
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceAnthocyanins inhibit tumor necrosis alpha-induced loss of Caco-2 cell barrier integrity†
Eleonora
Cremonini
 ab,
Angela
Mastaloudis
c,
Shelly N.
Hester
c,
Sandra V.
Verstraeten
de,
Maureen
Anderson
f,
Steven M.
Wood
c,
Andrew L.
Waterhouse
f,
Cesar G.
Fraga
*agh and
Patricia I.
Oteiza
*ab
ab,
Angela
Mastaloudis
c,
Shelly N.
Hester
c,
Sandra V.
Verstraeten
de,
Maureen
Anderson
f,
Steven M.
Wood
c,
Andrew L.
Waterhouse
f,
Cesar G.
Fraga
*agh and
Patricia I.
Oteiza
*ab
aDepartment of Nutrition, University of California, Davis, CA, USA. E-mail: poteiza@ucdavis.edu
bDepartment of Environmental Toxicology, University of California, Davis, CA, USA
cPharmanex Research, NSE Products, Inc., Provo, UT, USA
dDepartamento de Química Biológica, Facultad de Farmacia y Bioquímica, Universidad de Buenos Aires, Buenos Aires, Argentina
eInstituto de Quimica y Fisicoquímica Biológicas (IQUIFIB), CONICET-Universidad de Buenos Aires, Buenos Aires, Argentina
fDepartment of Viticulture and Enology, University of California, Davis, CA, USA
gFisicoquímica, Facultad de Farmacia y Bioquímica, Universidad de Buenos Aires, Buenos Aires, Argentina. E-mail: cfraga@ffyb.uba.ar
hInstituto de Bioquímica y Medicina Molecular (IBIMOL), CONICET-Universidad de Buenos Aires, Buenos Aires, Argentina
First published on 18th July 2017
Abstract
An increased permeability of the intestinal barrier is proposed as a major event in the pathophysiology of conditions characterized by chronic gut inflammation. This study investigated the capacity of pure anthocyanins (AC), and berry and rice extracts containing different types and amounts of AC, to inhibit tumor necrosis alpha (TNFα)-induced permeabilization of Caco-2 cell monolayers. Caco-2 cells differentiated into intestinal epithelial cell monolayers were incubated in the absence/presence of TNFα, with or without the addition of AC or AC-rich plant extracts (ACRE). AC and ACRE inhibited TNFα-induced loss of monolayer permeability as assessed by changes in transepithelial electrical resistance (TEER) and paracellular transport of FITC-dextran. In the range of concentrations tested (0.25–1 μM), O-glucosides of cyanidin, and delphinidin, but not those of malvidin, peonidin and petunidin protected the monolayer from TNFα-induced decrease of TEER and increase of FITC-dextran permeability. Cyanidin and delphinidin acted by mitigating TNFα-triggered activation of transcription factor NF-κB, and downstream phosphorylation of myosin light chain (MLC). The protective actions of the ACRE on TNFα-induced TEER increase was positively correlated with the sum of cyanidins and delphinidins (r2 = 0.83) content in the ACRE. However, no correlation was observed between TEER and ACRE total AC, malvidin, or peonidin content. Results support a particular capacity of cyanidins and delphinidins in the protection of the intestinal barrier against inflammation-induced permeabilization, in part through the inhibition of the NF-κB pathway.
Introduction
The capacity of flavonoids to inhibit inflammation and modulate select signaling cascades may be important in their protective actions at the gastrointestinal tract.1,2 In particular, they may have a relevant role in the prevention and amelioration of intestinal barrier permeabilization.3–7 However, the large number of existing flavonoids with different chemical structures and spatial configurations, and the multiple metabolites generated by both the gut microbiota, and once absorbed, by tissue metabolism,8 complicate precise nutritional recommendations and/or pharmacological strategies.Anthocyanins (AC) constitute one of the major flavonoid subgroups,9,10 which are present in and provide color to many fruits and vegetables, e.g. berries, red cabbage, and black rice. Many different AC exist, differing in the bonded sugar moieties, the number and substitution of hydroxyl groups, and the aliphatic or aromatic acids attached to the sugars.11 Growing evidence suggests that AC consumption could provide benefits to human health.12 However, their potential benefits on intestinal health, and especially under conditions of increased intestinal permeability, are poorly understood and in need of further characterization.
Intestinal barrier permeabilization and the associated increased passage of deleterious substances present in the lumen, are key contributors to the pathophysiology of different diseases (e.g. inflammatory bowel diseases, celiac and extraintestinal autoimmune diseases) and contribute to the adverse consequences of obesity and type 2 diabetes.13–16 The small intestine, and to a lesser extent the large intestine, are exposed to significant amounts of AC from foods which may protect the intestinal epithelium against different luminal insults (e.g. oxidants, inflammation products, fats, toxins, bacteria, and viruses), helping to promote intestinal health and mitigate chronic intestinal and metabolic diseases. Current evidence indicates that ingested AC are present throughout the length of the gastrointestinal tract, both as parent compounds and as metabolites resulting from action of metabolizing systems and the gut microbiota.17,18
As an initial step to define the relevance of AC intake on intestinal health, in this work we investigated the capacity of purified AC and of AC-rich plant extracts (ACRE) to prevent tumor necrosis factor alpha (TNFα)-induced permeabilization of Caco-2 cell monolayers. We also assessed the effects of individual AC on NF-κB activation, as a central signaling pathway involved in TNFα-mediated intestinal barrier permeabilization. Results indicate that AC effectively protect Caco-2 monolayers from TNFα-induced permeabilization in part through their capacity to inhibit NF-κB activation, and that cyanidins and delphinidins exhibited the most potent anti-inflammatory benefits.
Materials and methods
Materials
Caco-2 cells were from the American Type Culture Collection (Rockville, MA). Cell culture media and reagents were from Invitrogen/Life technologies (Grand Island, NY). HBSS 1× (21-022-CV) was obtained from Corning (Manassas, VA). The Millicell cell culture inserts were from EMD Millipore (Hayward, CA). Fluorescein isothiocyanate (FITC)-dextran and human TNFα were obtained from Sigma Chem. Co. (St Louis, MO). Human interferon gamma (IFN-γ) and primary antibodies for β-actin (12620), phospho-IKKα/β (Ser176/180) (2697), IKKβ (2370), phospho-p65 (Ser536) (3033), p65 (3987), phospho-myosin light chain (MLC) (Thr18/Ser19) (3674), and MLC (8505) were from Cell Signaling Technology (Danvers, MA). Delphinidin-3-O-glucoside, cyanidin-3-O-glucoside, malvidin-3-O-glucoside, peonidin-3-O-glucoside, and petunidin-3-O-glucoside were from Extrasynthese (Genay Cedex, France). Phenolic acids and flavonoids used as reference standards included syringic, vanillic, protocatechuic, 4-hydroxybenzoic, gallic acids, (+)-catechin and (−)-epicatechin were obtained from Sigma-Aldrich (St Louis, MO); 3-O-methylgallic and quercetin-3-galactose were from Extrasynthese (Genay Cedex, France). The ACRE were provided by Pharmanex Research (NSE Products, Inc., Provo, UT, USA) and included: black chokeberry extract (35% w/w AC), crowberry extract (30% w/w AC), black rice extract (20% w/w AC), wild blueberry extract (5% w/w AC), bilberry extract (36% w/w AC), domesticated blueberry extract (25% w/w AC), and red grape extract (minimum 10% w/w AC).Molecular structure calculation
The optimization of chemical structures of select AC was performed by molecular mechanics (MM2) according to Allinger19 using the routines available in ChemBio3D Ultra 11.0.1 (Cambridge Scientific Computing Inc., Cambridge, UK).Cell culture and incubations
Caco-2 cells were cultured at 37 °C and 5% (v/v) CO2 atmosphere in minimum essential medium (MEM) supplemented with 10% (v/v) fetal bovine serum, antibiotics (50 U per ml penicillin, and 50 μg per ml streptomycin), 1% (v/v) of 100× non-essential amino acids, and 1 mM sodium pyruvate. The medium was replaced every 3 d during cell growth and differentiation. For the experiments, cells were used 21 d after reaching confluence to allow for differentiation into intestinal epithelial cells. Cells were used between passages 3 and 15. All the experiments were performed in serum- and phenol red-free MEM.Transepithelial electrical resistance
To measure transepithelial electrical resistance (TEER), cells were differentiated into polarized monolayers by culture on transwell inserts (12 mm, 0.4 μm pore polyester membranes) placed in 12-well plates at a seeding density of 0.3 × 106 cells per transwell. The volume of medium added to the upper and lower compartments was 0.4 and 0.6 ml, respectively. TEER was measured using a Millicell-ERS Resistance System (Millipore, Bedford, MA) that includes a dual electrode volt-ohm-meter. TEER was calculated as: TEER = (Rm − Ri) × A; being, Rm, transmembrane resistance; Ri, intrinsic resistance of a cell-free media; and A, the surface area of the membrane in cm2. Monolayers were used when TEER values were between 350–450 Ω cm2. For the evaluation of the effects of purified AC and ACRE on TEER, Caco-2 cell monolayers were preincubated for 24 h with IFNγ (10 ng ml−1) to upregulate the TNFα receptor.20 The monolayers were then incubated for 30 min with AC (0.25, 0.5, and 1 μM) or ACRE (1–10 μg ml−1) added to the upper compartment. TNFα (5 ng ml−1) was subsequently added to the lower compartment and cells were incubated for further 6 h. Both IFNγ and TNFα were diluted in MEM before treating cells. For TEER assessment, the incubation media were removed from the upper and lower compartments, cells rinsed with HBSS 1×, and the same solution was added to both compartments. Arbitrary units (AU) were calculated based on TEER values for the non-added (control) cells.Transport of fluorescein isothiocyanate–dextran (FITC-dextran)
The paracellular transport through Caco-2 cell monolayers was determined by measuring the apical-to-basolateral clearance of FITC-dextran (4 kDa). Cells were treated as described for TEER determination; after the 6 h of incubation with TNFα, the medium was replaced in both compartments with fresh serum and phenol red-free MEM; FITC-dextran was then added to the upper compartment (100 μM final concentration). After 3.5 h, 100 μl of the medium in the lower compartment was collected and diluted with 100 μl of HBSS 1×. The fluorescence was measured at λexc: 485 nm and λem: 530 nm in a fluorescence plate reader. The FITC-dextran clearance (CLFITC) was calculated using the equation fFITC/(FFITC/A), where fFITC is flux of FITC-dextran (in fluorescence units per h); FFITC, the fluorescence of FITC-dextran in the upper compartment at zero time (in fluorescence units per nl); and A, the surface area of the membrane (1 cm2). Arbitrary units (AU) were calculated based on the CL value for the non-added (control) cells (75 nl h−1 cm−2).Western blot analysis
Cells were pre-incubated for 30 min in the absence or presence of 0.5 μM cyanidin-3-O-glucoside delphinidin-3-O-glucoside, malvidin-3-O-glucoside, peonidin-3-O-glucoside or petunidin-3-O-glucoside and then incubated for 5 min (for IKK and p65 phosphorylation) or 6 h (for MLC phosphorylation) in the absence or presence of TNFα (5 ng ml−1). Cells were collected and total homogenates were prepared as previously described.4 Aliquots of total homogenates containing 30 μg protein were denatured with Laemmli buffer, separated by reducing 10% polyacrylamide gel electrophoresis, and electroblotted onto PVDF membranes. Membranes were blocked for 1 h in 5% (w/v) bovine serum albumin and subsequently incubated in the presence of the corresponding primary antibody (1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1000 dilution) overnight at 4 °C. After incubation for 1 h at room temperature in the presence of the corresponding secondary antibody (HRP conjugated) (1:10000 dilution) the conjugates were visualized by an ECL system using a Phosphoimager 840 (Amersham Pharmacia Biotech. Inc., Piscataway, NJ).
:1000 dilution) overnight at 4 °C. After incubation for 1 h at room temperature in the presence of the corresponding secondary antibody (HRP conjugated) (1:10000 dilution) the conjugates were visualized by an ECL system using a Phosphoimager 840 (Amersham Pharmacia Biotech. Inc., Piscataway, NJ).
Determination of anthocyanidins
The ACRE were analyzed by a liquid chromatography method.21,22 ACRE powders were dissolved in water and filtered through PTFE (0.22 μ) immediately before injection. Separation was performed using an Agilent series 1200 instrument (Agilent Technologies, Santa Clara, CA) with a Kinetex F5 pentafluorophenyl HPLC column (2.6 μm, 100 × 4.6 mm) and SecurityGuard® cartridge (PFP, 4.0 × 2.0 mm) (Phenomenex, Torrance, CA). A flow rate of 0.70 mL min−1 and a column temperature of 37 °C was set. The injector temperature was 4 °C with an injection volume of 7 μL. Detection was by UV-Vis DAD (wavelength monitored at 280 and 520 nm) and ESI-MS-MS for mass spectral results. Quantification was based on DAD peak area absorbance at 520 nm. A binary gradient was employed consisting of 1.0% formic acid (v/v) in water (mobile phase A) and 1.0% formic acid (v/v) in acetonitrile (mobile phase B). Gradient was as follows: 1% B at 0 minutes, 7.5% B at 7 min, 7.6% B at 14 min, 10% B at 17 min, 12% B at 18.5 min, 30% B at 24 min, 90% B at 25 min, 1% B at 26 to 30 min. Mass spectral data were acquired using an Agilent 6430 triple-quadrupole mass spectrometer with electrospray injection (Agilent Technologies, Santa Clara, CA, USA) set to scan mode with the following optimal MS/MS source parameters: nebulizer at 40 psi, capillary voltage +4000 V (or −3500 V), gas temperature 325 °C, and flow of 5 L min. Sheath gas was 250 °C and sheath flow of 11 L min−1. AC were identified after ionization in their molecular cation form under MS positive ion mode. Parent ions (m/z) were: cyanidin-3-O-galactoside, 449; cyanidin-3-O-glucoside, 449; cyanidin-3-arabinoside, 419; cyanidin-3-O-rutinoside, 595; delphinidin-3-O-galactoside, 465; delphinidin-3-O-glucoside, 465; delphinidin-3-arabinoside, 435; delphinidin-3-O-rutinoside, 611; malvidin-3-O-galactoside, 493; malvidin-O-glucoside, 493; malvidin-3-arabinoside, 463; malvidin-3-(6-acet) glucoside, 535; malvidin-3-(6-acet) galactoside, 535; malvidin-3-(6-coumaryl) glucoside, 639; peonidin-3-O-glucoside, 463; petunidin-3-O-galactoside, 479; petunidin-3-O-glucoside, 479; petunidin-3-O-arabinoside, 449; and petunidin-3-(6-acet) glucoside, 521.Statistical analysis
Data were analyzed by one-way analysis of variance (ANOVA) and simple regression analysis using Statview 5.0 (SAS Institute Inc., Cary, NC). Fisher least significance difference test was used to examine differences between group means. A P value < 0.05 was considered statistically significant. Data are shown as mean ± SEM of at least 3 independent experiments.Results
Chemical structure and spatial conformation of the studied AC
The chemical structure and spatial conformation of the non-glycosylated forms of the studied AC are shown in Fig. 1. While rings A and C are identical for all the AC studied, the substitutions and positioning of the B ring with respect to the C ring are different. In addition to the 3′,4′ catechol group in cyanidin's B ring, delphinidin contains an additional hydroxyl group in the 5′ position, whereas petunidin has an –OCH3 group in the 5′ position. Peonidin and malvidin each have an –OCH3 and a hydroxyl group in the 3′ and 4′ positions, respectively; malvidin also has an additional –OCH3 group in the 5′ position. The calculated dihedric angles between B and C rings show values of 39, 37, 43, 34, and 39 degrees for cyanidin, delphinidin, malvidin, peonidin, and petunidin, respectively. | ||
| Fig. 1 Anthocyanidin chemical structures and spatial conformation. Carbon atoms are depicted in turquoise, oxygen atoms in red, and hydrogen atoms in grey. | ||
AC differentially protect Caco-2 cell monolayers from TNFα-induced permeabilization
The capacity of five pure AC (as glucosides) to prevent TNFα-induced permeabilization of Caco-2 monolayers was assessed by measuring both TEER and FITC-dextran paracellular transport. Incubation in the presence of 5 ng ml−1 TNFα in the lower chamber (basolateral side of the Caco-2 monolayer) caused a significant decrease (30%, p < 0.05) in TEER, and a 1.7-fold increase in FITC-dextran paracellular transport (p < 0.05). These results confirm that under the current experimental conditions, TNFα caused an increase in the permeabilization of the Caco-2 cell monolayer. Addition of the AC (0.25 to 1 μM) to the upper chamber (apical side of the cell monolayer) caused a recovery of the monolayer TEER (Fig. 2A) and inhibition of the increased FITC-dextran transport (Fig. 2B) that depended on both the AC chemical structure and the AC concentration. | ||
| Fig. 2 Effects of 3-O-glucosides of cyanidin, delphinidin, malvidin, peonidin, and petunidin on TNFα-induced permeabilization of Caco-2 cell monolayers. Caco-2 cell monolayers were pre-incubated for 24 h with IFNγ (10 ng ml−1) to upregulate the TNFα receptor. Then, 0.25–1 μM of the 3-O-glucoside of cyanidin, delphinidin, malvidin, petunidin, or peonidin were added to the upper chamber and cells incubated for 30 min. TNFα (5 ng ml−1) was subsequently added to the lower chamber and cells incubated for an additional 6 h. Caco-2 cell monolayer permeability was evaluated by measuring: (A) TEER, and (B) FITC-dextran paracellular transport. Results are shown as mean ± SEM of 4–7 independent experiments. Values (A.U.: arbitrary units) were normalized to controls (1, dashed line). *Significantly different from control (no additions), **Significantly different from both control and TNFα-treated groups (p < 0.05, One-way ANOVA). | ||
Cyanidin-3-O-glucoside and delphinidin-3-O-glucoside both inhibited the TNFα-induced decrease in TEER in a concentration-dependent manner, attenuating the decrease at 0.25 μM, and providing total protection at 0.5 and 1 μM (Fig. 2A). On the contrary, malvidin- and peonidin-3-O-glucosides did not protect against the TNFα-induced TEER decrease within the range of concentrations tested (0.25–1 μM) (Fig. 2A). Petunidin-3-O-glucoside was protective only at the highest concentration tested (1 μM).
Both cyanidin-3-O-glucoside and delphinidin-3-O-glucoside inhibited the increase in FITC transport induced by TNFα at all concentrations tested (Fig. 2B). Petunidin-3-O-glucoside and peonidin-3-O-glucoside were both partially and fully protective at 0.5 and 1 μM concentrations, respectively, but showed no effect at 0.25 μM. Malvidin-3-O-glucoside had no protective effects at 0.25 and 0.5 μM, but did prevent TNFα-mediated increase in FITC-dextran transport at 1 μM concentration.
AC differentially protect Caco-2 cell monolayers from TNFα-induced NF-κB activation
The capacity of the AC glucosides to prevent TNFα-induced NF-κB activation was evaluated by measuring the activation (phosphorylation) of different steps in the pathway (IKK and p65 phosphorylation). After 5 min incubation, 5 ng ml−1 TNFα caused a 43 and 51% increase in IKKα (Ser176/180) and p65 (Ser536) phosphorylation, respectively, in differentiated Caco-2 cells. At 0.5 μM concentration, cyanidin and delphinidin 3-O-glucosides completely prevented TNFα-mediated increases in IKKα and p65 phosphorylation (Fig. 3A and B). Conversely, neither malvidin-3-O-glucoside, petunidin-3-O-glucoside nor peonidin-3-O-glucoside had the capacity to prevent IKKα and p65 phosphorylation. Downstream, NF-κB leads to the MLC kinase-mediated phosphorylation and activation of MLC. TNFα caused a 1.4 fold increase of MLC phosphorylation (Fig. 3C). While cyanidin, and delphinidin, prevented TNFα-induced MLC phosphorylation, peonidin and petunidin showed no protective capacity. MLC phosphorylation levels in cells incubated with TNFα and malvidin were not significantly different from controls and TNFα-treated cells.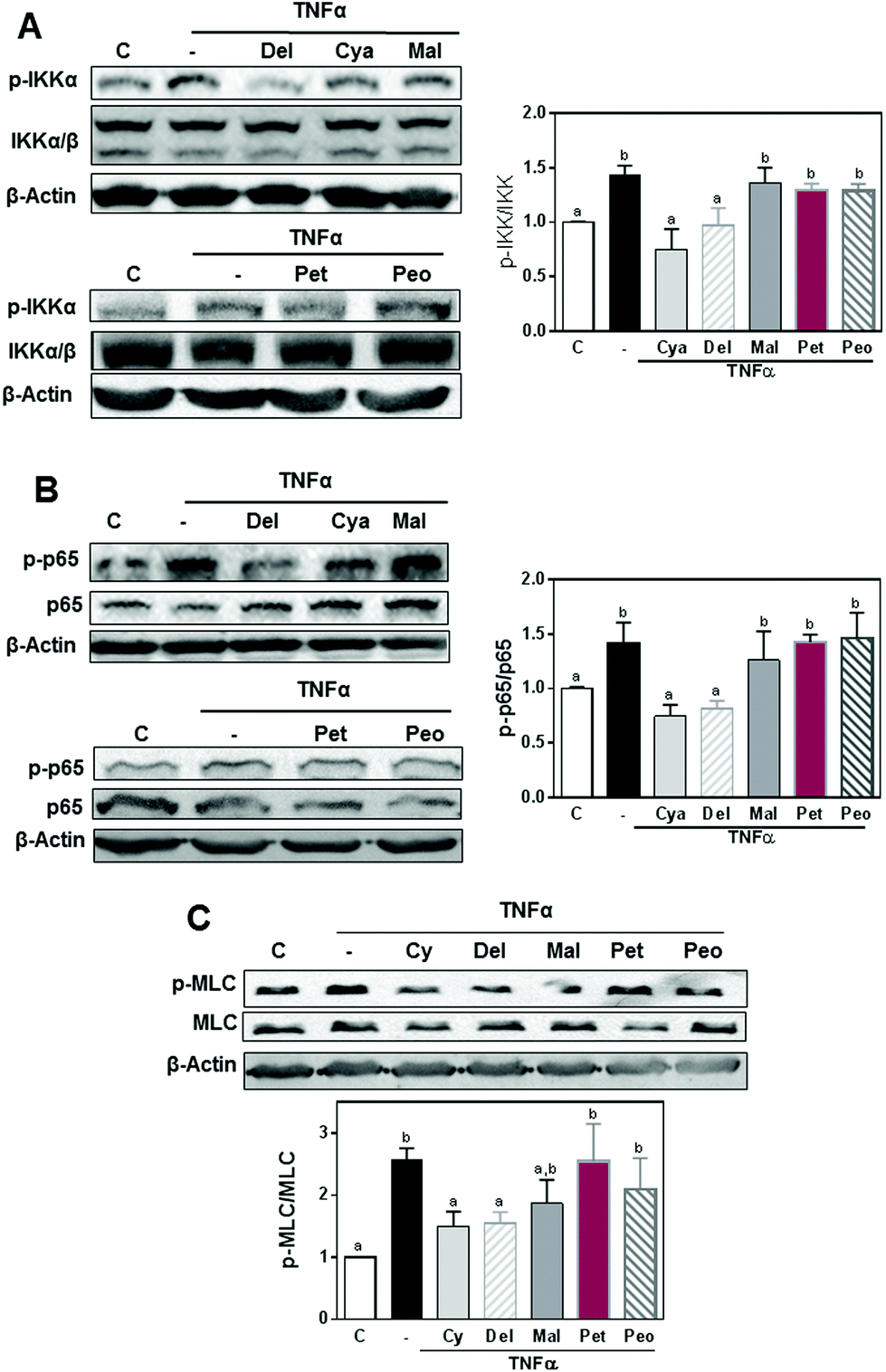 | ||
| Fig. 3 Effects of 3-O-glucosides of cyanidin, delphinidin, malvidin, peonidin, and petunidin on TNFα-induced NF-κB activation and MLC phosphorylation in differentiated Caco-2 cells. Caco-2 cells were incubated for 30 min at 37 °C in the absence or the presence of 0.5 μM of the 3-O-glucoside of cyanidin, delphinidin, or malvidin, and subsequently in the presence of TNFα (5 ng ml−1) for 5 min (for IKK and p65) or 6 h (for MLC phosphorylation). Western blots for (A) total and phosphorylated IKK (Ser178/180), and (B) total and phosphorylated p65 (Ser536), and (C) total and phosphorylated MLC (Thr18/Ser19). β-actin was measured as loading control. Western blot bands were quantified and results are shown as mean ± SEM of 3–5 independent experiments. Values having different superscripts are significantly different (p < 0.05, One-way ANOVA). | ||
ACRE differentially protect Caco-2 cell monolayers from TNFα-induced permeabilization
We next tested a series of ACRE, with variable AC content and composition, for their capacity to prevent TNFα-induced Caco-2 cell monolayer permeabilization. The content and identity of AC present in the studied ACRE was assessed by HPLC-UV and are shown in Table 1. Mass spectral data was used to confirm peaks observed at 520 nm, by matching the parent ion to the expected mass, as well as observing typical AC fragmentation (i.e. loss of sugar moiety).| ACRE | |||||||
|---|---|---|---|---|---|---|---|
| Anthocyanins | 1. Black chokeberry | 2. Black kernel rice | 3. Wild blueberry | 4. Bilberry | 5. Crowberry | 6. Blueberry | 7. Red grape |
| (μmol g−1) | |||||||
| Cyanidin-3-O-galactoside | 17.0 | 67 | 119 | 5.3 | |||
| Cyanidin-3-O-glucoside | 380 | 365 | 51 | 31.7 | |||
| Cyanidin-3-O-arabinoside | 162.6 | 55 | 38.3 | 142 | |||
| Cyanidin-3-O-rutinoside | 19.6 | ||||||
| Total cyanidin glycosides | 579 | 365 | 173 | 158 | 174 | 5.3 | |
| Delphinidin-3-O-galactoside | 78 | ||||||
| Delphinidin-3-O-glucoside | 14.5 | 98 | 213 | 92 | 14.5 | ||
| Delphinidin-3-O-arabinoside | 6.0 | 78 | 46 | ||||
| Delphinidin-3-O-rutinoside | 312 | ||||||
| Total delphinidin glycosides | 20.5 | 254 | 259 | 404 | 14.5 | ||
| Malvidin-3-O-galactoside | 17.8 | 32.7 | 30.7 | ||||
| Malvidin-3-O-glucoside | 24.2 | 73.6 | 310 | 145 | |||
| Malvidin-3-O-arabinoside | 10.4 | 21.5 | 39.8 | ||||
| Malvidin-3-O-(6′′-acetyl) glucoside | 8.7 | ||||||
| Malvidin-3-O-(6′′-acetyl) galactoside | 37 | ||||||
| Malvidin-3-O-(6′′-coumaryl) glucoside | 35 | ||||||
| Total malvidin glycosides | 55 | 128 | 381 | 225 | |||
| Peonidin-3-O-glucoside | 33 | 32 | 51 | 23.5 | |||
| Petunidin-3-O-galactoside | 10.8 | 38.7 | |||||
| Petunidin-3-O-glucoside | 12.2 | 66.6 | 114 | 32.8 | |||
| Petunidin-3-O-arabinoside | 4.1 | 25.3 | |||||
| Petunidin-3-O-(6′′-acetyl) glucoside | 10 | ||||||
| Total petunidin glycosides | 27.1 | 131 | 114 | 43 | |||
| Total anthocyanins | 579 | 398 | 103 | 718 | 963 | 578 | 311 |
Among the different extracts, AC content determined by HPLC-UV varied between 103 and 963 μmol g−1 extract. For the individual AC, their content ranged from 0 to 579, 404, 381, 51 and 131 μmol g−1 extract for the different glycosides of cyanidin, delphinidin, malvidin, peonidin, and petunidin, respectively.
The crowberry extract contained the highest AC content, 963 μmol g−1, and delivered a large diversity of individual AC (cyanidin, delphinidin, malvidin, peonidin, and petunidin glycosides). Addition of the crowberry extract (1–10 μg ml−1) to the apical side of the Caco-2 monolayer caused a concentration-dependent recovery of TNFα-induced TEER loss (Fig. 4A) and FITC-dextran transport increase (Fig. 4B). For both parameters of permeabilization, values were similar to control levels at the highest concentration assayed, i.e. 10 μg ml−1.
 | ||
| Fig. 4 Effects of the anthocyanins rich extracts (ACRE) on TNFα-induced permeabilization of Caco-2 cell monolayers. Caco-2 cell monolayers were pre-incubated for 24 h with IFNγ (10 ng ml−1) to upregulate the TNFα receptor. Then 1–10 μg ml−1 of crowberry extract (A,B) or 5 μg ml−1 of the each of the ACRE: (1) black chokeberry, (2) black kernel rice, (3) wild blueberry, (4) bilberry, (5) crowberry, (6) domesticated blueberry and (7) red grape (C,D) was added to the upper chamber and cells were incubated for 30 min. TNFα (5 ng ml−1) was subsequently added to the lower chamber and cells were incubated for an additional 6 h. Caco-2 cell monolayer permeability was evaluated by measuring: TEER (A,C), and FITC-dextran paracellular transport (B,D). Results are shown as mean ± SEM of 3–6 independent experiments. Values were normalized to controls (1, dashed line). *Significantly different from control (no additions), #Significantly different from the TNFα-treated group (p < 0.05, One-way ANOVA). | ||
The relative capacity of the remaining ACRE to inhibit TNFα-induced Caco-2 cell monolayer permeabilization was next investigated at 5 μg ml−1 concentration. The various ACRE differentially inhibited TNFα-induced alterations on TEER (Fig. 4C) and FITC-dextran paracellular transport (Fig. 4D). Black chokeberry, black kernel rice, and domesticated blueberry extracts most effectively inhibited TNFα-induced alterations on TEER, whereas black kernel rice, bilberry, and crowberry extracts most effectively inhibited TNFα-induced alterations on FITC-dextran paracellular transport.
To evaluate the contribution of the AC present in ACRE on the protective effects of the extracts on Caco-2 cell barrier integrity we analyzed the associations between TEER and the total and individual AC added to cell monolayers. Cyanidins and delphinidins defined the effects of the ACRE on TEER values as can be drawn from the positive correlations found between TEER and the sum of both AC and TEER values (r = 0.83, p < 0.02). Conversely, no significant correlations were found between TEER and the total determined AC, malvidin, or peonidin ACRE content. For FITC-dextran paracellular transport a significant negative correlation (r: 0.63, p < 0.005) was only found with petunidin ACRE content (data not shown).
Discussion
A permeable or “leaky” intestinal barrier is proposed as a major pathophysiological event underlying chronic inflammatory conditions of the gut, including those associated with lifestyle factors, such as consumption of western style diets.15 Dietary bioactives could contribute to a better gastrointestinal health by preserving intestinal permeability.7 In the present study, we investigated the capacity of a group of AC and ACRE to inhibit the permeabilization of Caco-2 cell monolayers. Cyanidins and delphinidins were selectively active compared to malvidins, peonidins and petunidins in the prevention of TNFα-induced loss of TEER and increase of paracellular transport. Such protective effects were confirmed using both pure AC and ACRE, and mechanistically associated with the inhibition NF-κB activation.The pure AC tested were the same as those present in the ACRE and accounted for most of the AC present in the extracts (>98%). Although the ACRE contained different glycosides, we only tested the action of pure glucosides in an attempt to compare the influence of the B-ring structures on monolayer permeability. The obtained results show that both the amount and the identity of AC define their protective effects on TNFα-induced permeabilization of Caco-2 cell monolayers. When permeability was assessed as TEER, regardless of the observed dose-dependent effects of both individual AC and ACRE, there was no correlation between ACRE capacity to inhibit TEER loss and the total AC content in the ACRE. This suggests that the observed protective actions are mainly dependent on the chemical structure and conformation of individual ACs. When pure 3-O-glucosides were evaluated the extent of protective effects on TEER decrease was cyanidin ∼ delphinidin ≫ petunidin > malvidin ∼ peonidin. Consistently, a significant correlation was observed between TEER and the content of cyanidin and delphinidin glucosides, but not of malvidin, peonidin or petunidin glucosides in the different ACRE studied. The most effective extracts in protecting Caco-2 monolayers from TNFα-induced TEER loss were those containing the highest cyanidin and delphinidin content (providing more than 0.11 μM AC in the cell culture medium), i.e. black chokeberry, black kernel rice, and domesticated blueberry extracts. Such effects of cyanidins were previously observed for an extract of AC-rich carrot containing only cyanidin glycosides where the extract attenuated the TEER decrease in Caco-2 monolayers when co-incubated with lipopolysaccharide-stimulated macrophages.23 In the present study, when cell monolayer permeability was evaluated as FITC-dextran transport, cyanidin and delphinidin glucosides were the most protective at the lowest concentration tested (0.25 μM); while at 1 μM all five pure AC were protective. This was consistent with results for the ACRE showing that all five pure AC were protective at the highest concentration tested. It is important to note that the concentrations of other flavonoids, e.g. (−)-epicatechin, and other phenolics, e.g. benzoic acids, and hydroxycinnamates, present in ACRE were relatively low (ESI Table 1†) and did not correlate with TEER or FITC-dextran transport values.
It has been described that TNFα affects intestinal permeability by modulating different signaling pathways,24 particularly NF-κB,25 affecting the functionality and structure of the tight junction.26 NF-κB increases the expression of MLCK which phosphorylates MLC at Ser19 leading to actin contraction and increased tight junction permeability.27 We observed that concurrent with their capacity to protect monolayer permeabilization, cyanidin and delphinidin, but not malvidin, peonidin or petunidin glucosides inhibited TNFα-induced NF-κB activation, and MLC phosphorylation. These results agree with a previous report showing that cyanidin inhibited TNFα-induced NF-κB activation and Nrf2 upregulation in Caco-2 cells.28 Consistently, the flavan-3-ol (−)-epicatechin, which shares the same chemical structure of the B ring as cyanidin, inhibited TNFα-induced Caco-2 cell monolayer permeabilization through the inhibition of NF-κB, attenuation of MLCK upregulation, and prevention of downregulation and redistribution of tight junction proteins.3
One important consideration when evaluating AC bioactivity in cell cultures is the possibility that their interactions with cell components may be dictated by their stability in the culture media, interaction with cell membranes, incorporation into cells, and/or transport across cells forming monolayers.22,29–31 Although, we did not evaluate the stability of the AC or their presence inside the cells, it was recently demonstrated that under similar experimental conditions the assayed AC are mainly incorporated into the Caco-2 cells.31 Not only can AC be transported across Caco-2 monolayers, they can also affect the transport of other flavonoids.32 Furthermore, other structural characteristics not evaluated in this study, i.e. AC acylation33 and glycosylation30 may affect AC uptake, absorption and bioactivities.
Based on the obtained results and the crosstalk between NF-κB and cell redox conditions, it could be speculated that AC affect redox signaling by preventing NADPH-oxidase activation.2 We previously described that NADPH oxidase activation is a major player in TNFα- and bile acid-mediated NF-κB activation and subsequent Caco-2 monolayer permeabilization.3,4,34 The capacity of flavonoids to inhibit NADPH oxidase is attributed mostly to substitutions in the flavonoid B ring.35,36 In this regard, the chemical characteristics of cyanidins and delphinidins that may be responsible for their prevention of NADPH-oxidase activation could be the presence of free hydroxyl groups (or absence of bulky methoxy groups) on the B ring and/or the positioning of ring B with regard to C.
In summary, this paper shows that cyanidin and delphinidin AC are selectively and highly effective in protecting Caco-2 monolayers against TNFα-induced NF-κB activation and subsequent barrier permeabilization. Their action was observed within a range of concentrations that would be realistically expected in the gut after consumption of AC containing foods. These findings support the concept that consumption of fruit, berries, vegetables, derived foods, or supplements containing AC, especially cyanidins and delphinidins, could afford beneficial health effects at the level of the gastrointestinal tract. These effects would be relevant not only in disease states, but also in conditions of chronic gut inflammation associated with environmental factors including diet and excess body weight.
Acknowledgements
This work was supported by NIFA-USDA (CA-D*-XXX-7244-H) and a gift from Pharmanex Research, NSE Products Inc., Provo, UT, USA. PIO is correspondent researcher from CONICET, Argentina.References
- C. G. Fraga, M. Galleano, S. V. Verstraeten and P. I. Oteiza, Basic biochemical mechanisms behind the health benefits of polyphenols, Mol. Aspects Med., 2010, 31, 435–445 CrossRef CAS PubMed.
- C. G. Fraga and P. I. Oteiza, Dietary flavonoids: Role of (-)-epicatechin and related procyanidins in cell signaling, Free Radicals Biol. Med., 2011, 51, 813–823 CrossRef CAS PubMed.
- T. C. Contreras, E. Ricciardi, E. Cremonini and P. I. Oteiza, (-)-Epicatechin in the prevention of tumor necrosis alpha-induced loss of Caco-2 cell barrier integrity, Arch. Biochem. Biophys., 2015, 573, 84–91 CrossRef CAS PubMed.
- M. Da Silva, G. K. Jaggers, S. V. Verstraeten, A. G. Erlejman, C. G. Fraga and P. I. Oteiza, Large procyanidins prevent bile-acid-induced oxidant production and membrane-initiated ERK1/2, p38, and Akt activation in Caco-2 cells, Free Radicals Biol. Med., 2012, 52, 151–159 CrossRef CAS PubMed.
- A. G. Erlejman, C. G. Fraga and P. I. Oteiza, Procyanidins protect Caco-2 cells from bile acid- and oxidant-induced damage, Free Radicals Biol. Med., 2006, 41, 1247–1256 CrossRef CAS PubMed.
- A. G. Erlejman, G. Jaggers, C. G. Fraga and P. I. Oteiza, TNFalpha-induced NF-kappaB activation and cell oxidant production are modulated by hexameric procyanidins in Caco-2 cells, Arch. Biochem. Biophys., 2008, 476, 186–195 CrossRef CAS PubMed.
- U. Wenzel, Flavonoids as drugs at the small intestinal level, Curr. Opin. Pharmacol., 2013, 13, 864–868 CrossRef CAS PubMed.
- A. Crozier, D. Del Rio and M. N. Clifford, Bioavailability of dietary flavonoids and phenolic compounds, Mol. Aspects Med., 2010, 31, 446–467 CrossRef CAS PubMed.
- J. Fang, Classification of fruits based on anthocyanin types and relevance to their health effects, Nutrition, 2015, 31, 1301–1306 CrossRef CAS PubMed.
- A. Castañeda-Ovando, M. Pacheco-Hernández, M. Páez-Hernández, J. Rodríguez and C. Galán-Vidal, Chemical studies of anthocyanins: A review, Food Chem., 2009, 113, 859–871 CrossRef.
- J. M. Kong, L. S. Chia, N. K. Goh, T. F. Chia and R. Brouillard, Analysis and biological activities of anthocyanins, Phytochemistry, 2003, 64, 923–933 CrossRef CAS PubMed.
- T. Tsuda, Dietary anthocyanin-rich plants: biochemical basis and recent progress in health benefits studies, Mol. Nutr. Food Res., 2012, 56, 159–170 CAS.
- A. Fasano and T. Shea-Donohue, Mechanisms of disease: the role of intestinal barrier function in the pathogenesis of gastrointestinal autoimmune diseases, Nat. Clin. Pract. Gastroenterol. Hepatol., 2005, 2, 416–422 CrossRef CAS PubMed.
- J. M. Wells, R. J. Brummer, M. Derrien, T. T. MacDonald, F. Troost, P. D. Cani, V. Theodorou, J. Dekker, A. Meheust, W. M. de Vos, A. Mercenier, A. Nauta and C. L. Garcia-Rodenas, Homeostasis of the gut barrier and potential biomarkers, Am. J. Physiol.: Gastrointest. Liver Physiol., 2017, 312, G171–G193 CrossRef PubMed.
- S. C. Bischoff, G. Barbara, W. Buurman, T. Ockhuizen, J. D. Schulzke, M. Serino, H. Tilg, A. Watson and J. M. Wells, Intestinal permeability–a new target for disease prevention and therapy, BMC Gastroenterol., 2014, 14, 189 CrossRef PubMed.
- J. Konig, J. Wells, P. D. Cani, C. L. Garcia-Rodenas, T. MacDonald, A. Mercenier, J. Whyte, F. Troost and R. J. Brummer, Human intestinal barrier function in health and disease, Clin. Transl. Gastroenterol., 2016, 7, e196 CrossRef PubMed.
- R. Gonzalez-Barrio, G. Borges, W. Mullen and A. Crozier, Bioavailability of anthocyanins and ellagitannins following consumption of raspberries by healthy humans and subjects with an ileostomy, J. Agric. Food Chem., 2010, 58, 3933–3939 CrossRef PubMed.
- I. A. Ludwig, P. Mena, L. Calani, G. Borges, G. Pereira-Caro, L. Bresciani, D. Del Rio, M. E. Lean and A. Crozier, New insights into the bioavailability of red raspberry anthocyanins and ellagitannins, Free Radicals Biol. Med., 2015, 89, 758–769 CrossRef CAS PubMed.
- U. Buckert and N. L. Allinger, Molecular mechanics, ACS Monograph 177, 1982 Search PubMed.
- A. Fischer, M. Gluth, U. F. Pape, B. Wiedenmann, F. Theuring and D. C. Baumgart, Adalimumab prevents barrier dysfunction and antagonizes distinct effects of TNF-alpha on tight junction proteins and signaling pathways in intestinal epithelial cells, Am. J. Physiol.: Gastrointest. Liver Physiol., 2013, 304, G970–G979 CrossRef CAS PubMed.
- R. M. de Ferrars, C. Czank, S. Saha, P. W. Needs, Q. Zhang, K. S. Raheem, N. P. Botting, P. A. Kroon and C. D. Kay, Methods for isolating, identifying, and quantifying anthocyanin metabolites in clinical samples, Anal. Chem., 2014, 86, 10052–10058 CrossRef CAS PubMed.
- C. D. Kay, P. A. Kroon and A. Cassidy, The bioactivity of dietary anthocyanins is likely to be mediated by their degradation products, Mol. Nutr. Food Res., 2009, 53, S92–S101 Search PubMed.
- A. Olejnik, K. Kowalska, M. Kidon, J. Czapski, J. Rychlik, M. Olkowicz and R. Dembczynski, Purple carrot anthocyanins suppress lipopolysaccharide-induced inflammation in the co-culture of intestinal Caco-2 and macrophage RAW264.7 cells, Food Funct., 2016, 7, 557–564 CAS.
- D. Ye, I. Ma and T. Y. Ma, Molecular mechanism of tumor necrosis factor-alpha modulation of intestinal epithelial tight junction barrier, Am. J. Physiol.: Gastrointest. Liver Physiol., 2006, 290, G496–G504 CrossRef CAS PubMed.
- T. Y. Ma, G. K. Iwamoto, N. T. Hoa, V. Akotia, A. Pedram, M. A. Boivin and H. M. Said, TNF-alpha-induced increase in intestinal epithelial tight junction permeability requires, NF-kappa B activation, Am. J. Physiol.: Gastrointest. Liver Physiol., 2004, 286, G367–G376 CrossRef CAS PubMed.
- T. Y. Ma, M. A. Boivin, D. Ye, A. Pedram and H. M. Said, Mechanism of TNF-{alpha} modulation of Caco-2 intestinal epithelial tight junction barrier: role of myosin light-chain kinase protein expression, Am. J. Physiol.: Gastrointest. Liver Physiol., 2005, 288, G422–G430 CrossRef CAS PubMed.
- K. E. Cunningham and J. R. Turner, Myosin light chain kinase: pulling the strings of epithelial tight junction function, Ann. N. Y. Acad. Sci., 2012, 1258, 34–42 CrossRef CAS PubMed.
- D. Ferrari, A. Speciale, M. Cristani, D. Fratantonio, M. S. Molonia, G. Ranaldi, A. Saija and F. Cimino, Cyanidin-3-O-glucoside inhibits NF-kB signalling in intestinal epithelial cells exposed to TNF-alpha and exerts protective effects via Nrf2 pathway activation, Toxicol. Lett., 2016, 264, 51–58 CrossRef CAS PubMed.
- S. Kamiloglu, E. Capanoglu, C. Grootaert and J. Van Camp, Anthocyanin absorption and metabolism by human intestinal Caco-2 cells-a review, Int. J. Mol. Sci., 2015, 16, 21555–21574 CrossRef CAS PubMed.
- C. D. Kay, G. Pereira-Caro, I. A. Ludwig, M. N. Clifford and A. Crozier, Anthocyanins and flavanones are more bioavailable than previously perceived: a review of recent evidence, Annu. Rev. Food Sci. Technol., 2017, 8, 155–180 CrossRef CAS PubMed.
- S. Kuntz, S. Rudloff, H. Asseburg, C. Borsch, B. Frohling, F. Unger, S. Dold, B. Spengler, A. Rompp and C. Kunz, Uptake and bioavailability of anthocyanins and phenolic acids from grape/blueberry juice and smoothie in vitro and in vivo, Br. J. Nutr., 2015, 113, 1044–1055 CrossRef CAS PubMed.
- B. W. Redan, G. P. Albaugh, C. S. Charron, J. A. Novotny and M. G. Ferruzzi, Adaptation in Caco-2 human intestinal cell differentiation and phenolic transport with chronic exposure to blackberry (Rubus sp.) extract, J. Agric. Food Chem., 2017, 65, 2694–2701 CrossRef CAS PubMed.
- J. A. Novotny, B. A. Clevidence and A. C. Kurilich, Anthocyanin kinetics are dependent on anthocyanin structure, Br. J. Nutr., 2012, 107, 504–509 CrossRef CAS PubMed.
- Y. Araki, T. Katoh, A. Ogawa, S. Bamba, A. Andoh, S. Koyama, Y. Fujiyama and T. Bamba, Bile acid modulates transepithelial permeability via the generation of reactive oxygen species in the Caco-2 cell line, Free Radicals Biol. Med., 2005, 39, 769–780 CrossRef CAS PubMed.
- Y. Steffen, C. Gruber, T. Schewe and H. Sies, Mono-O-methylated flavanols and other flavonoids as inhibitors of endothelial NADPH oxidase, Arch. Biochem. Biophys., 2008, 469, 209–219 CrossRef CAS PubMed.
- Y. Steffen, T. Schewe and H. Sies, (-)-Epicatechin elevates nitric oxide in endothelial cells via inhibition of NADPH oxidase, Biochem. Biophys. Res. Commun., 2007, 359, 828–833 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c7fo00625j |
| This journal is © The Royal Society of Chemistry 2017 |