
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceVisible-range hemi-indigo photoswitch: ON–OFF fluorescent binder for HIV-1 RNA†
Daria V.
Berdnikova

Universität Siegen, Organische Chemie II, Adolf-Reichwein-Str. 2, 57076 Siegen, Germany. E-mail: berdnikova@chemie-bio.uni-siegen.de
First published on 22nd June 2019
Abstract
A proof-of-principle for the application of hemi-indigo derivatives as RNA binders with photocontrollable fluorescence is presented. The photoswitch binds to the human immunodeficiency virus type 1 (HIV-1) RNA with a significant light-up effect. The fluorescence of the RNA-bound ligand can be reversibly switched ON and OFF by light without destroying the ligand–RNA associates.
Application of photoswitches to control the structure and functions of biomolecules is an emerging research area in the interdisciplinary field between chemistry, biology and medicine.1–4 Specifically, significant progress has been made in the photoinduced manipulation of nucleic acids.1–4 However, in the majority of cases the photoregulation of nucleic acids was accomplished by a covalent integration of photoswitches that results in the disturbance of the native structure.1–4 At the same time, examples of photoactive noncovalent nucleic acid binders are rather limited and mainly related to DNA.5–15 Thus, to the best of my knowledge, there are only a few examples of photoswitchable noncovalent RNA-binders, including the derivatives of benzo-dihydropyrene,16 spiropyrans17,18 and azobenzene-containing peptides.19 Additionally, these studies were mainly performed on synthetic RNA aptamers. Therefore, except for one example,18 no studies on noncovalent interactions of organic photoswitches with naturally occurring RNAs are available. However, due to their multiple functions in cells,20–22 natural RNAs represent an important target for small organic molecules as analytical probes or therapeutic agents,23–26 and the development of novel functional RNA binders is a topical interdisciplinary task.23,27
The aim of this work was to explore the field of noncovalent photoactive binders for naturally occurring RNAs and adopt a new photochromic scaffold for this. As a result, a novel type of photoswitchable RNA binders was developed on the basis of a hemi-indigo photoswitch. Hemi-indigo has not been used for nucleic acid studies previously, although structurally related aurones were reported as fluorescent DNA stains, however, without studies of photoswitching in the presence of DNA.28 Despite the fact that hemi-indigo was synthesized for the first time more than 100 years ago,29 its photoswitching properties have remained essentially unexplored up to now.30–33 At the same time, hemi-indigo possesses valuable features for bioapplications, namely, switching with visible light only and high bistability.30,31 In this work, a hemi-indigo derivative was designed and studied as an HIV-1 RNA binder with photoswitchable fluorescent properties.
Hemi-indigo derivative 1 was obtained as a pure Z-isomer with 24% yield by O-alkylation of vanilline with 3-chloro-N,N-dimethylpropylamine hydrochloride followed by aldol condensation of the resulting benzaldehyde derivative 2 with indoxyl acetate (Scheme 1). Product Z-1 was identified and fully characterized by NMR spectroscopy, mass spectrometry and elemental analysis (cf. the ESI†). The configuration of Z-1 was confirmed by 2D ROESY NMR (Fig. S24, ESI†). Hemi-indigo Z-1 is well soluble in water and its photochromic properties were tested in aqueous medium. The most pronounced Z–E conversion of Z-1 was achieved upon irradiation with 470 nm light (Fig. S1, ESI†). The rather fast photoreaction reached the photostationary state PSS470 in 3 min (Fig. 1). The backward switching of E-1 from PSS470 upon irradiation with 590 nm light occurred within 3.5 h (Fig. 1). Evaluation of the extent of the conversion in PSS470 and PSS590 by the Fischer method34 showed that the Z–E conversion of Z-1 is almost independent of the ionic strength of the medium (Table 1; Section 3.6, ESI†). The half-life of the E-form of 1 in the buffer solution is 10.8 h.
 | ||
| Scheme 1 Synthesis of hemi-indigo 1. | ||
 | ||
| Fig. 1 Photoinduced Z–E (blue) and E–Z (red) isomerization of hemi-indigo derivative Z-1 in water (c = 15 μM). | ||
| Species | λ abs (Z)/nm | λ abs (E)a/nm | λ fl (Z)/nm | I/I0b | Φ fl | t | PSS470/(%Z/E)a | PSS590/(%Z/E)a |
ϕ
Z–E/ϕE–Z/10−2![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) e e |
τ
25°C
/h |
K
A
/105 M−1 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| a Calculated according to Fischer [ref. 34]. b Emission light-up effect. c Fluorescence quantum yield determined relative to Rhodamine 6G in EtOH (Φ = 0.95) [ref. 39]. d Forward (at 470 nm) and backward (at 590 nm) reaction times. e Photoisomerization quantum yields of the forward ϕZ–E (at 470 nm) and backward ϕE–Z (at 590 nm) reactions. f Half-lifes of the E-form at 25 °C. g Association constants with RNA. h Could not be determined due to the very low emission intensity. i Could not be determined due to the presence of several equilibria. j Binding did not reach saturation. k The data were recorded at cTAR–Tat/c1 = 4. | |||||||||||
| 1 | 478 | 514 | 595–598h | — | n.d.h | 3 min/3.5 h | 20/80 | 97/3 | 2.7/0.20 | 10.8 | — |
| 1–TAR | 488 | 523 | 563 | 47 | 0.010 | 3 min/3.5 h | 23/77 | 92/8 | 2.1/0.06 | n.d.i | 1.1 |
| 1–RRE | 493 | 529 | 565 | 75 | 0.015 | 3 min/3.5 h | 29/71 | 89/11 | 1.2/0.02 | 51.0 | 1.7 |
| 1–TAR–Tatj | 486k | 522k | 562k | 50k | n.d.j | 3 min/5 h | n.d.j | n.d.j | n.d.j | n.d.j | n.d.j |
Two elements of human immunodeficiency virus type 1 (HIV-1) RNA were chosen as targets, namely, the transactivation response element (TAR) and the stem IIB region of the Rev response element (RRE-IIB) because these RNA structures play a crucial role in the regulation of HIV-1 gene expression (Fig. 2).35 TAR triggers the transcription of HIV in infected cells upon complexation with Tat protein.36 The truncated Tat peptide comprising the basic subdomain (47–57) was used because its TAR-binding properties resemble those of the full-length protein.37 RRE-IIB acts as a switch between viral latency and active replication.38
 | ||
| Fig. 2 Structures of HIV-1 TAR, RRE-IIB and the Tat peptide (47–57). | ||
Firstly, the interactions of hemi-indigo Z-1 with TAR and RRE RNA were studied by absorption spectroscopy. Photometric titrations of Z-1 with TAR and RRE-IIB RNA revealed a hypochromic effect along with a moderate bathochromic shift of the absorption maximum in both cases (Fig. 3A1, A2 and Table 1). The extent of the bathochromic shift was clearly dependent on the RNA type due to the different microenvironment of each sequence. In the case of TAR RNA, an isosbestic point was not formed, indicating that Z-1 changes its binding sites within the oligonucleotide depending on the ligand-to-RNA ratio (Fig. 3A1). In the case of RRE-IIB RNA, the distinct isosbestic point indicated a single dominating binding mode of Z-1 (Fig. 3A2). Interestingly, on titration of a complex of TAR RNA and Tat peptide (Tat–TAR) to the solution of hemi-indigo Z-1, the absorption maximum of the ligand initially shifted bathochromically with a slight decrease followed by an increase of the absorbance without reaching saturation; no isosbestic points were formed either (Fig. 3A3). Such behavior indicates the presence of several binding equilibria that may arise from a non-specific interaction of Z-1 with the Tat–TAR complex or multiple binding sites. Overall, no selectivity of hemi-indigo Z-1 towards a certain element of HIV-1 RNA was observed.
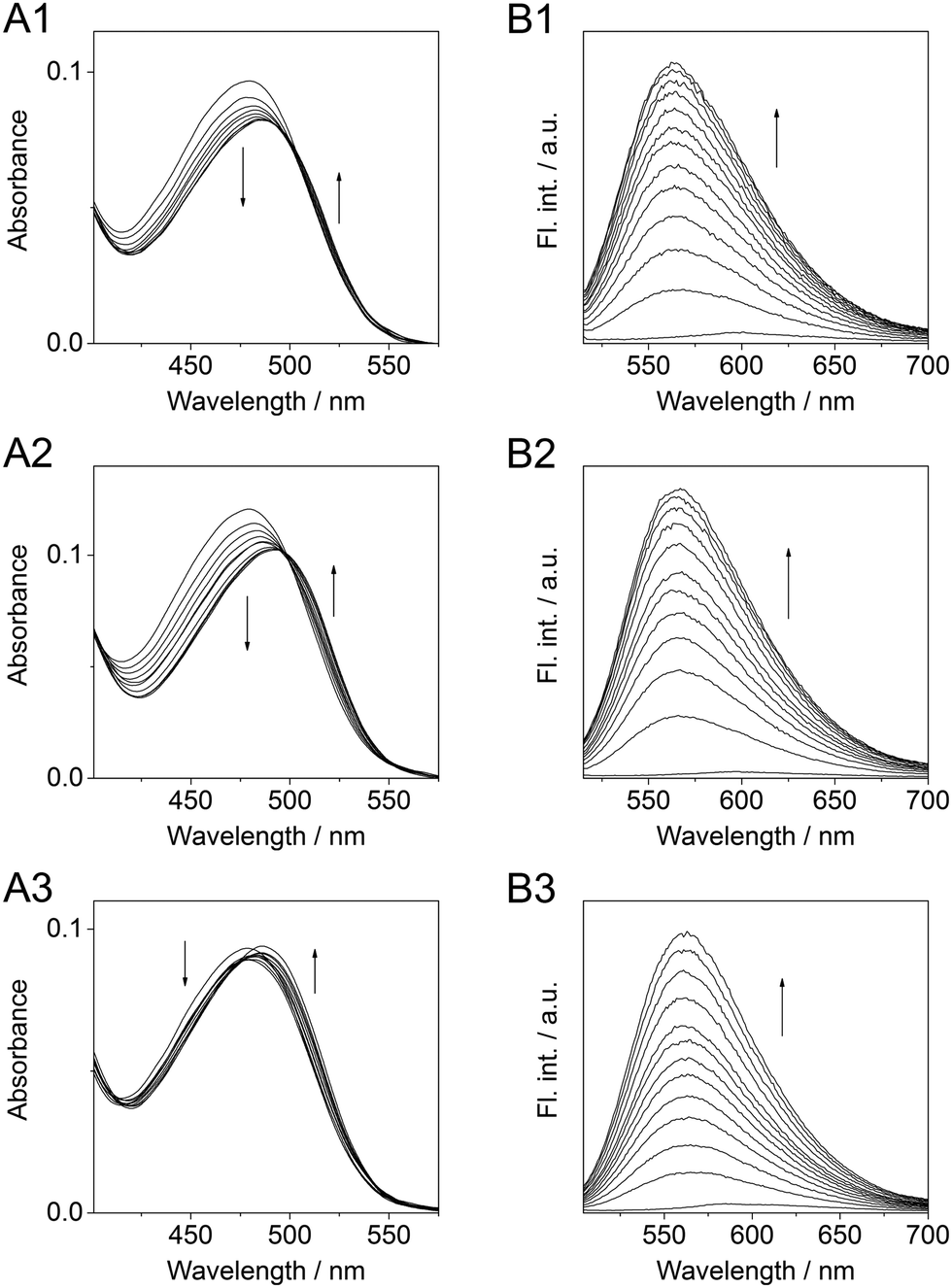 | ||
| Fig. 3 (A) Spectrophotometric titrations of Z-1 with TAR (A1, c1 = 0.9 mM, cTAR/c1 = 0–3), RRE-IIB (A2, c1 = 1.2 mM, cRRE/c1 = 0–3.5) and the Tat–TAR complex (A3, c1 = 0.9 mM, cTat-TAR/c1 = 0–4). (B) Fluorimetric titrations of hemi-indigo Z-1 (c1 = 10 μM) with TAR (B1, cTAR/c1 = 0–3, λex = 498 nm), RRE-IIB (B2, cRRE/c1 = 0–4, λex = 497 nm) and the Tat–TAR complex (B3, cTat–TAR/c1 = 0–4, λex = 485 nm). In all cases: 10 mM Na-phosphate buffer with 0.1 M NaCl and 1 mM EDTA, pH = 7, 20 °C; incubation time after addition of every aliquot: 1 min. | ||
Both isomers E-1 and Z-1 are non-fluorescent in aqueous medium (Fig. S7, ESI†); however, upon binding to TAR and RRE RNA the fluorescence of Z-1 increases drastically (Fig. 3B1, B2 and Table 1). Thus, binding to TAR RNA produced a 47-fold enhancement of the fluorescence, whereas association with RRE-IIB RNA led to a 75-fold light-up. A possible reason for the light-up effect is the suppression of the rotational relaxation of the ligand within the RNA binding site.33 Although the fluorescence quantum yields of the RNA-bound forms are not high (Table 1), the light-up effect can be detected by the naked eye (Fig. 4B). The analysis of the isotherms resulting from the fluorimetric titrations (Fig. S18, ESI†) allowed to estimate the binding constants and stoichiometries of the ligand–RNA complexes. Thus, in both cases the preferential formation of 1:1 ligand–RNA oligonucleotide complexes was deduced. The values of the overall association constants are K = 1.1 × 105 M−1 for Z-1–TAR and K = 1.7 × 105 M−1 for Z-1–RRE. The interaction of Z-1 with Tat–TAR also resulted in a pronounced light-up effect of the ligand molecule (Fig. 3B3). However, such as the photometric analysis, the fluorimetric titration did not lead to saturation; therefore, the binding constant could not be calculated. Displacement analysis with the Tat peptide showed that both isomers of 1 do not compete with Tat, but rather the ternary associate 1–Tat–TAR is formed (Fig. S2 and S3, ESI†).
 | ||
| Fig. 4 (A) Fluorescence OFF–ON switching cycles of hemi-indigo 1 in the complexes: (a) 1–TAR, c1 = cTAR = 10 μM, λex = 495 nm, λdet = 563 nm; (b) 1–RRE, c1 = cRRE = 10 μM, λex = 498 nm, λdet = 567 nm; (c) 1–Tat–TAR, c1 = cTat–TAR = 10 μM, λex = 480 nm, λdet = 561 nm. (B) Fluorescence of (a) free Z-1 and Z-1–TAR complex before (b) and after (c) switching, λex = 365 nm. The forward reaction occurred upon irradiation with blue light (470 nm) and the backward reaction occurred upon irradiation with amber light (590 nm). | ||
Irradiation of the Z-1–TAR and Z-1–RRE complexes with 470 nm light resulted in the switching of the RNA-bound ligand to the E-form (Fig. 4). The significant bathochromic shift of the E-isomer absorption maximum in the presence of RNA indicates that it remains bound to the RNA (Table 1, Fig. S10 and S11, ESI†). The extent of the Z–E conversion of the 1–RNA complexes in PSS470 is slightly dependent on the RNA type (Table 1 and Fig. S6, ESI†). Accommodation of E-1 in the RNA binding site significantly increases its half-life, as was shown for the 1–RRE complex (Table 1; Section 3.8, ESI†). The extent of conversion for the backward E–Z switching of the E-1–RNA complexes was slightly reduced in comparison to the free state (Table 1 and Table S1, ESI†). The times of both forward and backward switching were not affected by association with RNA. However, for 1–Tat–TAR, the backward reaction time increased significantly, most likely, due to the stiffening of the ternary structure (Table 1).
Estimation of the binding sites by circular dichroism (CD) spectroscopy showed that Z-1 associates with TAR RNA in the UCU-bulge region (Fig. S14A, ESI†) that results in unstacking of the bulge bases,40 as additionally confirmed by displacement experiments with neomycin (Fig. S12, ESI†). Binding of Z-1 to RRE RNA also induced conformational changes of RNA (Fig. S14B, ESI†); however, the position of the binding site was difficult to deduce. In the case of 1–Tat–TAR, Z-1 presumably binds near the bulge from the minor groove side (Fig. S14C, ESI†). Importantly, the photoswitching of the bound ligand had just a small influence on the structures of the TAR and RRE, whereas the structural rearrangement of 1–Tat–TAR was pronounced.
Most notably, the Z–E photoswitching of the RNA-bound Z-1 upon irradiation with 470 nm led to a significant decrease of the fluorescence intensity (Fig. S6, ESI†). The residual emission intensity correlated with the calculated amount of Z-1 in PSS470 (23–29%). This indicates that, unlike Z-1, the photoisomer E-1 has very weak fluorescence upon binding to RNA. Considering that both E-1 and Z-1 remained bound to RNA, it was proposed that the system could be used as an ON–OFF fluorescent binder of viral RNA. To prove this proposal, the emission switching cycles of 1–TAR, 1–RRE and 1–Tat–TAR complexes were recorded (Fig. 4A). The extent of the emission restoration was dependent on the isomeric composition of PSS590 in each case (Table 1). Overall, the switching provided a significant difference between the ON and OFF fluorescence states of the RNA-bound ligand (Fig. 4B). The switching cycles showed good photostability of the binary complexes 1–TAR and 1–RRE over three cycles (Fig. 4A). In the case of the ternary system 1–Tat–TAR, however, photodegradation was already noticeable after the second cycle (Fig. 4A), presumably because of the much slower backward reaction that required longer irradiation times. Notably, the switching of Z-1 does not interfere with the detection of fluorescence (Fig. S9, ESI†).
In summary, a proof-of-principle for the application of the hemi-indigo scaffold as a platform for a novel class of photoswitchable RNA-binders is provided. As an RNA-binding ligand, hemi-indigo offers the following valuable features: (i) good photoswitching performance in aqueous medium; (ii) switching in both directions by visible light with significantly different wavelengths; (iii) a strong light-up effect of the emission upon binding of the Z-form to RNA; and (iv) switchable fluorescence characteristics in the RNA-bound state. Additionally, the hemi-indigo ligand forms a ternary complex with TAR RNA and Tat protein, retaining its switching properties. The photocontrollable fluorescence of hemi-indigo derivatives makes them promising candidates for the application in real-time monitoring of biological systems because photoswitchable fluorophores allow implementing novel bioimaging strategies that would be impossible with conventional fluorophores.41 Along these lines, binding of both isomers of hemi-indigo to RNA is advantageous because the ligand, therefore, can be used to study dynamic processes. Further modification of hemi-indigo with bulky substituents will be performed to realize photocontrolled manipulation of the spatial structure of RNA.
This project has received funding from the European Union's Horizon 2020 research and innovation programme, Marie Skłodowska-Curie actions (MSCA), under grant agreement No. 749788 – PHOTORNA. I thank Prof. Dr Heiko Ihmels (Universität Siegen, Germany) for fruitful discussions and valuable advices. I thank Prof. Ilarion Troitsky (Moscow Theological Academy, Russia) for his kind support.
Conflicts of interest
There are no conflicts to declare.Notes and references
- Z. L. Pianowski, Chem. – Eur. J., 2019, 25, 5128 CrossRef CAS PubMed.
- H. Cheng, J. Yoon and H. Tian, Coord. Chem. Rev., 2018, 372, 66 CrossRef CAS.
- A. S. Lubbe, W. Szymanski and B. L. Feringa, Chem. Soc. Rev., 2017, 46, 1052 RSC.
- W. Szymanski, J. M. Beierle, H. A. V. Kistemaker, W. A. Velema and B. L. Feringa, Chem. Rev., 2013, 113, 6114 CrossRef CAS PubMed.
- K. Starčević, G. Karminski-Zamola, I. Piantanida, M. Žinić, L. Šuman and M. Kralj, J. Am. Chem. Soc., 2005, 127, 1074 CrossRef PubMed.
- (a) C. Dohno, S. Uno and K. Nakatani, J. Am. Chem. Soc., 2007, 129, 11898 CrossRef CAS PubMed; (b) C. Dohno, T. Yamamoto and K. Nakatani, Eur. J. Org. Chem., 2009, 4051 CrossRef CAS.
- J. Andersson, S. Li, P. Lincoln and J. Andréasson, J. Am. Chem. Soc., 2008, 130, 11836 CrossRef CAS PubMed.
- M. L. Di Pietro, F. Puntoriero, F. Tuyéras, P. Ochsenbein, P. P. Lainé and S. Campagna, Chem. Commun., 2010, 46, 5169 RSC.
- M. I. Sánchez, O. Vázquez, M. E. Vázquez and J. L. Mascareñas, Chem. Commun., 2011, 47, 11107 RSC.
- A. Mammana, G. T. Carroll, J. Areephong and B. L. Feringa, J. Phys. Chem. B, 2011, 115, 11581 CrossRef CAS PubMed.
- S. V. Paramonov, V. Lokshin, H. Ihmels and O. A. Fedorova, Photochem. Photobiol. Sci., 2011, 10, 1279 RSC.
- H. Ihmels, J. Mattay, F. May and L. Thomas, Org. Biomol. Chem., 2013, 11, 5184 RSC.
- M. Hammarson, J. R. Nilsson, S. Li, P. Lincoln and J. Andréasson, Chem. – Eur. J., 2014, 20, 15855 CrossRef CAS PubMed.
- (a) D. V. Berdnikova, T. M. Aliyeu, T. Paululat, Y. V. Fedorov, O. A. Fedorova and H. Ihmels, Chem. Commun., 2015, 51, 4906 RSC; (b) D. Berdnikova, O. Fedorova, E. Gulakova and H. Ihmels, Chem. Commun., 2012, 48, 4603 RSC.
- M. P. O’Hagan, S. Haldar, M. Duchi, T. A. A. Oliver, A. J. Mulholland, J. C. Morales and M. C. Galan, Angew. Chem., Int. Ed., 2019, 58, 4334 CrossRef PubMed.
- H. W. Lee, S. G. Robinson, S. Bandyopadhyay, R. H. Mitchel and D. Sen, J. Mol. Biol., 2007, 371, 1163 CrossRef CAS PubMed.
- (a) D. D. Young and A. Deiters, ChemBioChem, 2008, 9, 1225 CrossRef CAS PubMed; (b) X. Zhang, J. Zhang, Y.-L. Ying, H. Tian and Y.-T. Long, Chem. Sci., 2014, 5, 2642 RSC.
- A. A. Ali, M. Kang, R. Kharbash and Y. Kim, BMC Biomed. Eng., 2019, 1, 6 CrossRef.
- (a) G. Hayashi, M. Hagihara, C. Dohno and K. Nakatani, J. Am. Chem. Soc., 2007, 129, 8678 CrossRef CAS PubMed; (b) G. Hayashi, M. Hagihara and K. Nakatani, Chem. – Eur. J., 2009, 15, 424 CrossRef CAS PubMed.
- RNA Worlds: From Life's Origins to Diversity in Gene Regulation, ed. J. F. Atkins, R. F. Gesteland, T. R. Cech, Cold Spring Harbor Laboratory Press, New York, 5th edn, 2019 Search PubMed.
- Nucleic Acids in Chemistry and Biology, ed. G. M. Blackburn, RSC Publishing, Cambridge, 3rd edn, 2006 Search PubMed.
- R. L. P. Adams, J. T. Knowler and D. P. Leader, The biochemistry of the nucleic acids, Chapman & Hall, London, 1992 Search PubMed.
- K. D. Warner, C. E. Hajdin and K. M. Weeks, Nat. Rev. Drug Discovery, 2018, 17, 547 CrossRef CAS PubMed.
- N. F. Rizvi and G. F. Smith, Bioorg. Med. Chem. Lett., 2017, 27, 5083 CrossRef CAS PubMed.
- L. Guan and M. D. Disney, ACS Chem. Biol., 2012, 7, 73 CrossRef CAS PubMed.
- J. R. Thomas and P. J. Hergenrother, Chem. Rev., 2008, 108, 1171 CrossRef CAS PubMed.
- M. You and S. R. Jaffrey, Ann. N. Y. Acad. Sci., 2015, 1352, 13 CrossRef CAS PubMed.
- N. Shanker, O. Dilek, K. Mukherjee, D. W. McGee and S. L. Bane, J. Fluoresc., 2011, 21, 2173 CrossRef CAS PubMed.
- A. Baeyer, Ber. Dtsch. Chem. Ges., 1883, 16, 2188 CrossRef.
- C. Petermayer, S. Thumser, F. Kink, P. Mayer and H. Dube, J. Am. Chem. Soc., 2017, 139, 15060 CrossRef CAS PubMed.
- C. Petermayer and H. Dube, J. Am. Chem. Soc., 2018, 140, 13558 CrossRef CAS PubMed.
- M. Ikegami and T. Arai, Bull. Chem. Soc. Jpn., 2003, 76, 1783 CrossRef CAS.
- M. Ikegami and T. Arai, Chem. Lett., 2005, 34, 492 CrossRef CAS.
- E. Fischer, J. Phys. Chem., 1967, 71, 3704 CrossRef CAS.
- J. Karn and C. M. Stoltzfus, Cold Spring Harbor Perspect. Med., 2012, 4, a00691 Search PubMed.
- J. Karn, J. Mol. Biol., 1999, 293, 235 CrossRef CAS PubMed.
- K. M. Weeks, C. Ampe, S. C. Schultz, T. A. Steitz and D. M. Crothers, Science, 1990, 249, 1281 CrossRef CAS PubMed.
- S. M. Kingsman and A. J. Kingsman, The regulation of human immunodeficiency virus type-1 gene expression in EJB Reviews, Springer, Berlin, Heidelberg, 1996 Search PubMed.
- K. H. Drexhage, Structure and properties of laser dyes in dye lasers, topics in applied physics, ed. F. P. Schäfer, Springer, Berlin, 2nd edn, 1977 Search PubMed.
- S. Wang, P. W. Huber, M. Cui, A. W. Czarnik and H.-Y. Mei, Biochemistry, 1998, 37, 5549 CrossRef CAS PubMed.
- (a) Y. Zhang, S. Tang, E. R. Thapaliya, L. Sansalone and F. M. Raymo, Chem. Commun., 2018, 54, 8799 RSC; (b) Y. Zhang, S. Tang, L. Sansalone, J. D. Baker and F. M. Raymo, Chem. – Eur. J., 2016, 22, 15027 CrossRef CAS PubMed; (c) Y. Xiong, P. Rivera-Fuentes, E. Sezgin, A. V. Jentzsch, C. Eggeling and H. L. Anderson, Org. Lett., 2016, 18, 3666 CrossRef CAS PubMed; (d) M. N. Tran and D. M. Chenoweth, Angew. Chem., Int. Ed., 2015, 54, 6442 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Synthetic procedures, optical spectroscopy and photochemical studies data, and 1H and 13C NMR spectra. See DOI: 10.1039/c9cc04270a |
| This journal is © The Royal Society of Chemistry 2019 |