
Structure advantage and peroxidase activity enhancement of deuterohemin-peptide–inorganic hybrid flowers†
Zijian Zhaoab,
Ji Zhangb,
Mingyang Wangc,
Zhi Wangb,
Liping Wangc,
Li Ma*bd,
Xuri Huang*a and
Zhengqiang Li*b
aInstitute of Theoretical Chemistry, Jilin University, Liutiao Road 2, Changchun 130023, Jilin Province, PR China. E-mail: huangxr@jlu.edu.cn
bKey Laboratory for Molecular Enzymology, Engineering of Ministry of Education, College of Life Science, Jilin University, 2699 Qianjin Street, Changchun 130012, Jilin Province, PR China. E-mail: lzq@jlu.edu.cn; Tel: +86-431-85155201
cNational Engineering Laboratory for AIDS Vaccine, College of Life Science, Jilin University, 2699 Qianjin Street, Changchun 130012, Jilin Province, PR China
dDepartment of Physics, Georgia Southern University, Statesboro, GA 30460, USA. E-mail: lma@georgiasouthern.edu
First published on 27th October 2016
Abstract
This work reports a facile method for hybridizing deuterohemin-peptide (DhHP-6) with copper phosphate to form deuterohemin-peptide–inorganic hybrid flowers (DhHP-6–Cu3(PO4)2) by self-assembly. The DhHP-6–Cu3(PO4)2 flowers have been characterized by scanning electron microscopy, Fourier transform infrared spectroscopy and solid state UV-vis. In the assembly process, the DhHP-6 peptides are fixed on Cu3(PO4)2 through the coordination of the end amino acids (Lys6) with copper(II) centers. The rearrangement of Lys6 prevents DhHP-6 aggregation. Hence, DhHP-6–Cu3(PO4)2 flowers exhibit a nearly 300% enhancement of peroxidase-like activity in comparison with free DhHP-6 in solution. Meanwhile, the DhHP-6–Cu3(PO4)2 flowers show stronger affinity towards 3,3,5,5-tetramethylbenzidine (TMB) and H2O2 than free DhHP-6 and horseradish peroxidase (HRP). In addition, EPR results provide direct evidences for the mechanisms of DhHP-6–Cu3(PO4)2 flower growth and activity enhancement. Furthermore, the sample presents excellent reusability and storage stability.
Introduction
Self-assembly processes are responsible for the generation of order without outside help. They involve components at different scales, such as molecules, cells and organisms.1–3 Molecular self-assembly is becoming a popular method to construct different types of micro- and nanostructured materials.4,5 It is a strategy directed at self-fabrication by controlling the assembly of ordered, integrated, and connected operational systems toward hierarchical growth. The interactions involved usually are non-covalent between inorganic and organic components that transit from a less ordered state to a highly ordered state without any external guidance.6In recent years, many studies have shown the ability of much larger bioorganic molecules, including peptides, polysaccharides, and nucleic acids, to self-assemble into uniform and well-ordered nanostructures.7–10 The nanostructures assembled from biomolecules attract increasing attention due to their biocompatibility, versatility, simple chemical and biological modification as well as easy availability for bottom-up fabrication.11,12 These nanostructures utilize the specificity of biological systems for biosensing, catalyzing, and highly specific molecular recognizing. Recently, Zare and co-workers report a facile biomolecular self-assembly for the synthesis of enzyme–inorganic hybrid flowers that show greatly enhanced catalytic activity and stability.13 The enzyme–inorganic hybrid flowers have been formed via the coordination of the copper phosphate nanocrystals and the nitrogen atoms of the amide groups in the enzymes. Subsequently, many other researchers have prepared hybrid flowers with various enzymes or proteins using the same synthetic route to achieve both high activity and stability.14–16 For example, the HRP incorporated into copper phosphate nanoflowers has been reported to show 300% enhancement of activity compared to free HRP.17 Trypsin immobilized on flowers is also determined to have 270% higher activity than free trypsin.18 In addition, we have recently reported a new simple method for fabricating the hybrid flowers using copper(II) ions and 20 kinds of amino acids,19 which provided the inspiration for this study.
Heme-peptide, as the peroxidase model compound, has been studied for many years due to its structural similarity to peroxidase and commercial availability.20,21 Heme-peptide complexes are obtained by covalent linking of peptide residues to the side chains of the porphyrins or proteolytic degradation from natural hemoproteins.22–24 They have great potential for numerous applications in chemistry and biology, such as biocatalysis, biosensor technology, degradation of pollutants, and drug or food processing. The best characterized among these complexes are the micro-peroxidases (MPs), a series of heme-peptides from degradation products of cytochrome c proteolysis. The heme moiety in the MPs structure is covalently bound through thioether links to the cysteine side chains and a histidine is maintained as the fifth heme iron ligand.24 Owing to the small size and relative simple structure, MPs possess reasonably high stability and could serve as good models to mimic the function of macrobiomolecules, such as hemeproteins and some enzymes that can be applied into biofuel cells, biocatalysis, and biosensors.25–27 However, in the absence of distal environment, the sixth coordination position of ferric centers in the models can be occupied by an amino group from another heme-peptide molecule which causes aggregation.28,29 In order to prevent the aggregation, immobilization of MPs in/on a suitable host material may prevent aggregation and render the heme more accessible to substrates. Previously, Ma's group has immobilized microperoxidase-11 (MP-11) into a mesoporous metal–organic framework (MOF) preventing aggregation and rendered the heme more accessible to substrates.30 While Yang's group has adsorbed MP-11 on boron nitride nanotubes to enhance in catalytic activity.31 Deuterohemin-β-Ala-His-Thr-Val-Glu-Lys (DhHP-6) is a synthesized heme-peptide with six amino acid residues (Fig. S1†).32 DhHP-6, referred to the residues of micro-peroxidase modified on the propionic acid side chain of iron porphyrin, presents peroxidase-like activity.33 In addition, DhHP-6 gives biological activities in inhibiting apoptosis against reactive oxygen species and improving cell survivals.34,35 Similar to micro-peroxidase, DhHP-6 has a tendency to aggregate in aqueous solutions that can be attributed to the inter-molecular interaction of Lys at the end of six-amino-acid chain in DhHP-6 with the ferric center of another DhHP-6 through the 6th coordination position.36 In order to decrease the interaction between DhHP-6s, PEGylation has been applied on the Lys of DhHP-6 to reduce the aggregation and to improve its stability.37 However, in this strategy, it is difficult to remove excess PEG, causing the decrease of the peroxidase activity of PEGylated DhHP-6.37 Hence, it is necessary to develop a facile strategy for improving the stability and activity of DhHP-6.
In this study, hierarchical flowerlike peptide–inorganic hybrid DhHP-6–Cu3(PO4)2 has been synthesized with a simple approach using DhHP-6, copper sulphate aqueous solution, and phosphate buffered saline.13 After fixation of DhHP-6 peptides on the copper phosphate nanocrystals, the aggregation of free DhHP-6 is prevented and the structure stability is improved. The enzyme-like activity of DhHP-6–Cu3(PO4)2 has been evaluated by catalyzing the oxidation of 3,3,5,5-tetramethylbenzidine (TMB) in the presence of hydrogen peroxide (H2O2) and the enhancement of catalytic activity of the DhHP-6–Cu3(PO4)2 flowers achieved in comparison with free DhHP-6. EPR spectroscopy has been used to further investigate the mechanisms of flower-growth and enzyme-like activity enhancement, providing direct evidences for the interaction between copper ions and amino acids. Furthermore, recyclability and high stability in storage of the hybrid flowers have been observed.
Experimental
Materials
DhHP-6 was obtained from College of Life Science, Jilin University (Changchun, China)32 and the structure of DhHP-6 is shown in Fig. S1.† Hydrogen peroxide (H2O2, 30%, w/v) was purchased from Shantou Xilong Chemical Factory (Guangdong, China). 3,3,5,5-Tetramethylbenzidine (TMB), horseradish peroxidase (HRP) (Enzyme Commission Number 1.11.1.7, Mw 44 kDa) and copper sulphate pentahydrate (CuSO4·5H2O) were purchased from Sigma-Aldrich (St. Louis, USA). All other chemicals and reagents were of analytical grade unless specified. Phosphate buffered saline (PBS) was used at 50 mM in saline. All aqueous solutions were prepared with Milli-Q water.Synthesis of DhHP-6–Cu3(PO4)2 hybrid flowers
The synthesis of DhHP-6–Cu3(PO4)2 flowers was accomplished using a previous reported method with minor modifications of scaling up.13 First, 120 mM CuSO4 stock solution in ultrapure water was freshly prepared. Then, 2 mL of CuSO4 stock solution was added to 300 mL DhHP-6 solution in PBS (pH 7.4) with various DhHP-6 concentrations ranging from 0.006 to 0.06 mg mL−1. The final mixtures were left undisturbed for 72 hours at 25 °C, and then centrifuged to obtain the precipitates. To remove unreacted components, the collected precipitates were washed at least 3 times with water and centrifuged at 12![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 rpm for 10 min after each washing then dried under vacuum at room temperature. Cu3(PO4)2 control sample was prepared by the same method in the absence of DhHP-6. Targeted, actual, and unreacted DhHP-6 concentrations in the reaction mixture were determined by colorimetric method at 389 nm (absorbance of DhHP-6, ε389 = 54520 M−1 cm−1). The encapsulation yield was determined from the percentage ratio of reacted to total DhHP-6 added. The weight percentage of DhHP-6 in the DhHP-6–Cu3(PO4)2 flowers was calculated based on the encapsulation yield and the weight of the DhHP-6–Cu3(PO4)2.13
000 rpm for 10 min after each washing then dried under vacuum at room temperature. Cu3(PO4)2 control sample was prepared by the same method in the absence of DhHP-6. Targeted, actual, and unreacted DhHP-6 concentrations in the reaction mixture were determined by colorimetric method at 389 nm (absorbance of DhHP-6, ε389 = 54520 M−1 cm−1). The encapsulation yield was determined from the percentage ratio of reacted to total DhHP-6 added. The weight percentage of DhHP-6 in the DhHP-6–Cu3(PO4)2 flowers was calculated based on the encapsulation yield and the weight of the DhHP-6–Cu3(PO4)2.13
Characterization methods
For morphology study, DhHP-6–Cu3(PO4)2 flowers loaded on monocrystalline silicon were examined using a XL-30 scanning electron microscope (Philips Company, Germany). To confirm the hybridizing between DhHP-6 and Cu3(PO4)2, FTIR spectra was obtained using VERTEX 80v vacuum FTIR (Bruker, Germany) in a wavelength range of 370 to 2500 cm−1. The FTIR samples were prepared by mixing with KBr (spectroscopic grade) and compressed into thin films. Solid-state UV-vis spectra were measured on U-4100 Spectrophotometer (Hitachi, USA) to conform ferric centers in DhHP-6 after hybridizing. A quartz cell was used and spectra were collected in a range 300–800 nm using BaSO4 as a reference.To study the coordination changes on metal centers (Cu(II) and Fe(III)), Electron Paramagnetic Resonance (EPR) measurements were performed using a Bruker EMXplus X-band spectrometer equipped with ColdEdge CH-210N Cryocooler. Samples were loaded in quartz tubes (1 mm i.d., 2 mm o.d., Sigma USA). The tubes were frozen in liquid nitrogen before the measurements at low the temperature. Spectra were recorded under the following EPR conditions: centerfield, 2500 G; scan range, 5000 G; field modulation, 5.0 G; microwave power, 2 mW; time constant, 0.1 s.
Enzyme-like activity and kinetic analysis
The enzyme-like activity of the DhHP-6–Cu3(PO4)2 flowers was tested according TMB peroxidase assay.31 This assay utilized colorimetric absorption of oxidized TMB at 652 nm (ε652 nm = 39000 M−1 cm−1) in order to avoid assay interference with the Soret band of DhHP-6. Briefly, suspended DhHP-6–Cu3(PO4)2 flowers weighted at 3.8 μg DhHP-6 equivalent in 1 mL PBS (pH 6.0) were used to catalytically convert substrate H2O2 to water and oxidize TMB to blue at same time. The color reactions were monitored for 2 min in kinetic mode on UV-2770 at 35 °C. To demonstrate activity enhancement of DhHP-6 in flower structure, the activity of free DhHP-6 in solution was also determined according to the same procedures. One unit specific activity (U mg−1) of DhHP-6–Cu3(PO4)2 flowers or free DhHP-6 is defined as 1 mg of DhHP-6 that catalyzes 1.0 μmol of H2O2 per minute.
The apparent steady-state kinetic analysis was carried out by varying concentrations of TMB at a fixed concentration of H2O2 and vice versa. The apparent kinetic parameters, maximum reaction rate (Vmax) and Michaelis–Menten constant (Km), were determined. First, the reaction rates (velocities) were obtained from the slops of linear regression curves of absorbance versus time at different concentrations of H2O2 ranging from 0.1 mM to 1.0 mM and a fixed TMB concentration of 0.8 mM. The same steps were applied for determining the rates of the reactions at varying concentrations of TMB from 0.02 mM to 0.83 mM and a fixed H2O2 concentration of 2 mM. Second, a plot of velocity (V) versus substrate concentration [S] was created and Vmax and Km were determined by nonlinear fitting of the plot to the Michaelie–Menten equation, V = Vmax[S]/(Km + [S]), where V is the velocity obtained from the first step, Vmax is the maximal velocity, [S] corresponding concentration of the substrate and Km is the Michaelis–Menten constant related to the actual affinity of the enzyme for the substrate. The smaller Km would reflect the higher affinity for the substrate.
Stabilities
Recycling stability was tested in this study. The DhHP-6–Cu3(PO4)2 flowers were recycled after each catalytic reaction at 2 mM H2O2 and 0.8 mM TMB through centrifugation and thoroughly rinsing with PBS (pH 6.0) to use for next reaction cycle. The recycling stability of DhHP-6–Cu3(PO4)2 flowers was evaluated by measuring the enzyme-like activity in each successive cycle. The recycling efficiency was defined as the ratio of the activity of DhHP-6–Cu3(PO4)2 flowers after recycling to their initial activity.For storage stability testing, the free DhHP-6 and DhHP-6–Cu3(PO4)2 flowers were stored at 4 °C for a certain period of time. The storage stability was evaluated in terms of storage efficiency and defined as the ratio of the activity of free DhHP-6 or DhHP-6–Cu3(PO4)2 flowers after storage to their initial activity.
Results and discussions
Morphology characterization of DhHP-6–Cu3(PO4)2 flowers
Fig. 1 shows the morphologies of the DhHP-6–Cu3(PO4)2 flowers prepared from four starting (targeted) concentrations of DhHP-6 at 0.06, 0.024, 0.012 and 0.006 mg mL−1. SEM images in Fig. 1A, C, E and G indicate the diameters of the flowers becoming smaller from about 30 μm to 15 μm when DhHP-6 concentrations reduced by 10 times. The same growth pattern was observed in Cu3(PO4)2 hybrid flowers of silk protein sericin by Koley38 but not in Ge's preparations of hybrid flowers of carbonic anhydrase or lipase.13 Nevertheless, the high resolution images in Fig. 1B, D, F and H reveal that flowers have dense structures when prepared at higher starting DhHP-6 concentrations and larger porous size at low concentrations. However, different from the protein–inorganic hybrid flowers,38,39 the petals of DhHP-6–Cu3(PO4)2 are always thin regardless of the peptide concentrations as observed in our previous work of amino acids-incorporated nanoflowers.19 The results demonstrate that the size of flowers is dependent on preparation conditions including the precursor concentrations, incubation times and etc. The thickness of petals could be influenced by types of proteins or amino acids. In the process of protein–inorganic hybrid flowers, the amide groups on proteins serve as “glue” to link Cu3(PO4)2 petals and to form flowers eventually.38 There are multiple amide groups on the protein backbones which means there are many binding sites on proteins. Hence, the binding between Cu3(PO4)2 crystals becomes tight as protein concentration gradually increasing. This could result in thickening in petals and possibly shrinking in size of the protein–inorganic hybrid flowers. Different from protein or enzyme, DhHP-6 has only one amide group on the Lys of the peptide which cannot provide many binding sites to Cu3(PO4)2 crystals. Therefore, DhHP-6 will make the flowers with thin and large porous when formed at low concentrations, but thin petals and narrow spaces between them at high concentrations. | ||
| Fig. 1 SEM images of DhHP-6–Cu3(PO4)2 flowers with varying targeted concentrations of DhHP-6 with low resolution at left and high at right: (A) and (B) 0.06 mg mL−1, (C) and (D) 0.024 mg mL−1, (E) and (F) 0.012 mg mL−1, (G) and (H) 0.006 mg mL−1. | ||
To study the mechanism of formation of the DhHP-6–Cu3(PO4)2 flowers, EPR spectroscopy is used to monitor coordination changes of paramagnetic Cu(II) centers during the reaction. The results provide direct evidences of the binging of copper centers with lysine amino acids on DhHP-6. Detailed results are presented in the EPR section of this paper.
FTIR and solid state absorption studies
The FTIR spectra of the DhHP-6, Cu3(PO4)2, and DhHP-6–Cu3(PO4)2 flowers are shown in Fig. 2, spectrum (a), (b), and (c) respectively. In spectrum (a), the characteristic bands at 1655 cm−1 and 1538 cm−1 arise from the amide bonds that link the amino acids called amide I (C![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) O stretching) and amide II (N–H bending).40 These two bands keep no changes in the positions after hybridizing shown in spectrum (c) that indicates the peptide structure of DhHP-6 keeping no changes before and after. On the other hand, the peaks at 988, 623 and 556 cm−1 in spectrum (b) are assigned to the characteristic vibrations of P–O.41 All three peaks are observed in the spectrum (c) of DhHP-6–Cu3(PO4)2. FTIR spectra confirm hybrid formation of the flowers without changing the secondary structure of the peptides.
O stretching) and amide II (N–H bending).40 These two bands keep no changes in the positions after hybridizing shown in spectrum (c) that indicates the peptide structure of DhHP-6 keeping no changes before and after. On the other hand, the peaks at 988, 623 and 556 cm−1 in spectrum (b) are assigned to the characteristic vibrations of P–O.41 All three peaks are observed in the spectrum (c) of DhHP-6–Cu3(PO4)2. FTIR spectra confirm hybrid formation of the flowers without changing the secondary structure of the peptides.
 | ||
| Fig. 2 FTIR spectra of (a) DhHP-6, (b) Cu3(PO4)2 and (c) DhHP-6–Cu3(PO4)2 flowers. | ||
To confirm ferric centers of DhHP-6 are still intact after hybridizing, solid state UV-vis spectra are presented before and after flower formation in Fig. 3. The absorption peak at 390 nm in spectrum (a) is the characteristic Soret band of free DhHP-6 due to π–π* transition of porphyrin under the influence of Fe3+ centers. A similar peak is observed in the flowers' spectrum (c) indicating no interruption on the heme structure of DhHP-6 during the hybridizing. Likewise, the characteristic absorption band at 707 nm (ref. 42) in spectrum (b) of Cu3(PO4)2 appears in the spectrum (c) of DhHP-6–Cu3(PO4)2. The spectrum changes can be revealed from the color changes after incorporating DhHP-6 in Cu3(PO4)2 as easily visualized in Fig. 2S.† Due to the additional absorption in blue range, DhHP-6–Cu3(PO4)2 shows green but Cu3(PO4)2 in light blue. The results of solid-state UV-vis confirm that the flowers are composed of Cu3(PO4)2 and DhHP-6 components and ferric centers are intact.
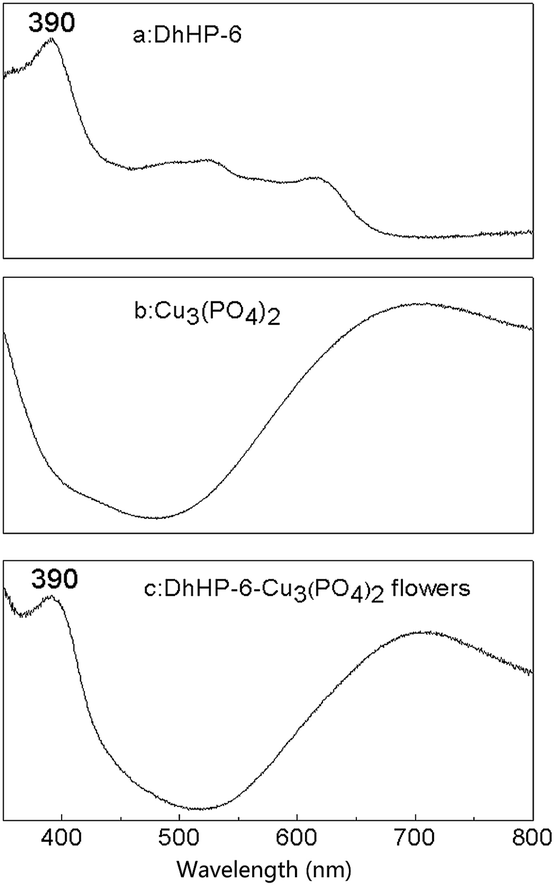 | ||
| Fig. 3 Solid-state UV-vis spectra of (a) DhHP-6, (b) Cu3(PO4)2 and (c) DhHP-6–Cu3(PO4)2 flowers. | ||
Enzyme-like activity and kinetics studies
The activities of five DhHP-6–Cu3(PO4)2 flowers prepared at different starting DhHP-6 concentrations have been compared with free DhHP-6 by TMB peroxidase assay as described in the method. The absorbance of the oxidized TMB at 652 nm as a function of time for four types of DhHP-6–Cu3(PO4)2 flowers, free DhHP-6 and Cu3(PO4)2 prepared in absence of DhHP-6 are shown in Fig. S3.† The specific activities are calculated based on that one unit (U) is 1.0 mg of DhHP-6 that catalyzes 1.0 μmol of H2O2 per minute at pH 6.0, 35 °C. The encapsulation efficiencies, weight percentages, and corresponding specific activities of these flowers are listed in Table S1.† Overall, the flowers show higher activities than free DhHP-6 at equivalent molar concentrations except the flowers prepared from the highest starting concentration at 0.06 mg mL−1. In particular, the flowers prepared from the lowest concentration at 0.006 mg mL−1 exhibit nearly 300% activity increase compared with free DhHP-6. The structural advantages of the flowers over free peptides are investigated using EPR spectroscopy and discussed in details in the EPR studies later. Compared among the flowers, activity increases 7 times activity increasing when the starting concentration is reduced from 0.06 to 0.006 mg mL−1. The flowers with the highest activity displays smaller diameter and larger pore size under SEM shown in Fig. 1G and H. These features may facilitate the substrates entering resulting in the activity enhancement. Furthermore, the flowers sample prepared from highest starting concentration has lowest activity which may be too tight in the structure (Fig. 1B) to be efficiently reacting with the substrates. In addition, the effects of pH (pH 4.0–8.0) and temperature (20–50 °C) on the catalytic activity are explored (Fig. S4A and B†). Based on the results, pH 6.0 and 35 °C are selected as the optimal conditions for both DhHP-6 and flowers in activity and kinetic studies. It is concluded that DhHP-6–Cu3(PO4)2 flowers with loosen porous structure have higher peroxidase-like activity than free DhHP-6 over broad pH and temperature ranges. Besides the morphology advantages of the small and loosen flowers, the structure advantages are also studied at ligand binding level of the ferric centers by EPR.To further investigate the structure advantages of the flowers, DhHP-6–Cu3(PO4)2 synthesized from lowest starting DhHP-6 concentration has been used for the kinetic studies at the optimized reaction condition of pH 6 and temperature of 35 °C. The kinetic data have been obtained by changing the concentration of one substrate while keeping the other constant and vice versa. The velocity of the reaction follows a typical Michaelis–Menten curves the tested range of substrate concentrations (TMB (Fig. 4A) and H2O2 (Fig. 4B)). Both the maximum initial velocity (Vmax) and Michaelis–Menten constant (Km) are calculated from Lineweaver–Burk plots, and the results are listed in Table 1. By comparing the apparent kinetic parameters, the Km value of DhHP-6–Cu3(PO4)2 is nearly nine times and six times lower than that of free DhHP-6 and HRP, respectively when using TMB as the substrate. In addition, the Km value of DhHP-6–Cu3(PO4)2 when varying H2O2 concentrations is nearly two times and seven times lower than that in free DhHP-6 and HRP, respectively. It is known that Km is identified as an indicator of enzyme affinity to substrate and a lower Km value represents a stronger affinity of enzyme for substrates and vice versa. So the decreasing in Km values suggests that DhHP-6–Cu3(PO4)2 flowers have higher catalytic affinity toward TMB and H2O2 than that of free DhHP-6 and HRP. In addition, the apparent Vmax values of the DhHP-6–Cu3(PO4)2 flowers with TMB and H2O2 as the substrates are near 2 times faster than those of free DhHP-6.
 | ||
| Fig. 4 Steady-state kinetic assay of DhHP-6–Cu3(PO4)2 flowers (black) and free DhHP-6 (red). (A) The concentration of H2O2 at 2 mM and the TMB concentrations varied from 0.02 mM to 0.86 mM. (B) The concentration of TMB at 0.8 mM and the H2O2 concentrations varied from 0.1 mM to 1.0 mM. | ||
| Catalyst | Substrate | Km (mM) | Vmax (10−8 M s−1) |
|---|---|---|---|
| DhHP-6 | TMB | 0.60 | 3.65 |
| H2O2 | 0.87 | 3.80 | |
| DhHP-6–Cu3(PO4)2 flowers | TMB | 0.071 | 5.63 |
| H2O2 | 0.53 | 6.11 | |
| HRP43 | TMB | 0.43 | 10 |
| H2O2 | 3.70 | 8.71 |
For investigating the mechanism of the activity of DhHP-6–Cu3(PO4)2 flowers, we measure the activity of DhHP-6–Cu3(PO4)2 over a range of TMB and H2O2 concentrations. The double reciprocal plots of initial velocity versus one substrate concentrations are obtained in the tested range (Fig. 5A and B). The slope of the each plot is nearly parallel each other, which is characteristics of a ping-pong mechanism, as observed for peroxidase enzyme (HRP) in Sun's work.44 This indicates that DhHP-6–Cu3(PO4)2 flowers bind and react with the first substrate, and release the first product before reacting with the second substrate.
 | ||
| Fig. 5 Double reciprocal plots of velocity of the reaction of DhHP-6–Cu3(PO4)2 flowers as a function of (A) TMB concentration with 0.6, 0.8 and 1.0 mM of H2O2 and (B) H2O2 with 0.2, 0.4, and 0.6 mM of TMB. The velocity (V) of the reaction was measured using 3.8 μg equivalent DhHP-6 in DhHP-6–Cu3(PO4)2 flowers prepared with 0.006 mg mL−1 starting DhHP-6 in 1 mL of PBS with pH 6.0 at 35 °C. | ||
EPR studies for growth mechanism and activity enhancement
There are many discussions regarding the fabrication mechanisms of protein–inorganic hybrid flowers.13,39 It has been consistently believed that the coordination between copper ions and amino acid backbones provides the nucleation of the primary crystals.13 EPR method could be used to directly detect the coordination changes of paramagnetic Cu(II) centers by analyzing the spectrum shape.45 The X-band EPR spectra of precursor CuSO4, fragment Cu3(PO4)2, DhHP-6–Cu3(PO4)2 and albumin–Cu3(PO4)2 are presented in Fig. S6A–D† respectively. At room temperature, the mononuclear CuSO4 shows close to isotropic spectrum with rhombic distortions as observed previously.46 While the tri-nuclear Cu3(PO4)2 prepared in absence of amino acids has very broaden axial EPR signal due to intra-molecular spin–spin interaction between Cu(II) centers. Both EPR spectra (Fig. S6A and B†) lack the nuclear hyperfine structure (NHS) that is typical Cu(II) in inhomogeneous solids. Importantly when hybridized with DhHP-6 or albumin, the Cu(II) centers give axial spectra with larger g⊥ = 2.07 than g⊥ = 2.03 in Cu3(PO4)2 and much narrow spectra. The narrowing makes NHS to be resolved clearly with HF splitting constants of A∥ = 159 G and A∥ = 141 G in DhHP-6–Cu3(PO4)2 and albumin–Cu3(PO4)2 respectively (A⊥ is still not able to be determined accurately). These spectra suggest that the Cu(II) centers have typical square pyramidal or elongated octahedron geometry after hybridizing with peptides or proteins. Nevertheless the EPR changing after linking amino acids on Cu3(PO4)2 indicates the changing in axial coordination and homogeneity. The EPR results provide direct evidences for the interaction between copper ions with amino acids. The similarity in spectrum shapes in both hybrid flowers indicates the nature of Cu(II) coordination is similar regardless of the interaction with peptides or proteins.Different from traditionally prepared immobilized enzymes,47 a number of previously reported enzyme–inorganic hybrid flowers exhibit much higher activities than free enzymes.48 In this study, to investigate the mechanism of the activity enhancement of DhHP-6, low temperature EPR of DhHP-6–Cu3(PO4)2 (prepared from 0.006 mg mL−1 DhHP-6) has been compared with free DhHP-6. This study focuses on the coordination of the paramagnetic Fe(III) centers. It has been known that a ferric center with no or weak axial ligand at 6th position given a high-spin state (S = 2/3), provides great accessibility to substrates resulting high enzyme activity.29 While when a strong ligand occupies the 6th position, the Fe(III) centers present a low-spin state (S = 1/2) as a result the ligand hard to be replaced by substartes. Fig. 6 exhibits EPR spectra of free DhHP-6 in (A) and DhHP-6–Cu3(PO4)2 flowers in (B). Free DhHP-6 peptides give two groups of EPR signals, labelled I and II which are from high-spin and low-spin species of ferric centers respectively. The high-spin signal with the lowest g value (species I) at 1.99 and the highest g at 6.00 are from the |±1/2> doublet of high spin (HS) S = 5/2 ferric centers with E/D ≈ 0.0 and the signal g = 4.24 is from |±3/2> with E/D ≈ 0.3.49 Besides the HS signals, significant amount of LS S = 1/2 also present in free DhHP-6 (species II, g1 = 2.93, g2 = 2.27 and g3 = 1.56). Like other micro-peroxidases,29 free DhHP-6 has a tendency to inter-molecularly link together through the Lys6 at the end of the peptide with a ferric center of another DhHP-6 resulting the formation of six-coordinated low-spin (LS) complex (Scheme 1). However, after hybridizing DhHP-6–Cu3(PO4)2, EPR shows disappearance of the LS species II and no significant changes on the positions of HS EPR signals (species I, g = 5.98, 4.23 and 1.99). The absence of LS state may indicate that, in the process of self-assembly, the Lys6 residue on a DhHP-6 switches its coordination from adjacent Fe(III) in free DhHP-6 to a Cu(II) center on Cu3(PO4)2 petal. As a result, the availability of the sixth positions of Fe(III) to substrates increases which enhances the enzyme mimetic activity of DhHP-6–Cu3(PO4)2. The new EPR signals labelled as species III, g1 = 2.27 and g2 = 2.06, in the flowers are from Cu(II) centers indicating anisotropic characteristic of the centers (Table 2).45
 | ||
| Fig. 6 EPR spectra of (A) free DhHP-6 and (B) DhHP-6–Cu3(PO4)2 flowers measured at 12 K. | ||
 | ||
| Scheme 1 Proposed growth mechanism of DhHP-6–Cu3(PO4)2 flowers. | ||
| Samples | g-Values | ||
|---|---|---|---|
| Species I | Species II | Species III | |
| DhHP-6 | 6.0 | 2.93 | |
| 4.24 | 2.27 | ||
| 1.99 | 1.56 | ||
| DhHP-6–Cu3(PO4)2 flowers | 5.98 | 2.27 | |
| 4.23 | 2.06 | ||
| 1.99 | |||
All the facts and results above prove that DhHP-6–Cu3(PO4)2 flowers facilitate high catalytic activity of substrate oxidation, which can be attributed to two factors. First, the hybridizing DhHP-6 with Cu3(PO4)2 prevents the tendency of free peptide aggregation in solution caused by its inter-chain coordination to the sixth position of Fe(III), leading the heme to be less accessible (Scheme 1). This aggregation is believed to be through Lys6 at the end of DhHP-6 peptide with another DhHP-6. However, in DhHP-6–Cu3(PO4)2 flowers, the Lys6 amino acids coordinate with Cu(II) centers on Cu3(PO4)2 petals indicated by EPR results. For this reason, the activity center of DhHP-6 on Cu3(PO4)2 is more open to substrates and DhHP-6–Cu3(PO4)2 flowers will have higher catalytic activity and possess higher affinity to substrates than free DhHP-6. Second, the morphology of the flowers is also an important factor.50 It is observed that the flowers prepared from 0.006 mg mL−1 starting concentration have higher activity than other flowers in this study. The thin petals, large pore sizes in this DhHP-6–Cu3(PO4)2 could facilitate high surface-to-volume ratio which could result in incremental concentration of substrates around the surface region.
Stability of DhHP-6–Cu3(PO4)2 flowers
One of the benefits for enzyme immobilization is that it makes the enzyme feasible to be reused since it can be easily recovered or separated from the reaction medium. Fig. 7 reveals that DhHP-6–Cu3(PO4)2 flowers still retain 65% of its peroxidase-like activity after eight cycles of use. The decrease of peroxidase-like activity could be primarily due to the following reasons: (i) the active sites of DhHP-6 are occupied with the reaction products; (ii) flowers are loss during the eluting process. The storage stability of free DhHP-6 and DhHP-6–Cu3(PO4)2 flowers is investigated by determining the residual activities after keeping them for a period of 40 days at 4 °C and the results are given in Fig. 8. The DhHP-6–Cu3(PO4)2 flowers show better stability than free DhHP-6, preserving their initial activity over 84% after 40 days, whereas free DhHP-6 loses nearly 60% of the initial activity. The poor storage stability of free DhHP-6 could be attributed the DhHP-6 tendency to form aggregates during long storage in aqueous solutions.37 In contrast, the DhHP-6–Cu3(PO4)2 flowers present stable structure in aqueous solutions, and the immobilized DhHP-6 disperse on the nanopetals, thus prevent the DhHP-6 aggregation. These advantages in stability and reusability of DhHP-6–Cu3(PO4)2 flowers could be widely employed in various applications for biosensor and catalysis.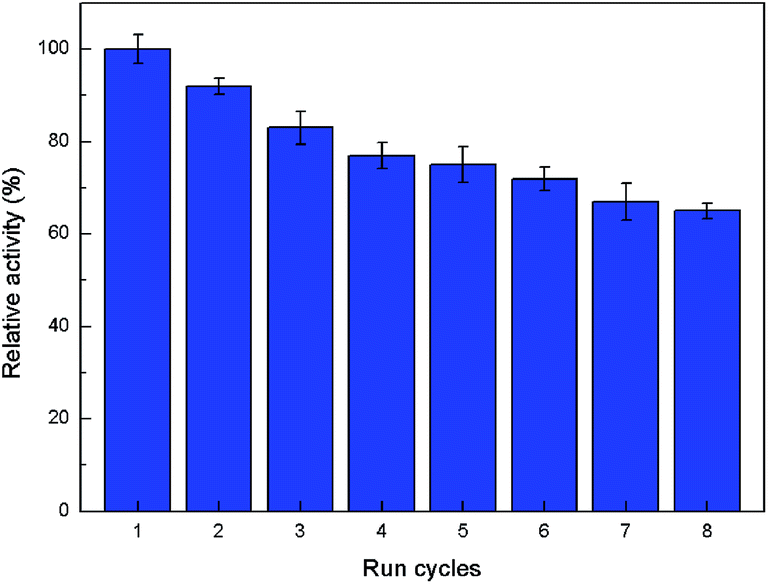 | ||
| Fig. 7 Reusability of DhHP-6–Cu3(PO4)2 flowers. | ||
 | ||
| Fig. 8 Storage of free DhHP-6 and DhHP-6–Cu3(PO4)2 flowers. | ||
Conclusions
In summary, we present a simple method for synthesizing peptide–inorganic hybrid flowers (DhHP-6–Cu3(PO4)2) from free DhHP-6 (an artificial peroxidase). This self-assembly approach can inhibit the heme-peptide aggregation and increase the affinity of the ferric centers to substrates. The newly synthesized flowers enhance the peroxidase-like activity when compared with free DhHP-6. EPR results provide the direct evidences that, during the hybridizing, the interaction occurs between copper ions and Lys6 amino acids which coordinate with Fe(III) centers in precursor DhHP-6. As a result, the 6th position of ferric centers is free and the coordination change of the centers from LS to HS is observed in EPR spectra. This could be the reason that DhHP-6–Cu3(PO4)2 has higher peroxidase-like activity than free DhHP-6. Moreover, the DhHP-6–Cu3(PO4)2 flowers exhibit good reusability and stability. We believe that the new heme peptide-incorporated flowers have great potential applications in biosensors, bioanalytical devices, and environmental chemistry.Acknowledgements
The authors are grateful for the financial support from the National High Technology Research and Development Program of China (“863” Program, 2012AA022202B) and the basic scientific research service fee of Jilin University (No. 451160301163).Notes and references
- G. M. Whitesides, J. P. Mathias and C. T. Seto, Science, 1991, 254, 1312–1319 CAS.
- G. M. Whitesides and B. Grzybowski, Science, 2002, 295, 2418–2421 CrossRef CAS PubMed.
- H. Li, J. D. Carter and T. H. Labean, Mater. Today, 2009, 12, 24–32 CrossRef CAS.
- Y. Guo, L. Xu, H. Liu, Y. Li, C. M. Che and Y. Li, Adv. Mater., 2015, 27, 985–1013 CrossRef CAS PubMed.
- P. Zhou, R. Shi, J. F. Yao, C. F. Sheng and H. Li, Coord. Chem. Rev., 2015, 292, 107–143 CrossRef CAS.
- S. Mann, Nat. Mater., 2009, 8, 781–792 CrossRef CAS PubMed.
- S. Toledano, R. J. Williams, V. Jayawarna and R. V. Ulijn, J. Am. Chem. Soc., 2006, 128, 1070–1071 CrossRef CAS PubMed.
- S. M. Brosnan, H. Schlaad and M. Antonietti, Angew. Chem., Int. Ed., 2015, 54, 9715–9718 CrossRef CAS PubMed.
- K. S. Lim, D. Y. Lee, G. M. Valencia, Y. W. Won and D. A. Bull, Adv. Funct. Mater., 2015, 25, 5444 CrossRef.
- X. Yan, Y. Cui, Q. He, K. Wang and J. Li, Chem. Mater., 2008, 20, 1522–1526 CrossRef CAS.
- X. Yan, P. Zhu and J. Li, Chem. Soc. Rev., 2010, 39, 1877–1890 RSC.
- S. Zhang, Nat. Biotechnol., 2003, 21, 1171–1178 CrossRef CAS PubMed.
- J. Ge, J. Lei and R. N. Zare, Nat. Nanotechnol., 2012, 7, 428–432 CrossRef CAS PubMed.
- Y. Huang, X. Ran, Y. Lin, J. Ren and X. Qu, Chem. Commun., 2015, 51, 4386–4389 RSC.
- Z. Lin, Y. Xiao, Y. Yin, W. Hu, W. Liu and H. Yang, ACS Appl. Mater. Interfaces, 2014, 6, 10775–10782 CAS.
- Y. Yu, X. Fei, J. Tian, L. Xu, X. Wang and Y. Wang, Colloids Surf., B, 2015, 130, 299–304 CrossRef CAS PubMed.
- B. Somturk, M. Hancer, I. Ocsoy and N. Özdemir, J. Chem. Soc., Dalton Trans., 2015, 44, 13845–13852 RSC.
- Z. Lin, Y. Xiao, L. Wang, Y. Yin, J. Zheng, H. Yang and G. Chen, RSC Adv., 2014, 4, 13888–13891 RSC.
- Z. F. Wu, Z. Wang, Y. Zhang, Y. L. Ma, C. Y. He, H. Li, L. Chen, Q. S. Huo, L. Wang and Z. Q. Li, Sci. Rep., 2016, 6, 22412 CrossRef CAS PubMed.
- T. Tatsuma and T. Watanabe, Anal. Chem., 1991, 63, 1580–1585 CrossRef CAS PubMed.
- F. Nastri, L. Lista, P. Ringhieri, R. Vitale, M. Faiella, C. Andreozzi, P. Travascio, O. Maglio, A. Lombardi and V. Pavone, Chem.–Eur. J., 2011, 17, 4444–4453 CrossRef CAS PubMed.
- E. S. Ryabova, P. Rydberg, M. Kolberg, E. Harbitz, A. L. Barra, U. Ryde, K. K. Andersson and E. Nordlander, J. Inorg. Biochem., 2005, 99, 852–863 CrossRef CAS PubMed.
- P. A. Arnold, D. R. Benson, D. J. Brink, M. P. Hendrich, G. S. Jas, M. L. Kennedy, D. T. Petasis and M. Wang, Inorg. Chem., 1997, 36, 5306–5315 CrossRef CAS.
- H. M. Marques, J. Chem. Soc., Dalton Trans., 2007, 251, 4371–4385 RSC.
- K. S. Prasad, C. Walgama and S. Krishnan, RSC Adv., 2015, 5, 11845–11849 RSC.
- A. Ramanavicius and A. Ramanaviciene, Fuel Cells, 2009, 9, 25–36 CrossRef CAS.
- H. Wariishi, M. Kabuto, J. Mikuni, M. Oyadomari and H. Tanaka, Biotechnol. Prog., 2002, 18, 36–42 CrossRef CAS PubMed.
- J. S. Wang and H. E. V. Wart, J. Phys. Chem., 1989, 93, 7925–7931 CrossRef CAS.
- A. Riposati, T. Prieto, C. S. Shida, I. L. Nantes and O. R. Nascimento, J. Inorg. Biochem., 2006, 100, 226–238 CrossRef CAS PubMed.
- V. Lykourinou, Y. Chen, X. S. Wang, L. Meng, T. Hoang, L. J. Ming, R. L. Musselman and S. Ma, J. Am. Chem. Soc., 2011, 133, 10382–10385 CrossRef CAS PubMed.
- R. Li, J. Liu, L. Li, H. Wang, Z. Weng, S. K. Lam, A. Du, Y. Chen, C. J. Barrow and W. Yang, Chem. Commun., 2014, 50, 225–227 RSC.
- S. Guan, P. Li, J. Luo, Y. Li, L. Huang, G. Wang, L. Zhu, H. Fan, W. Li and L. Wang, Free Radical Res., 2010, 44, 813–820 CrossRef CAS PubMed.
- (a) H. Zhou, W. Jiang, N. An, Q. Zhang, S. Xiang, L. Wang and J. Tang, RSC Adv., 2015, 5, 42728–42735 RSC; (b) Q. G. Dong, Y. Zhang, M. S. Wang, J. Feng, H. H. Zhang, Y. G. Wu, T. J. Gu, X. H. Yu, C. L. Jiang and Y. Chen, Amino Acids, 2012, 43, 2431–2441 CrossRef CAS PubMed.
- L. Huang, P. Li, G. Wang, S. Guan, X. Sun and L. Wang, Free Radical Res., 2013, 47, 316–324 CrossRef CAS PubMed.
- S. Guan, P. Li, J. Luo, Y. Li, L. Huang, G. Wang, L. Zhu, H. Fan, W. Li and L. Wang, Free Radical Res., 2010, 44, 813–820 CrossRef CAS PubMed.
- Z. Zhao, D. Wang, M. Wang, X. Sun, L. Wang, X. Huang, L. Ma and Z. Li, RSC Adv., 2016, 6, 78858–78864 RSC.
- H. Lin, Y. Li, H. Zhou, L. Wang, H. Cao, J. Tang and W. Li, J. Appl. Polym. Sci., 2013, 128, 706–711 CrossRef CAS.
- P. Koley, M. Sakurai and M. Aono, ACS Appl. Mater. Interfaces, 2016, 8, 2380–2392 CAS.
- Y. Yin, Y. Xiao, G. Lin, Q. Xiao, Z. Lin and Z. Cai, J. Mater. Chem. B, 2015, 3, 2295–2300 RSC.
- Z. Zhao, X. Xie, Z. Wang, Y. Tao, X. Niu, X. Huang, L. Liu and Z. Li, J. Biosci. Bioeng., 2016, 121, 645–651 CrossRef CAS PubMed.
- I. S. Cho, D. W. Kim, S. Lee, C. H. Kwak, S. T. Bae, J. H. Noh, S. H. Yoon, H. S. Jung, D. W. Kim and K. S. Hong, Adv. Funct. Mater., 2008, 18, 2154–2162 CrossRef CAS.
- Y. R. Morgan, P. Turner, B. J. Kennedy, T. W. Hambley, P. A. Lay, J. R. Biffin, H. L. Regtop and B. Warwick, Inorg. Chim. Acta, 2001, 324, 150–161 CrossRef CAS.
- L. Gao, J. Zhuang, L. Nie, J. Zhang, Y. Zhang, N. Gu, T. Wang, J. Feng, D. Yang and S. Perrett, Nat. Nanotechnol., 2007, 2, 577–583 CrossRef CAS PubMed.
- J. Tian, Q. Liu, A. M. Asiri, A. H. Qusti, A. O. Alyoubi and X. Sun, Nanoscale, 2013, 5, 11604–11609 RSC.
- M. Valko, P. Pelikan, S. Biskupič and M. Mazur, Chem. Pap., 1990, 44, 805–813 CAS.
- A. Tani, T. Ueno, C. Yamanaka, M. Katsura and M. Ikeya, Appl. Radiat. Isot., 2005, 62, 343–348 CrossRef CAS PubMed.
- E. Taqieddin and M. Amiji, Biomaterials, 2004, 25, 1937–1945 CrossRef CAS PubMed.
- S. W. Lee, S. A. Cheon, M. I. Kim and T. J. Park, J. Nanobiotechnol., 2015, 13, 1–10 CrossRef.
- W. R. Hagen, J. Chem. Soc., Dalton Trans., 2006, 37, 4415–4434 RSC.
- L. B. Wang, Y. C. Wang, R. He, A. Zhuang, X. Wang, J. Zeng and J. G. Hou, J. Am. Chem. Soc., 2013, 135, 1272–1275 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c6ra24192a |
| This journal is © The Royal Society of Chemistry 2016 |