
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Protein conformation in pure and hydrated deep eutectic solvents†
A.
Sanchez-Fernandez
 *ab,
K. J.
Edler
a,
T.
Arnold
c,
D.
Alba Venero
d and
A. J.
Jackson
be
*ab,
K. J.
Edler
a,
T.
Arnold
c,
D.
Alba Venero
d and
A. J.
Jackson
be
aDepartment of Chemistry, University of Bath, Claverton Down, Bath, BA2 7AY, UK. E-mail: a.sanchez.fernandez@bath.ac.uk
bEuropean Spallation Source, 221 00, Lund, Sweden
cDiamond Light Source, Harwell Campus, Didcot, OX11 0DE, UK
dISIS Spallation Neutron Source, Harwell Campus, Didcot, OX11 OQX, UK
eDepartment of Physical Chemistry, Lund University, SE-221 00, Lund, Sweden
First published on 14th March 2017
Abstract
Deep eutectic solvents (DES) have recently been postulated as possible environments where protein structure may be preserved in the absence of water. Here we present our results towards understanding protein conformation in choline chloride-based DES and mixtures with water. Lysozyme and bovine serum albumin have been investigated by means of circular dichroism and small-angle neutron scattering.
The investigation of proteins and enzymes in the absence of water has experienced a recent upsurge with the emergence of neoteric solvents, mainly deep eutectic solvents.1 The green character of the latter has attracted attention for possible applications in biochemistry and biocatalysis as an alternative to organic solvents used in traditional processes.2 Deep eutectic solvents have also been postulated as a matrix where cell metabolism may occur in extreme environments (e.g. cryogenic temperatures or total absence of water).3 Therefore understanding the behaviour of proteins in these solvents not only provides a new alternative to aqueous solvents, but may also help to elucidate information on protein activity in extreme environments.
Deep eutectic solvents are formed through the complexation of an ionic species (e.g. choline chloride) and a hydrogen bond donor (e.g. glycerol).1 The interaction between these compounds produces a large depression of the freezing point which allows the formation of a stable solvent at room temperature.4 DES have been demonstrated to promote amphiphile self-assembly of surfactants and phospholipids, both essential for the formation of biological membranes.5–9 Protein stability has been investigated in various DES,10–13 and enzymatic activity has been explored as an alternative to water for enzyme-catalysed reactions.14–16 Bacterial preservation has also been demonstrated in DES.17
Unlike in organic solvents, where it has been shown that enzymatic activity is dramatically reduced,18 DES and DES/water mixtures have been shown to partially preserve such activity.12 The structure of proteins in these media has been previously studied by means of circular dichroism (CD).10 Although such studies provide an insight on the secondary structure of proteins, information on the overall conformation of the proteins in solution is yet missing and not fully understood. In this communication we present the first investigation on protein conformation and structure in pure and hydrated DES. The extent to which these proteins remain folded in the total or partial absence of water is explored here for two model proteins: Hen egg white lysozyme and bovine serum albumin (BSA). Circular dichroism was used to study changes in the secondary and tertiary structure of the proteins, whilst protein conformation in solution was investigated by means of small-angle neutron scattering (SANS).‡ Protein characteristics have been investigated and compared in terms of protein environment (pure DES, DES/water mixtures and buffer) and thermal stability.
BSA structure has been investigated in choline chloride:glycerol, and choline chloride:glycerol/water mixtures (two solvents: 75 wt% and 50 wt% of DES) and buffer (phosphate buffered saline, 0.01 M, pH = 7.4).§ Further investigations on conformational changes of BSA with temperature are also presented here. Lysozyme has been studied in choline chloride:urea, choline chloride:glycerol, choline chloride:urea/water mixtures and buffer (same as above). The systems investigated in the present study are summarised in Table 1.
| Solvent | Protein |
|---|---|
| Choline chloride:glycerol | Bovine serum albumin |
| Choline chloride:glycerol/water | |
| Phosphate buffered saline | |
| Choline chloride:glycerol | Lysozyme |
| Choline chloride:urea | |
| Choline chloride:glycerol/water | |
| Phosphate buffered saline | |
The CD spectra for BSA in these solvents is shown in Fig. 1. The far-UV CD region is sensitive to changes in the secondary structure of the protein while the near-UV CD region is sensitive to the tertiary structure.19 Our measurements show that the secondary structure of BSA does not vary between solvent environments.
 | ||
| Fig. 1 CD spectra of BSA: (a) far-UV region and (c) near-UV region of BSA in pure choline chloride:glycerol, choline chloride:glycerol/water mixtures (50 and 75 wt% of DES) and phosphate buffer; (b) far-UV region and (d) near-UV region of BSA in pure choline chloride:glycerol at different temperatures. | ||
The shape of the spectra in the far-UV region indicates the presence of an alpha helix. The near-UV CD spectra of BSA in buffer and in the DES/water mixtures appear to be similar, indicating minimal differences between the tertiary structure of the protein in these solvents. However, the spectrum of BSA in pure choline chloride:glycerol is considerably different from those solvents that contain water, indicating a difference in the tertiary structure of the protein. Whilst CD is also sensitive to small changes in the tertiary structure, it is not possible to extract a detailed model from the data. However this technique can be applied to follow changes in the protein folding while varying the protein environment or temperature.19–21
On heating the BSA samples to 80 °C with equilibration for at least 1 hour before measurement, the CD spectra show a clear decrease in the signal and therefore in the degree of secondary structure of the protein. Also the near-UV region spectra showed a significant loss of intensity and flattening of the signal, indicating unfolding of the protein (Fig. 1c and d). These samples were afterwards re-equilibrated at room temperature and remeasured demonstrating that the change was not reversible and therefore indicative of denaturation.22
Lysozyme secondary structure gives a CD signal arising from a combination of alpha helices and beta sheets in buffer solution.23 The secondary structure of lysozyme in DES has been previously investigated by Esquembre et al. and Xin et al. using CD.10,12 Our results were found to be in agreement with those previously published, showing a minimal change of the alpha helices and beta sheets in choline chloride:glycerol, and choline chloride:urea.10 Lysozyme has also been studied using CD in a choline chloride:trehalose:water buffer, showing an increase in the thermostability of the protein.12
Small-angle neutron scattering data were analysed following a well-established procedure for proteins in solution, using the Indirect Fourier Transformation approach.24,25 Examining the data using this technique provides a good way to follow conformational changes in the protein. Guinier analysis was used to obtain the extrapolated intensity at zero scattering angle, I(0), and the radius of gyration, Rg, of the protein (see ESI†). These parameters were subsequently used to plot the data and use a normalised Kratky analysis.26,27 This dimensionless plot provides an assessment of protein conformation, showing peak maxima at √3 for globular particles regardless of concentration, composition and size. Deviations from this condition suggest asymmetry or flexibility in the protein structure.
The pair distance distribution function, p(r), shows that the maximum dimension of the scatterer (Dmax), and therefore the protein conformation, vary between pure DES and DES/water mixtures. In the case of BSA in pure D-choline chloride:D-glycerol, Dmax was found to be larger than in buffer but significantly shorter that the denatured protein (131 ± 4 Å vs. 314 ± 12 Å) (Fig. 2b), indicating a partially folded structure with a Rg of 42.2 ± 0.4 Å (see ESI†). The Dmax of BSA was fitted to 98 ± 3 Å in 75/25 DES/water mixture and 94 ± 5 Å in 50/50 DES/water, whilst they showed a Rg of 28.4 ± 0.2 and 28.2 ± 0.3 Å, respectively. These values are close to those seen in the buffered solution where Dmax = 93 ± 3 Å and Rg = 28.1 ± 0.3 Å.
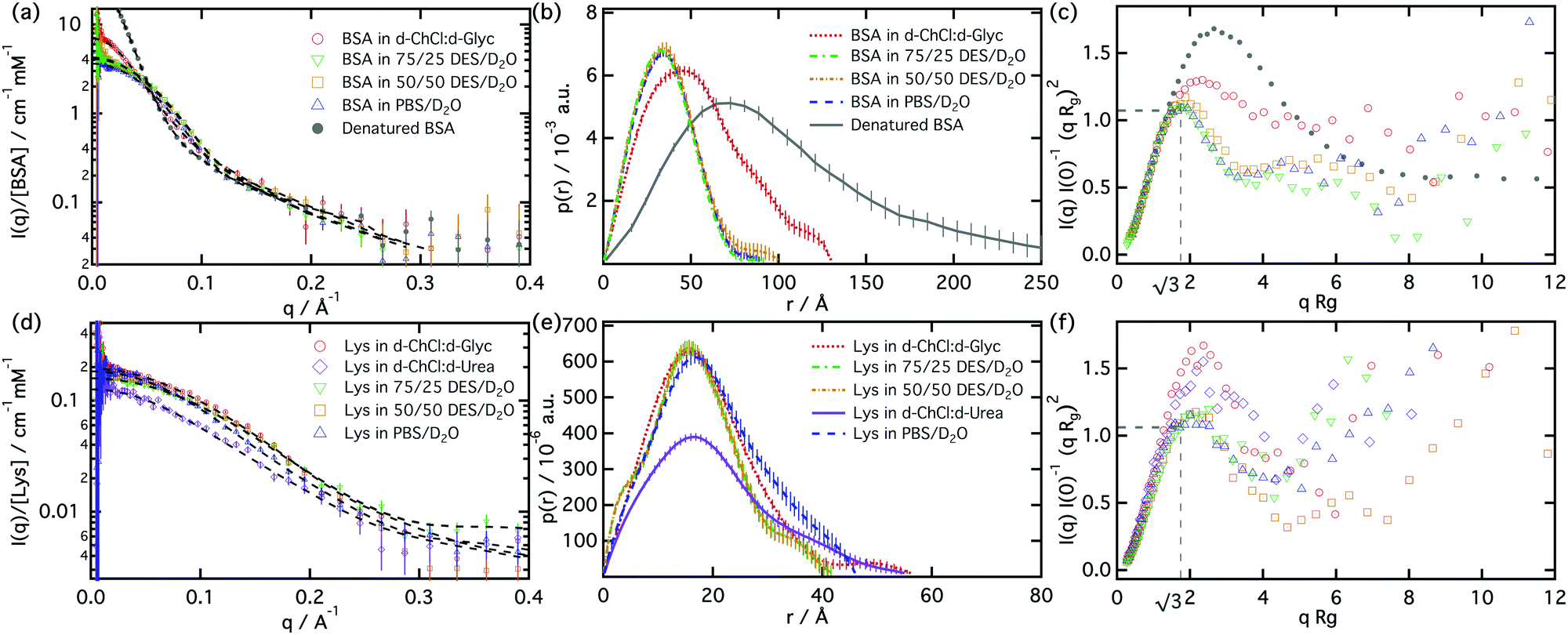 | ||
| Fig. 2 SANS data from (a) BSA and (d) lysozyme in different solvents. The black dashed lines correspond to the best fits using the Inverse Fourier Transformation approach. Pair distance distribution function of (b) BSA and (e) lysozyme in different solvents. Normalised Kratky plot of (c) BSA and (f) lysozyme in different solvents. The grey lines correspond to the position of the maximum, which is indicative the presence of globular structures in solution. | ||
The addition of water appears to allow a more efficient folding into a globular shape, with the normalised Kratky plot showing how the protein changes from globular in DES/water systems to a less symmetric conformation in pure choline chloride:glycerol (Fig. 2c). This suggests that this difference is related to changes in the local environment of the functional groups that promote the folding of the protein. As has been previously demonstrated, despite the presence of the hydrophobic effect and specific ion–ion interactions on surfactant–DES systems,6,7 the solvation ability of DES differs from water and therefore the mechanism of solvation still remains unclear.
The hydrated systems however do not follow the same behaviour. The fits indicate that BSA conformation is maintained, even at high concentration of DES acting as a co-solvent. This observation would support the hypothesis which argues that DES structure is maintained even at high water concentration, with discrete strongly bonded DES clusters remaining in solution, which do not directly interact with the protein.28 Thus, water could form a shell protecting the protein and providing a favourable environment for the protein.
In agreement with our CD results, BSA was found to undergo hot denaturation above 80 °C, irreversibly losing its conformation. Cold denaturation was not observed, with the protein remaining folded after cooling down to −80 °C (see ESI†).
In the case of lysozyme, choline chloride:urea and choline chloride:glycerol DES were tested as solvents for the protein. As in the BSA/DES systems, lysozyme was found to remain partially folded in both of the pure DES (Fig. 2d). The scattering from the protein in choline chloride:glycerol indicates that the protein folding is similar to that in choline chloride:urea (Dmax = 57.3 ± 1.1 Å, Rg = 17.2 ± 0.8 Å and 58.0 ± 0.8 Å, Rg = 17.8 ± 1.0 Å, respectively) (Fig. 2e). This is surprising given that urea has been widely used as a natural agent to cause denaturation of proteins,11 and yet here lysozyme retains its structure in a solvent with a high content of urea. Lysozyme in mixtures of choline chloride:glycerol and water showed a similar structure. The protein seems to retain its structure in these mixtures, with small differences in conformation compared with that seen in the pure buffer. However these differences in the structure can be appreciated in the normalised Kratky plot (Fig. 2f), where the peak diverges from the position of the completely folded lysozyme structure in buffer.
Esquembre et al. presented investigations on lysozyme activity in pure and hydrated solvents.10 These results showed a decrease of the protein activity with increasing concentration of DES in the solvent. Although the conformation of lysozyme does not vary between hydrated DES and buffer, the activity of the protein decreases in the presence of the DES. Similar observations have been also reported for lysozyme in different ionic liquids, where the structure of the protein is maintained but the activity is reduced.29,30 We believe that although the protein structure is not affected, the high ionic strength and slower diffusion, arising from higher viscosity of the DES solution, hinder the interaction of the interaction sites with the bacteria used in the assay.
Conclusions
DES and mixtures of those with water have been investigated as media to support protein folding. Circular dichroism has shown that the secondary structure of the proteins examined is minimally affected as the solvent varies from phosphate buffer to pure DES. Near-UV measurements have provided information regarding the change of the tertiary structure of the protein. These results showed that the protein folding was similar in buffer and DES/water mixtures (75/25 and 50/50). However a considerable change was found for the protein solubilised in pure DES. BSA was found to irreversibly denature once heated up to 80 °C, showing that the DES provided no improvement in thermostability for this protein in the present systems. As shown by CD, this transition affects both the secondary and tertiary structure of the protein.Small-angle neutron scattering studies provided information about the structure of two proteins in DES. The structure of BSA and lysozyme in pure DES was found to be partially folded, as can be seen through the pair distance-distribution function. Our fits situate the structure of BSA in pure choline chloride:glycerol between the optimal conformation in buffer and the denatured elongated structure. In the case of glycerol and urea, these different hydrogen bond donors were found to promote a similar partially-folded conformation of lysozyme in the pure DES.
Interestingly the proteins in DES/water mixtures showed minimal differences from the proteins in buffer, despite the concentrations of DES being rather high (50 and 75 wt% of DES). Further investigations will be needed to determine whether this high concentration of DES is acting as a co-solvent, and therefore retaining its solvent structure regardless the presence of water, or the components are found dissociated in such media.
Based on our results on protein conformation in DES, we hypothesize the conformation–activity relationship as follows: protein conformation is dependent on the local environments of the amino acids that promote protein folding – these amino acids may also be part of the active site of the protein which is responsible for its enzymatic activity. The presence of pure DES, replacing water around the amino acids in the active site therefore promotes partially folded conformations and reduces the activity via a combination of specific binding and reduction of solvophobic effects. In the case of hydrated DES, water may form a shell around those active sites, allowing the folding of the protein as in buffer. However, the presence of DES molecules in the system still decreases the activity by limiting the diffusion of components and limiting the binding between protein and the bacteria used in the standard lysozyme assay.
Overall, the results presented here provide a new insight into protein folding in deep eutectic solvents and their mixtures with water. The decrease of enzyme activity observed by others in pure DES12,14,31,32 can likely be explained through the more unfolded conformation seen in pure DES compared to buffer solutions. However, the addition of water allows the protein to adopt a more normal conformation and could restore enzyme activity.
Acknowledgements
This work was performed with support from The Crafoord Foundation (Grant #20140788), the European Spallation Source, and Bath University Alumni Funding. The authors thank the Physical Chemistry Department, University of Lund, for access to equipment and to the ISIS Facility of the Science and Technology Facilities Council, UK, for the neutron beamtime (RB1520285).Notes and references
- E. L. Smith, A. P. Abbott and K. S. Ryder, Chem. Rev., 2014, 114, 11060–11082 CrossRef CAS PubMed.
- Y. Dai, J. van Spronsen, G. J. Witkamp, R. Verpoorte and Y. H. Choi, Anal. Chim. Acta, 2013, 766, 61–68 CrossRef CAS PubMed.
- Y. H. Choi, J. van Spronsen, Y. Dai, M. Verberne, F. Hollmann, I. W. C. E. Arends, G.-J. Witkamp and R. Verpoorte, Plant Physiol., 2011, 156, 1701–1705 CrossRef CAS PubMed.
- O. S. Hammond, D. T. Bowron and K. J. Edler, Green Chem., 2016, 18, 2736–2744 RSC.
- T. Arnold, A. J. Jackson, A. Sanchez-Fernandez, D. Magnone, A. E. Terry and K. J. Edler, Langmuir, 2015, 31, 12894–12902 CrossRef CAS PubMed.
- A. Sanchez-Fernandez, K. J. Edler, T. Arnold, R. K. Heenan, L. Porcar, N. J. Terrill, A. E. Terry and A. J. Jackson, Phys. Chem. Chem. Phys., 2016, 18, 14063–14073 RSC.
- A. Sanchez-Fernandez, T. Arnold, A. J. Jackson, S. L. Fussell, R. K. Heenan, R. A. Campbell and K. J. Edler, Phys. Chem. Chem. Phys., 2016, 18, 33240–33249 RSC.
- M. C. Gutiérrez, M. L. Ferrer, C. R. Mateo and F. del Monte, Langmuir, 2009, 25, 5509–5515 CrossRef PubMed.
- S. J. Bryant, R. Atkin and G. G. Warr, Soft Matter, 2016, 12, 1645–1648 RSC.
- R. Esquembre, J. M. Sanz, J. G. Wall, F. del Monte, C. R. Mateo and M. L. Ferrer, Phys. Chem. Chem. Phys., 2013, 15, 11248–11256 RSC.
- H. Monhemi, M. R. Housaindokht, A. A. Moosavi-Movahedi and M. R. Bozorgmehr, Phys. Chem. Chem. Phys., 2014, 16, 14882–14893 RSC.
- R. P. Xin, S. J. Qi, C. X. Zeng, F. I. Khan, B. Yang and Y. H. Wang, Food Chem., 2017, 217, 560–567 CrossRef CAS PubMed.
- J. Parnica and M. Antalik, J. Mol. Liq., 2014, 197, 23–26 CrossRef CAS.
- A. R. Harifi-Mood, R. Ghobadi and A. Divsalar, Int. J. Biol. Macromol., 2017, 95, 115–120 CrossRef CAS PubMed.
- A. A. Papadopoulou, E. Efstathiadou, M. Patila, A. C. Polydera and H. Stamatis, Ind. Eng. Chem. Res., 2016, 55, 5145–5151 CrossRef CAS.
- B.-P. Wu, Q. Wen, H. Xu and Z. Yang, J. Mol. Catal. B: Enzym., 2014, 101, 101–107 CrossRef CAS.
- M. C. Gutiérrez, M. L. Ferrer, L. Yuste, F. Rojo and F. del Monte, Angew. Chem., Int. Ed., 2010, 49, 2158–2162 CrossRef PubMed.
- A. M. Klibanov, Trends Biotechnol., 1997, 15, 97–101 CrossRef CAS PubMed.
- O. K. Gasymov, A. R. Abduragimov and B. J. Glasgow, J. Phys. Chem. B, 2014, 118, 986–995 CrossRef CAS PubMed.
- S. M. Kelly, T. J. Jess and N. C. Price, Biochim. Biophys. Acta, 2005, 1751, 119–139 CrossRef CAS PubMed.
- C. Sun, J. Yang, X. Wu, X. Huang, F. Wang and S. Liu, Biophys. J., 2005, 88, 3518–3524 CrossRef CAS PubMed.
- Y. Moriyama, E. Watanabe, K. Kobayashi, H. Harano, E. Inui and K. Takeda, J. Phys. Chem. B, 2008, 112, 16585–16589 CrossRef CAS PubMed.
- F. Tanaka, L. S. Forster, P. K. Pal and J. A. Rupley, J. Biol. Chem., 1975, 250, 6977–6982 CAS.
- O. Glatter, J. Appl. Crystallogr., 1979, 12, 166–175 CrossRef CAS.
- D. A. Jacques, J. M. Guss, D. I. Svergun and J. Trewhella, Acta Crystallogr., Sect. D: Biol. Crystallogr., 2012, 68, 620–626 CrossRef CAS PubMed.
- D. Durand, C. Vivès, D. Cannella, J. Pérez, E. Pebay-Peyroula, P. Vachette and F. Fieschi, J. Struct. Biol., 2010, 169, 45–53 CrossRef CAS PubMed.
- R. P. Rambo and J. A. Tainer, Biopolymers, 2011, 95, 559–571 CrossRef CAS PubMed.
- O. S. Hammond, D. T. Bowron and K. J. Edler, 2017, submitted manuscript.
- K. D. Weaver, R. M. Vrikkis, M. P. Van Vorst, J. Trullinger, R. Vijayaraghavan, D. M. Foureau, I. H. McKillop, D. R. MacFarlane, J. K. Krueger and G. D. Elliott, Phys. Chem. Chem. Phys., 2012, 14, 790–801 RSC.
- Y. Yoshimura, T. Takekiyo and T. Mori, Chem. Phys. Lett., 2016, 664, 44–49 CrossRef CAS.
- B. Kleiner, P. Fleischer and U. Schörken, Process Biochem., 2016, 51, 1808–1816 CrossRef CAS.
- J. T. Gorke, F. Srienc and R. J. Kazlauskas, Chem. Commun., 2008, 1235–1237, 10.1039/b716317g.
- R. K. Heenan, S. E. Rogers, D. Turner, A. E. Terry, J. Treadgold and S. M. King, Neutron News, 2011, 22, 3 CrossRef.
- O. Glatter, J. Appl. Crystallogr., 1977, 10, 415–421 CrossRef.
- M. V. Petoukhov, D. Franke, A. V. Shkumatov, G. Tria, A. G. Kikhney, M. Gajda, C. Gorba, H. D. T. Mertens, P. V. Konarev and D. I. Svergun, J. Appl. Crystallogr., 2012, 45, 342–350 CAS.
Footnotes |
| † Electronic supplementary information (ESI) available: Details regarding sample preparation, techniques and data analysis. Results of Guinier analysis of the scattering data and log–log plots of scattering data. See DOI: 10.1039/c7cp00459a |
| ‡ CD measurements were taken using a Jasco J-815 spectrometer. Data were normalised to protein concentration and molecular weight. SANS measurements were performed on Sans2d at the ISIS Facility, STFC, (UK).33 Indirect Fourier Transformation analysis of SANS data was done using then GNOM software from the ATSAS package.34,35 Technical details about measurements and data analysis are included in the ESI.† |
§ Choline chloride![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :glycerol (1:2 mole ratio) and choline chloride:urea (1:2 mole ratio) were prepared by mixing the precursors at 80 °C until homogeneous liquids were formed. Proteins were dissolved following the procedure described in the ESI.† :glycerol (1:2 mole ratio) and choline chloride:urea (1:2 mole ratio) were prepared by mixing the precursors at 80 °C until homogeneous liquids were formed. Proteins were dissolved following the procedure described in the ESI.† |
| This journal is © the Owner Societies 2017 |