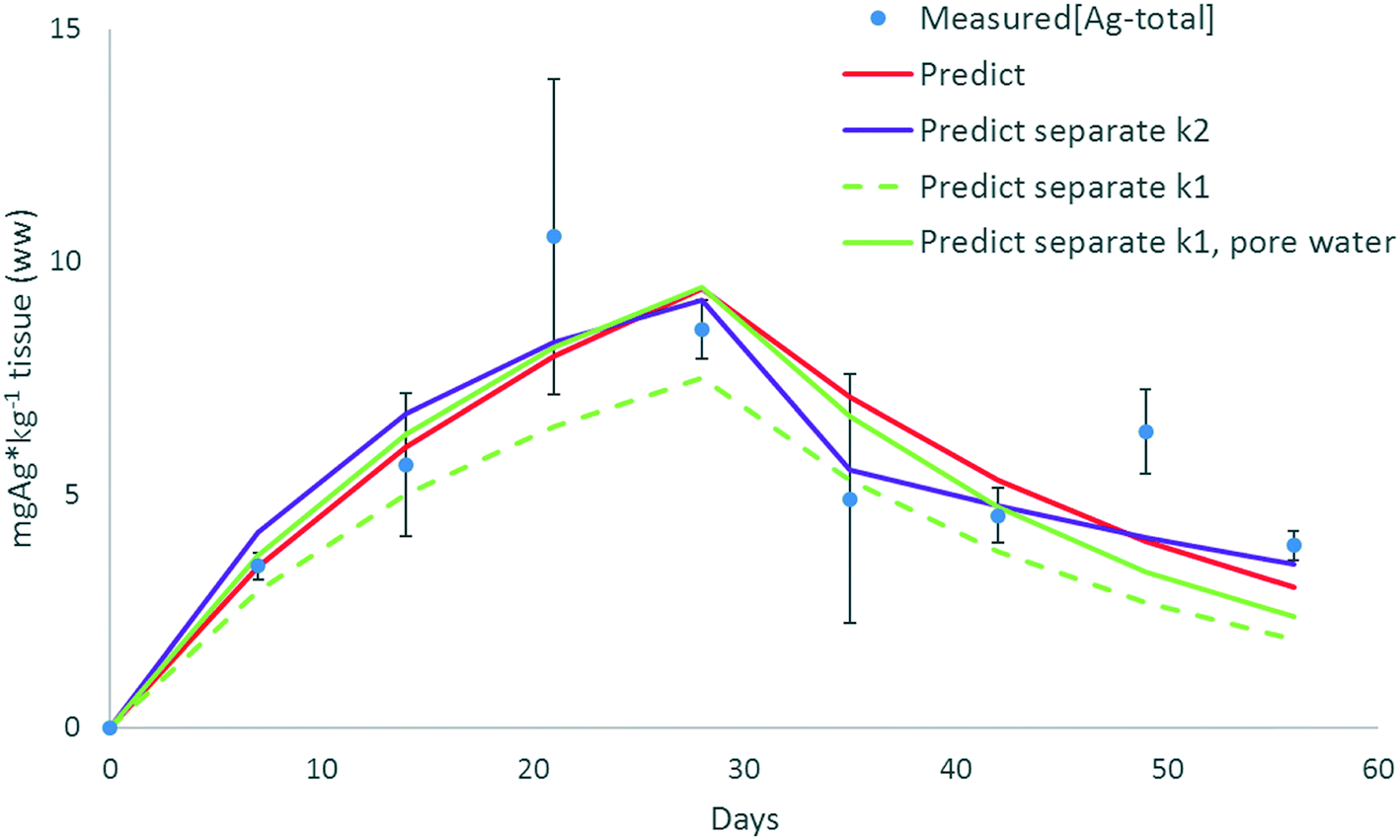 | ||
| Fig. 4 Modelling of [Agtotal] in earthworms exposed to pristine Ag-NMs (20 nm, 10 mg kg−1); measured[Ag-total]: empirical data Baccaro et al. (2018)14 (average and stdev); for equations and parameters for the different scenarios see Table 1. Day 0–28: earthworms in spiked soil, days 28–56: earthworms transferred to clean soil. | ||
| Model | k 1 | k 1ion | k 1NM | k 2 | k 2fast | k 2slow | k dis | Endk2fast | Equations | Significance (p-value) | R 2 | Parameters |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Scenario, Fig. 4 | kg kg−1 per day | kg kg−1 per day | kg kg−1 per day | Day−1 | Day−1 | Day−1 | Day−1 | Days | (Uptake phase, elimination phase) | |||
| Predict | 0.063 | — | — | 0.041 | — | — | — | — | (1) and (2) | <0.001 | 0.75 | k 1, k2 fitted to data |
| Predict separate k1 | — | 0.055 | 0.008 (ref. 14) | 0.054 | — | — | 0.82 | — | (6) and (2) | <0.001 | 0.74 | k 1ion,14k1NM;14kdis (ref. 17) |
| Predict separate k1, pore water | 16.8 (L kg−1 per day) | 0.008 | 0.044 | — | — | — | — | (6) and (2) | <0.001 | 0.74 | k 1ion,14k1NM,14 recalculated pore water concentrations14 | |
| Predict separate k2 | 0.085 | — | — | — | 0.055 | 0.018 | — | 7 | (8), (9a) and (9b) | <0.001 | 0.83 | k 1, k2fast, k2slow, Endk1 fitted to data |
These models were fitted to the uptake data of Ag from the pristine particle experiment. The accumulation patterns in this experiment in which the earthworms were exposed to two forms of Ag (Ag-NMs and dissolved Ag+), however, may also be predicted by using parameters from the other two experiments specifically on Ag+ and Ag-NM exposure, in Baccaro et al. (2018),14 by using a model with two uptake rate constants derived from those two experiments (eqn (5a), (5b), (6a) and (6b)). The uptake rate constant for the ionic uptake (k1ion) could be derived from the experiment with Ag-NO3,14 while k1NM was based on the experiment with Ag2S-NMs in that study (for parameters see Table 1). When applying k1ion, k1NM, an estimate of the dissolution rate of the pristine Ag-NMs had to be made, the source of ionic Ag in the soil, using eqn (11).
| CNMt = CNMt=0 × exp(−kdis×t) | (11) |
k dis was set at 0.82 day−1.78 The modelling with these parameters resulted in somewhat lower concentrations in the earthworms (Fig. 4, ‘Predict separate k1’, p = 0.15; r2: 0.74; Table 1). The relatively high r2 value of 0.74 indicates that the modelled uptake and elimination patterns did follow the patterns of the empirical data, which would indicate that the ionic uptake was mainly driving the overall uptake in both original experiments on Ag-salts and pristine Ag-NMs in Baccaro et al. (2018).14 Soil pore water concentrations could be indicative of the bioavailable fraction of the ionic Ag in the experiments, which were comparable between the original experiments (40.5 μg L−1 for Ag-NM exposure versus 37.9 μg L−1 for the Ag–NO3 exposure). When using the pore water concentrations as proxy for exposure levels for deriving k1ion (37.9 μg L−1), it results in a high k1ion value of 16.8 (Table 1). When applying this k1ion to predict Agtotal in the earthworms based on pore water concentrations in the pristine Ag-NM experiment (40.5 μg L−1), the resulting modelling fitted the data significantly (Fig. 4, ‘Predict separate k1, pore water’ p < 0.05; r2: 0.74). The fitted curve is close to the single k1 modelling exercise.
The modelling approaches in this case, using the different uptake rate constants, indicate the possibility of modeling the uptake of different forms of metal-based NMs and released ionic forms by organisms. In dynamic settings, rate constants derived in single-form experiments (i.e. ionic exposure or exposure to non-soluble NMs) may be used to predict accumulation patterns in experiments in which different forms may be included (in the case of soluble NMs). However, for such approaches, the fate of the NMs in the soil needs to be included in the modelling as well as their availability. This will be illustrated in greater depth in the following case.
Case study iii: modelling uptake of Ag by earthworms, including dissolution of Ag-NMs and adsorption/desorption onto the soil
Uptake experiments with NMs are mostly performed based on total concentrations, not taking into account the form of the material that is present. In the case of carbon based NMs the form is likely either fullerenes, carbon nanotubes or graphene, unless the NM is degraded by exogenous enzymes from the organisms. For metal-based NMs, both particulate and ionic metal forms may be taken up by the organisms, as was illustrated in Case study ii. Hence, when using NM concentrations to predict the uptake, the dissolution of the particles and the adsorption/desorption of ions onto soil particles needs to be taken into account. This can be done with the conceptual model, as depicted in Fig. 5. Based on this model, the uptake of Ag from different sources (ionic, dissolving NMs; stable NMs) by earthworms was modelled in three experimental scenarios using data from Baccaro et al. (2018):14 i) exposure of earthworms to AgNO3 ([Ag]tot: 9.3 mg kg−1, assuming all dissolved at the start); ii) exposure of earthworms to pristine Ag-NMs ([Ag]tot: 9.0 mg kg−1); iii) exposure of earthworms to (non-dissolving) Ag2S-NMs ([Ag]tot: 3.7 mg kg−1), all measured as total [Ag] in the samples. Based on this experimental data, the hypothesis that the Ag-uptake of earthworms is mainly via the ionic form can be explored. | ||
| Fig. 5 Conceptual model used in the current case, depicting the modelled fate of Ag-NMs in soil, and the potential accumulation of ionic Ag in earthworms. For the dissolution process, no equilibrium is assumed. | ||
Model description: In the model, Ag-NM concentrations decrease over time, due to dissolution, according to eqn (11). kdis differs among the types of NMs. For pristine Ag-NMs this was 0.82 day−1,78 while for sulphidised Ag-NMs (Ag2S-NMs), a much lower dissolution rate of 4.6 × 10−4 day−1 was derived from ref. 79 based on data for particles with a S/Ag ratio of 0.0192 (see ESI†). The S/Ag ratio of the sulphidised Ag-NMs used in Baccaro et al. (2018)14 was around 0.5 at fully sulphidised sites. However, the NMs were not fully sulphidised with sites with much lower S/Ag ratios, as low as 0.015 (unpublished data), so close to 0.0192 from ref. 79. Dissolved Ag may adsorb onto soil particles, diminishing its bioavailability since it is assumed that the biouptake of Ag ions is only from free Ag+ ions in the pore waters. The adsorption kinetics are described by the adsorption and desorption rate constants (kads, kdes (day−1)) which were derived based on eqn (12) and (13):
| kads = kd × kdes | (12) |
| kd = kf × ratiowatertosoil | (13) |
Using the models on dissolution and adsorption/desorption, and the inputs from ref. 14, the soil concentrations of the different forms (particulate, dissolved Ag adsorbed to soil particles and non-adsorbed ionic Ag) were predicted (Fig. 6). Concentrations in the earthworms could be fitted to the modelled variable dissolved Ag-concentrations in the soil, quantifying k1 and k2 based on (modelled) dissolved Ag-concentrations and not on total soil concentrations as was done by Baccaro et al. (2018).14 The expectation was that the variation of the k1 and k2 would be relatively small among the different scenarios, because the most important form of Ag driving the uptake (dissolved ionic Ag) was used as a base for the modelling.
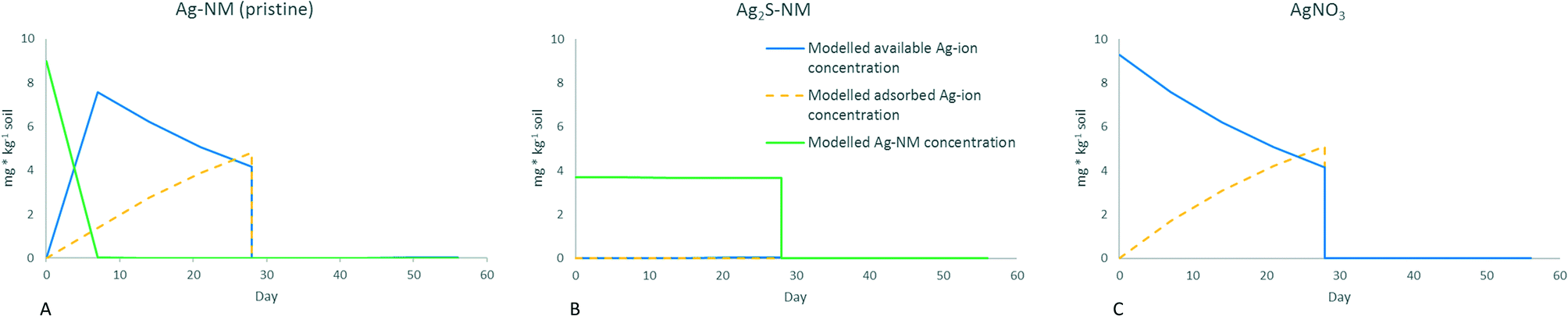 | ||
| Fig. 6 Modelled concentrations of different forms of Ag in the soil. A: Exposure to Ag-NM pristine, B: exposure to Ag2S-NM, C: exposure to AgNO3. For all graphs: red line: available desorbed Ag-ions, orange dotted line: Ag-ions adsorbed to soil particles, not available, green line: modelled Ag-NM concentrations. All concentrations in mg kg−1, initial concentrations used as inputs for modelling were taken from Baccaro et al. 2018.14 Day 0–28: earthworms in spiked soil, days 28–56: earthworms transferred to clean soil. | ||
Fig. 6 shows the distribution of the different forms of Ag in the soil while Fig. 7 shows the total Ag concentrations in the earthworms (modelled and measured) for the three scenarios. In the ionic exposure scenario, the initial dissolved Ag-ion concentration was 9.3 mg kg−1 which decreased over time due to adsorption to the soil. For the pristine Ag-NMs, a dissolution rate of 0.82 day−1 resulted in over 97% dissolution within three days, so although the dissolved Ag ionic concentration in this scenario started at 0 mg kg−1, it rapidly followed the same kinetics as the ionic exposure. For the Ag2S-NM exposure, the concentrations of dissolved Ag ions were orders of magnitude lower those for the other two scenarios, due to the low rate of dissolution. When the worm concentrations were fitted to the variable desorbed Ag-concentrations (Fig. 7), significant k1 and k2 parameters could be derived for the different forms of Ag (Table 2).
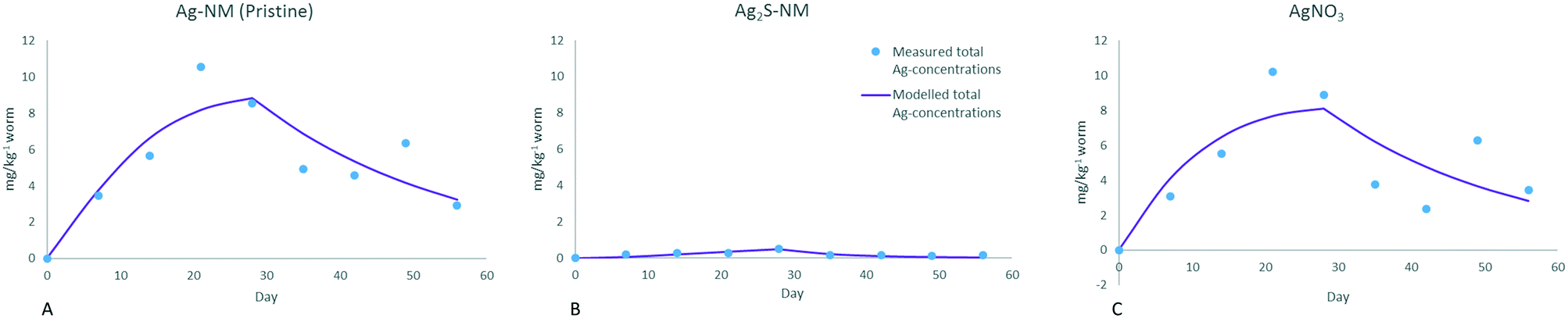 | ||
| Fig. 7 Modelled total Ag-concentrations in earthworms (mg kg−1 ww), based on different exposure scenarios and soil concentrations as shown in Fig. 6. Day 0–28: earthworms in spiked soil, days 28–56: earthworms transferred to clean soil. | ||
| Ag source | k 1 (kg kg−1 per day) | k 2 (day−1) |
|---|---|---|
| Ag-NM | 0.0866 | 0.0355 |
| Ag2S-NM | 2.309 | 0.113 |
| AgNO3 | 0.0787 | 0.0383 |
Uptake and elimination rate constants for the dissolving pristine Ag-NMs are similar to the ones for the ionic exposure, indicating similar uptake kinetics. This was also concluded by Baccaro et al. (2018).14 The uptake rate constants for Ag-NM and AgNO3 exposure only differed by approximately 10% within each study. These uptake rate constants are slightly higher in the current study when compared to Baccaro et al. (2018)14 (0.087 versus 0.061 and 0.079 versus 0.055 (kg kg−1 per day) for Ag-NMs and AgNO3, respectively). This is expected, since the uptake rate constants of the current study are based on available ionic Ag concentrations, while Baccaro et al. (2018)14 used (higher) total concentrations as inputs for the modelling. Ratios between the rate constants of the two studies are similar among the forms (1.42 and 1.43 respectively), indicating similar impacts of the inclusion of the availability of the Ag-ions for uptake for both forms. The overall modelled uptake of Ag from exposure to Ag2S-NMs is low, yet the uptake rate (k1) is larger in comparison with the other forms (2.3 kg kg−1 per day), which is also higher than was found by Baccaro et al. (2018)14 (0.008 kg kg−1 per day). The high rate constant of the current study is due to the extremely low concentrations of available Ag, indicating that sole uptake via available ionic Ag may not fully explain the uptake in the case of these non-soluble Ag2S-NMs. This would imply that in the case of such insoluble NMs, other routes of uptake (e.g. particulate) may also play a significant role, although the absolute accumulation of particulate Ag is much lower than that of the ionic form (the measured concentrations in the Ag2S exposed worms are significantly lower than in worms exposed to the other forms). The relatively large k1 value for the uptake of Ag from Ag2S-NMs in the current study may also be due to underestimation of the dissolution of the Ag2S-NMs used, although this is less likely considering the extent of the differences. Excretion rates (k2) are similar between the current study and Baccaro et al. (2018)14 for all forms (0.036 versus 0.04, 0.113 versus 0.064, and 0.038 versus 0.044 for Ag from Ag-NM, Ag2S and AgNO3, respectively) which is expected since k2 is mainly dependent on the internal worm concentrations. The higher excretion rate for Ag from Ag2S-NMs could suggest that a different form of Ag is excreted faster than the form in the ionic and pristine Ag-NM exposed earthworms, although this could not be confirmed in Baccaro et al. (2018).14
The results of this case study indicate that, in concurrence with ionic metal exposures, bioavailability is important to consider in the case of the accumulation assessment of dissolving metal-based NMs. Furthermore, uptake of particulate NMs seems to be relevant in the case of non-dissolving NMs, although the absolute accumulation of non-dissolving NMs may be significantly lower than that of dissolving NMs.
Recommendations and guidance
The assessment of bioaccumulation potential is an important facet of the environmental risk assessment of NMs, and currently regulatory bodies include the use of invertebrates for bioaccumulation testing. For example, the OECD technical guidance (TG) 317 on earthworm bioaccumulation testing indicates that uptake curves can be drawn and bioaccumulation factors derived. However, such technical guidance documents may need amending for NMs. Based on the available literature, neither BLMs nor bioaccumulation factors based on measured data are to be recommended for modelling of (longer term) bioaccumulation of different forms of nanomaterials. Assumptions underlying these modelling approaches, including equilibrium theory that relates to uptake of solutes, are not met in the case of NMs. Dynamic PBPK-modelling approaches are more suitable for nanomaterials. Different uptake and elimination processes can be included in the modelling frameworks, including fast (active) uptake and elimination but also slow (passive) processes. Storage as inert fractions, biogenic transformation of NMs, growth dilution and other internal kinetic processes can also be incorporated into the modelling formulations, although the determination of specific rate constant parameters may be a challenge. Accumulation experiments to assess kinetic parameters should include uptake and elimination phases and the analyses should in principle only include materials that cross epithelia, unless the focus is on biomagnification in food chains.In order to allow read across of data and parameters between studies, it is essential to quantify concentrations of the different forms of nanomaterials. Modelling of available metal ion concentrations in the pore water has been illustrated to be a useful proxy for bioavailability in experiments on soil organisms in the case of dissolving NMs. For particulate forms of NMs the main driver of uptake is phagocytosis, however, little is known on the drivers of NM bioavailability. Uptake of particulate forms is generally much lower than the ionic uptake but may still be relevant in modelling the total uptake of NM, especially for slowly, or non-dissolving NMs. The quantification of the availability of the different forms demands accurate protocols e.g. for pore-water extraction, with methodologies able to separate particles from ions to detail NM speciation. Filling this data gap is essential in order to be able to model the accumulation of different forms of specific NMs in an integrated way, coupling the environmental fate of NMs with uptake in biota. In addition, biogenic formation of particulate materials in tissues is a well-known protection mechanism by which organisms can store metals in a less toxic form. When addressing uptake of metallic NMs, it should always be confirmed that particles found in the organisms are similar to the ones they were exposed to. For this, the size, shape and elemental composition of NMs need to be established. Without this information, particulate uptake may be overestimated since biogenic particles are included in the derivation of the parameters.
More complex PBPK models demand more parameters that need to be quantified. In complex cases, exposure experiments should be designed with ample statistical power. It was shown in the examples that although the r2 value of a modelling approach increased (i.e. scenario ‘Predict separate k2’ versus ‘Predict’), its significance decreased due to decreased degrees of freedom (Table 2). Furthermore, only processes should be included for which kinetic rate constants can be quantified. For instance, the biogenic formation of particulate metal in organisms may depend on the internal metal concentration, hence, this may only be induced at higher concentrations. In this case, the kinetic rate constant of the biogenic formation is dependent on the internal concentration, which may hamper the modelling of form specific accumulation of NMs. In such cases, it may be needed to restrict the modelling to total concentrations of the material.
Author contributions
All the authors have contributed to the discussions leading to the manuscript and the writing of the manuscript.Conflicts of interest
The authors declare no conflicts of interest.Acknowledgements
All the authors were funded by the EU H2020 project NanoFASE (Nanomaterial Fate and Speciation in the Environment; grant no. 646002). DD and AJK received additional funds from the Slovenian Research Agency (research program, Integrative Zoology and Speleobiology, P1-0184). SL, PVS and ZK received additional financial support from FCT/MCTES, through national funds, to CESAM (UID/AMB/50017/2019); PVS was awarded with a PhD grant by FCT (SFRH/BD/51571/2014).References
- S. J. Klaine, A. A. Koelmans, N. Horne, S. Carley, R. D. Handy, L. Kapustka, B. Nowack and F. v. d. Kammer, Paradigms to assess the environmental impact of manufactured nanomaterials, Environ. Toxicol. Chem., 2012, 31, 3–14 CrossRef CAS PubMed.
- K. Hund-Rinke, A. Baun, D. Cupi, T. F. Fernandes, R. Handy, J. H. Kinross, J. M. Navas, W. Peijnenburg, K. Schlich, B. J. Shaw and J. J. Scott-Fordsmand, Regulatory ecotoxicity testing of nanomaterials – proposed modifications of OECD test guidelines based on laboratory experience with silver and titanium dioxide nanoparticles, Nanotoxicology, 2016, 10, 1442–1447 CrossRef CAS PubMed.
- R. D. Handy, N. W. van den Brink, M. Chappell, M. Mühling, R. Behra, M. Dušinská, P. Simpson, J. Ahtiainen, A. N. Jha, J. Seiter, A. Bednar, A. Kennedy, T. F. Fernandes and M. Riediker, Practical considerations for conducting ecotoxicity test methods with manufactured nanomaterials: what have we learnt so far?, Ecotoxicology, 2012, 21, 933–972 CrossRef CAS PubMed.
- E. Lahive, K. Jurkschat, B. J. Shaw, R. D. Handy, D. J. Spurgeon and C. Svendsen, Toxicity of cerium oxide nanoparticles to the earthworm Eisenia fetida: subtle effects, Environ. Chem., 2014, 11, 268–278 CrossRef CAS.
- Z. M. Świątek, C. A. M. van Gestel and A. J. Bednarska, Toxicokinetics of zinc-oxide nanoparticles and zinc ions in the earthworm Eisenia andrei, Ecotoxicol. Environ. Saf., 2017, 143, 151–158 CrossRef PubMed.
- C. W. Isaacson, L. Sigg, A. A. Ammann, J. Stadnicka-Michalak and K. Schirmer, Interactions of TiO2 nanoparticles and the freshwater nematode Plectus aquatilis: particle properties, kinetic parameters and bioconcentration factors, Environ. Sci.: Nano, 2017, 4, 712–719 RSC.
- J. R. Velicogna, E. E. Ritchie, R. P. Scroggins and J. I. Princz, A comparison of the effects of silver nanoparticles and silver nitrate on a suite of soil dwelling organisms in two field soils, Nanotoxicology, 2016, 10, 1144–1151 CrossRef CAS PubMed.
- J. Garcia-Alonso, F. R. Khan, S. K. Misra, M. Turmaine, B. D. Smith, P. S. Rainbow, S. N. Luoma and E. Valsami-Jones, Cellular Internalization of Silver Nanoparticles in Gut Epithelia of the Estuarine Polychaete Nereis diversicolor, Environ. Sci. Technol., 2011, 45, 4630–4636 CrossRef CAS PubMed.
- P. S. Tourinho, C. A. M. van Gestel, A. J. Morgan, P. Kille, C. Svendsen, K. Jurkschat, J. F. W. Mosselmans, A. M. V. M. Soares and S. Loureiro, Toxicokinetics of Ag in the terrestrial isopod Porcellionides pruinosus exposed to Ag NPs and AgNO3 via soil and food, Ecotoxicology, 2016, 25, 267–278 CrossRef CAS PubMed.
- M. J. C. van der Ploeg, R. D. Handy, P. L. Waalewijn-Kool, J. H. J. van den Berg, Z. E. Herrera Rivera, J. Bovenschen, B. Molleman, J. M. Bavco, P. Tromp, R. J. B. Peters, G. F. Koopmans, I. M. C. M. Rietjens and N. W. van den Brink, Effects of silver nanoparticles (NM-300K) on Lumbricus rubellus earthworms and particle characterisation in relevant test matrices, including soil, Environ. Toxicol. Chem., 2014, 33, 743–752 CrossRef CAS PubMed.
- P. L. Waalewijn-Kool, K. Klein, R. M. Fornies and C. A. M. van Gestel, Bioaccumulation and toxicity of silver nanoparticles and silver nitrate to the soil arthropod Folsomia candida, Ecotoxicology, 2014, 23, 1629–1637 CrossRef CAS PubMed.
- L. M. Rossbach, B. J. Shaw, D. Piegza, W. F. Vevers, A. J. Atfield and R. D. Handy, Sub-lethal effects of waterborne exposure to copper nanoparticles compared to copper sulphate on the shore crab (Carcinus maenas), Aquat. Toxicol., 2017, 191, 245–255 CrossRef CAS PubMed.
- S. Makama, R. Peters, A. Undas and N. W. van den Brink, A novel method for the quantification, characterisation and speciation of silver nanoparticles in earthworms exposed in soil, Environ. Chem., 2015, 12, 643–651 CrossRef CAS.
- M. Baccaro, A. K. Undas, J. de Vriendt, J. H. J. van den Berg, R. J. B. Peters and N. W. van den Brink, Ageing, dissolution and biogenic formation of nanoparticles: how do these factors affect the uptake kinetics of silver nanoparticles in earthworms?, Environ. Sci.: Nano, 2018, 5, 1107–1116 RSC.
- S. Böhme, M. Baccaro, M. Schmidt, A. Potthoff, H.-J. Stark, T. Reemtsma and D. Kuhnel, Metal uptake and distribution in the zebrafish (Danio rerio) embryo: differences between nanoparticles and metal ions, Environ. Sci.: Nano, 2017, 4, 1005–1015 RSC.
- J. M. Unrine, S. E. Hunyadi, O. V. Tsyusko, W. Rao, W. A. Shoults-Wilson and P. M. Bertsch, Evidence for Bioavailability of Au Nanoparticles from Soil and Biodistribution within Earthworms (Eisenia fetida), Environ. Sci. Technol., 2010, 44, 8308–8313 CrossRef CAS PubMed.
- M. Baalousha, G. Cornelis, T. A. J. Kuhlbusch, I. Lynch, C. Nickel, W. Peijnenburg and N. W. van den Brink, Modeling nanomaterial fate and uptake in the environment: current knowledge and future trends, Environ. Sci.: Nano, 2016, 3, 323–345 RSC.
- C. Schultz, K. Powell, A. Crossley, K. Jurkschat, P. Kille, A. J. Morgan, D. Read, W. Tyne, E. Lahive, C. Svendsen and D. J. Spurgeon, Analytical approaches to support current understanding of exposure, uptake and distributions of engineered nanoparticles by aquatic and terrestrial organisms, Ecotoxicology, 2015, 24, 239–261 CrossRef CAS PubMed.
- A. T. Wray and S. J. Klaine, Modeling the influence of physicochemical properties on gold nanoparticle uptake and elimination by Daphnia magna, Environ. Toxicol. Chem., 2015, 34, 860–872 CrossRef CAS PubMed.
- F. R. Khan, K. B. Paul, A. D. Dybowska, E. Valsami-Jones, J. R. Lead, V. Stone and T. F. Fernandes, Accumulation Dynamics and Acute Toxicity of Silver Nanoparticles to Daphnia magna and Lumbriculus variegatus: Implications for Metal Modeling Approaches, Environ. Sci. Technol., 2015, 49, 4389–4397 CrossRef CAS PubMed.
- M. Diez-Ortiz, E. Lahive, P. Kille, K. Powell, A. J. Morgan, K. Jurkschat, C. A. M. Van Gestel, J. F. W. Mosselmans, C. Svendsen and D. J. Spurgeon, Uptake routes and toxicokinetics of silver nanoparticles and silver ions in the earthworm Lumbricus rubellus, Environ. Toxicol. Chem., 2015, 34, 2263–2270 CrossRef CAS PubMed.
- M.-N. Croteau, S. K. Misra, S. N. Luoma and E. Valsami-Jones, Silver Bioaccumulation Dynamics in a Freshwater Invertebrate after Aqueous and Dietary Exposures to Nanosized and Ionic Ag, Environ. Sci. Technol., 2011, 45, 6600–6607 CrossRef CAS PubMed.
- R. D. Handy, G. Cornelis, T. Fernandes, O. Tsyusko, A. Decho, T. Sabo-Attwood, C. Metcalfe, J. A. Steevens, S. J. Klaine, A. A. Koelmans and N. Horne, Ecotoxicity test methods for engineered nanomaterials: Practical experiences and recommendations from the bench, Environ. Toxicol. Chem., 2012, 31, 15–31 CrossRef CAS PubMed.
- A. Praetorius, N. Tufenkji, K.-U. Goss, M. Scheringer, F. von der Kammer and M. Elimelech, The road to nowhere: equilibrium partition coefficients for nanoparticles, Environ. Sci.: Nano, 2014, 1, 317–323 RSC.
- E. E. Ruppert, R. S. Fox and R. D. Barnes, Invertebrate Zoology: A Functional Evolutionary Approach, Thomson-Brooks/Cole, Belmont, CA, 7th edn, 2004 Search PubMed.
- P. Cardoso, T. L. Erwin, P. A. V. Borges and T. R. New, The seven impediments in invertebrate conservation and how to overcome them, Biol. Conserv., 2011, 144, 2647–2655 CrossRef.
- R. G. Morgado, S. Loureiro, M. N. González-Alcaraz, A. C. Duarte, A. Cachada and T. Rocha-Santos, in Soil Pollution, Academic Press, 2018, pp. 59–87, DOI:10.1016/B978-0-12-849873-6.00003-0.
- K. W. Cummins and M. J. Klug, Feeding ecology of stream invertebrates, Annu. Rev. Ecol. Syst., 1979, 10, 147–172 CrossRef.
- K. W. Cummins, R. W. Merritt and P. C. N. Andrade, The use of invertebrate functional groups to characterize ecosystem attributes in selected streams and rivers in south Brazil, Stud. Neotrop. Fauna Environ., 2005, 40, 69–89 CrossRef.
- R. C. Brusca, W. Moore and S. M. Shuster, Invertebrates, Sinauer Associates, Sunderland, Massachusetts U.S.A., 3rd edn, 2016 Search PubMed.
- A. Laycock, M. Diez-Ortiz, F. Larner, A. Dybowska, D. Spurgeon, E. Valsami-Jones, M. Rehkämper and C. Svendsen, Earthworm Uptake Routes and Rates of Ionic Zn and ZnO Nanoparticles at Realistic Concentrations, Traced Using Stable Isotope Labeling, Environ. Sci. Technol., 2016, 50, 412–419 CrossRef CAS PubMed.
- J. R. Velicogna, D. M. Schwertfeger, A. H. Jesmer, R. P. Scroggins and J. I. Princz, The bioaccumulation of silver in Eisenia andrei exposed to silver nanoparticles and silver nitrate in soil, NanoImpact, 2017, 6, 11–18 CrossRef.
- R. D. Handy and F. B. Eddy, in Physicochemical Kinetics and Transport at Biointerfaces, ed. J. Buffle, H. P. Leeuwen and W. Koster, John Wiley, Chisester, 2004, DOI:10.1002/0470094044.ch7.
- F. R. Khan, S. K. Misra, N. R. Bury, B. D. Smith, P. S. Rainbow, S. N. Luoma and E. Valsami-Jones, Inhibition of potential uptake pathways for silver nanoparticles in the estuarine snail Peringia ulvae, Nanotoxicology, 2015, 9, 493–501 CrossRef CAS PubMed.
- G. Cornelis, K. Hund-Rinke, T. Kuhlbusch, N. van den Brink and C. Nickel, Fate and Bioavailability of Engineered Nanoparticles in Soils: A Review, Crit. Rev. Environ. Sci. Technol., 2014, 44, 2720–2764 CrossRef CAS.
- M. J. van der Ploeg, J. H. van den Berg, S. Bhattacharjee, L. H. de Haan, D. S. Ershov, R. G. Fokkink, H. Zuilhof, I. M. Rietjens and N. W. van den Brink, In vitro nanoparticle toxicity to rat alveolar cells and coelomocytes from the earthworm Lumbricus rubellus, Nanotoxicology, 2014, 8, 28–37 CrossRef CAS PubMed.
- I. Lynch, A. Salvati and K. A. Dawson, PROTEIN-NANOPARTICLE INTERACTIONS What does the cell see?, Nat. Nanotechnol., 2009, 4, 546–547 CrossRef CAS PubMed.
- D. Drobne and A. Fajgelj, Use of Tc-99m-Pertechnetate to follow liquid water uptake by Procellio Scaber, J. Exp. Biol., 1993, 178, 275–279 Search PubMed.
- H. M. Fox, Anal and oral intake of water by crustacea, J. Exp. Biol., 1952, 29, 583–599 Search PubMed.
- M. R. Warbur, Evolutionary Biology of Land Isopods, Springer-Verlag, Berlin, Germany, 1993 Search PubMed.
- M. G. Vijver, J. P. M. Vink, C. J. H. Miermans and C. A. M. van Gestel, Oral sealing using glue: A new method to distinguish between intestinal and dermal uptake of metals in earthworms, Soil Biol. Biochem., 2003, 35, 125–132 CrossRef CAS.
- S. P. Hopkin, Biology of the Springtails (Insecta: Collembola), OUP Oxford, Oxford, UK, 1997 Search PubMed.
- G. Eisenbeis, Physiological absorption of liquid water by collembola - absorption by the ventral tube at different salinities, J. Insect Physiol., 1982, 28, 11–20 CrossRef.
- T. Mesaric, C. Gambardella, T. Milivojevic, M. Faimali, D. Drobne, C. Falugi, D. Makovec, A. Jemec and K. Sepcic, High surface adsorption properties of carbon-based nanomaterials are responsible for mortality, swimming inhibition, and biochemical responses in Artemia salina larvae, Aquat. Toxicol., 2015, 163, 121–129 CrossRef CAS PubMed.
- J. Baumann, J. Koser, D. Arndt and J. Filser, The coating makes the difference: Acute effects of iron oxide nanoparticles on Daphnia magna, Sci. Total Environ., 2014, 484, 176–184 CrossRef CAS PubMed.
- S. Novak, A. Jemec Kokalj, M. Hočevar, M. Godec and D. Drobne, The significance of nanomaterial post-exposure responses in Daphnia magna standard acute immobilisation assay: Example with testing TiO2 nanoparticles, Ecotoxicol. Environ. Saf., 2018, 152, 61–66 CrossRef CAS PubMed.
- S. R. Tangaa, H. Selck, M. Winther-Nielsen and F. R. Khan, Trophic transfer of metal-based nanoparticles in aquatic environments: a review and recommendations for future research focus, Environ. Sci.: Nano, 2016, 3, 966–981 RSC.
- K. Tervonen, G. Waissi, E. J. Petersen, J. Akkanen and J. V. K. Kukkonen, Analysis of fullerene-C60 and kinetic measurements for its accumulation and depuration in Daphnia magna, Environ. Toxicol. Chem., 2010, 29, 1072–1078 CAS.
- W.-C. Hou, P. Westerhoff and J. D. Posner, Biological accumulation of engineered nanomaterials: a review of current knowledge, Environ. Sci.: Processes Impacts, 2013, 15, 103–122 RSC.
- T. L. Rocha, T. Gomes, J. P. Pinheiro, V. S. Sousa, L. M. Nunes, M. R. Teixeira and M. J. Bebianno, Toxicokinetics and tissue distribution of cadmium-based Quantum Dots in the marine mussel Mytilus galloprovincialis, Environ. Pollut., 2015, 204, 207–214 CrossRef CAS PubMed.
- C. A. Garcia-Negrete, J. Blasco, M. Volland, T. C. Rojas, M. Hampel, A. Lapresta-Fernandez, M. C. J. de Haro, M. Soto and A. Fernandez, Behaviour of Au-citrate nanoparticles in seawater and accumulation in bivalves at environmentally relevant concentrations, Environ. Pollut., 2013, 174, 134–141 CrossRef CAS PubMed.
- T. Gomes, C. G. Pereira, C. Cardoso, J. P. Pinheiro, I. Cancio and M. J. Bebianno, Accumulation and toxicity of copper oxide nanoparticles in the digestive gland of Mytilus galloprovincialis, Aquat. Toxicol., 2012, 118, 72–79 CrossRef PubMed.
- Y. H. Shu, G. R. Zhang and J. W. Wang, Response of the common cutworm Spodoptera litura to zinc stress: Zn accumulation, metallothionein and cell ultrastructure of the midgut, Sci. Total Environ., 2012, 438, 210–217 CrossRef CAS PubMed.
- S. Novak, D. Drobne, M. Golobič, J. Zupanc, T. Romih, A. Gianoncelli, M. Kiskinova, B. Kaulich, P. Pelicon, P. Vavpetič, L. Jeromel, N. Ogrinc and D. Makovec, Cellular Internalization of Dissolved Cobalt Ions from Ingested CoFe2O4 Nanoparticles: In Vivo Experimental Evidence, Environ. Sci. Technol., 2013, 47, 5400–5408 CrossRef CAS PubMed.
- I. Giska, C. A. M. van Gestel, B. Skip and R. Laskowski, Toxicokinetics of metals in the earthworm Lumbricus rubellus exposed to natural polluted soils - relevance of laboratory tests to the field situation, Environ. Pollut., 2014, 190, 123–132 CrossRef CAS PubMed.
- M. Diez-Ortiz, I. Giska, M. Groot, E. M. Borgman and C. A. M. Van Gestel, Influence of soil properties on molybdenum uptake and elimination kinetics in the earthworm Eisenia andrei, Chemosphere, 2010, 80, 1036–1043 CrossRef CAS PubMed.
- L. Dai, Toxicity and bioaccumulation of silver and copper oxide nanoparticles in two deposit feeders, a polychaete Capitella teleta and a mollusk, Macoma balthica, compared to other metallic forms, Roskilde University, Roskilde, Denmark, 2013 Search PubMed.
- R. Dallinger and P. S. Rainbow, Ecotoxicology of metals in invertebrates, Taylor and Francis, Bosa Roca, United States, 1993 Search PubMed.
- P. S. Rainbow, Trace metal concentrations in aquatic invertebrates: why and so what?, Environ. Pollut., 2002, 120, 497–507 CrossRef CAS PubMed.
- S. H. Liang, S. C. Chen, C. Y. Chen, C. M. Kao, J. I. Yang, B. S. Shieh, J. H. Chen and C. C. Chen, Cadmium-induced earthworm metallothionein-2 is associated with metal accumulation and counteracts oxidative stress, Pedobiologia, 2011, 54, 333–340 CrossRef CAS.
- P. S. Rainbow, Trace metal bioaccumulation: Models, metabolic availability and toxicity, Environ. Int., 2007, 33, 576–582 CrossRef CAS PubMed.
- F. R. Khan, S. K. Misra, J. Garcia-Alonso, B. D. Smith, S. Strekopytov, P. S. Rainbow, S. N. Luoma and E. Valsami-Jones, Bioaccumulation Dynamics and Modeling in an Estuarine Invertebrate Following Aqueous Exposure to Nanosized and Dissolved Silver, Environ. Sci. Technol., 2012, 46, 7621–7628 CrossRef CAS PubMed.
- N. Adam, F. Leroux, D. Knapen, S. Bals and R. Blust, The uptake and elimination of ZnO and CuO nanoparticles in Daphnia magna under chronic exposure scenarios, Water Res., 2015, 68, 249–261 CrossRef CAS PubMed.
- F. R. Khan, G. M. Kennaway, M. N. Croteau, A. Dybowska, B. D. Smith, A. J. A. Nogueira, P. S. Rainbow, S. N. Luoma and E. Valsami-Jones, In vivo retention of ingested Au NPs by Daphnia magna: No evidence for trans-epithelial alimentary uptake, Chemosphere, 2014, 100, 97–104 CrossRef CAS PubMed.
- F. Ribeiro, C. A. M. Van Gestel, M. D. Pavlaki, S. Azevedo, A. M. V. M. Soares and S. Loureiro, Bioaccumulation of silver in Daphnia magna: Waterborne and dietary exposure to nanoparticles and dissolved silver, Sci. Total Environ., 2017, 574, 1633–1639 CrossRef CAS PubMed.
- K. R. Timmermans and P. A. Walker, The fate of trace-metals during the metamorphosis of chironomids (Diptera, Chironomidae), Environ. Pollut., 1989, 62, 73–85 CrossRef CAS PubMed.
- M. O. Montes, S. K. Hanna, H. S. Lenihan and A. A. Keller, Uptake, accumulation, and biotransformation of metal oxide nanoparticles by a marine suspension-feeder, J. Hazard. Mater., 2012, 225, 139–145 CrossRef PubMed.
- S. Thakali, H. E. Allen, D. M. Di Toro, A. A. Ponizovsky, C. P. Rooney, F.-J. Zhao, S. P. McGrath, P. Criel, H. Van Eeckhout, C. R. Janssen, K. Oorts and E. Smolders, Terrestrial Biotic Ligand Model. 2. Application to Ni and Cu Toxicities to Plants, Invertebrates, and Microbes in Soil, Environ. Sci. Technol., 2006, 40, 7094–7100 CrossRef CAS PubMed.
- M. M. Ardestani and C. A. M. van Gestel, Using a toxicokinetics approach to explain the effect of soil pH on cadmium bioavailability to Folsomia candida, Environ. Pollut., 2013, 180, 122–130 CrossRef CAS PubMed.
- M.-N. Croteau, S. K. Misra, S. N. Luoma and E. Valsami-Jones, Bioaccumulation and Toxicity of CuO Nanoparticles by a Freshwater Invertebrate after Waterborne and Dietborne Exposures, Environ. Sci. Technol., 2014, 48, 10929–10937 CrossRef CAS PubMed.
- J. C. McGeer, K. V. Brix, J. M. Skeaff, D. K. DeForest, S. I. Brigham, W. J. Adams and A. Green, Inverse relationship between bioconcentration factor and exposure concentration for metals: Implications for hazard assessment of metals in the aquatic environment, Environ. Toxicol. Chem., 2003, 22, 1017–1037 CrossRef CAS PubMed.
- S. N. Luoma and P. S. Rainbow, Why Is Metal Bioaccumulation So Variable? Biodynamics as a Unifying Concept, Environ. Sci. Technol., 2005, 39, 1921–1931 CrossRef CAS PubMed.
- M. M. Ardestani, N. M. van Straalen and C. A. M. van Gestel, Uptake and elimination kinetics of metals in soil invertebrates: A review, Environ. Pollut., 2014, 193, 277–295 CrossRef CAS PubMed.
- M. G. Vijver, C. A. M. van Gestel, N. M. van Straalen, R. P. Lanno and W. J. G. M. Peijnenburg, Biological significance of metals partitioned to subcellular fractions within earthworms (Aporrectodea caliginosa), Environ. Toxicol. Chem., 2006, 25, 807–814 CrossRef CAS PubMed.
- M. G. Vijver, J. P. M. Vink, T. Jager, N. M. Van Straalen, H. T. Wolterbeek and C. A. M. Van Gestel, Kinetics of Zn and Cd accumulation in the isopod Porcellio scaber exposed to contaminated soil and/or food, Soil Biol. Biochem., 2006, 38, 1554–1563 CrossRef CAS.
- T. Ramskov, A. Thit, M.-N. Croteau and H. Selck, Biodynamics of copper oxide nanoparticles and copper ions in an oligochaete – Part I: Relative importance of water and sediment as exposure routes, Aquat. Toxicol., 2015, 164, 81–91 CrossRef CAS PubMed.
- M. N. Croteau, S. N. Luoma, B. R. Topping and C. B. Lopez, Stable metal isotopes reveal copper accumulation and less dynamics in the freshwater bivalve corbicula, Environ. Sci. Technol., 2004, 38, 5002–5009 CrossRef CAS PubMed.
- A. Taghavy, A. Mittelman, Y. Wang, K. D. Pennell and L. M. Abriola, Mathematical Modeling of the Transport and Dissolution of Citrate-Stabilized Silver Nanoparticles in Porous Media, Environ. Sci. Technol., 2013, 47, 8499–8507 CAS.
- C. Levard, B. C. Reinsch, F. M. Michel, C. Oumahi, G. V. Lowry and G. E. Brown, Sulfidation Processes of PVP-Coated Silver Nanoparticles in Aqueous Solution: Impact on Dissolution Rate, Environ. Sci. Technol., 2011, 45, 5260–5266 CrossRef CAS PubMed.
- L. Zhan, MSc thesis, Louisiana State University and Agricultural and Mechanical College, 2013 Search PubMed.
- D. M. Di Toro, H. E. Allen, H. L. Bergman, J. S. Meyer, P. R. Paquin and R. C. Santore, Biotic ligand model of the acute toxicity of metals. 1. Technical basis, Environ. Toxicol. Chem., 2001, 20, 2383–2396 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c8en01122b |
| This journal is © The Royal Society of Chemistry 2019 |