
Biosorption and bioaccumulation of chromate from aqueous solution by a newly isolated Bacillus mycoides strain 200AsB1†
 *abc
*abc
Abstract
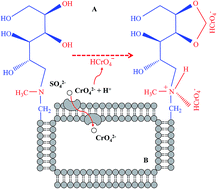
Microbial chromate (Cr6+) reduction and consequent chromite (Cr3+) biosorption have exhibited potential for the remediation of chromium (Cr)-contaminated water. However, few microorganisms that can accumulate amounts of Cr6+ in their cells have been reported. In this study, a new Cr6+-resistant bacterium was isolated and characterized by physiological, biochemical and molecular tests. Its Cr6+ resistant ability, removal efficiency and mechanisms were also investigated at different initial Cr6+ concentrations, solution pHs and incubation temperatures. Results showed that the strain 200AsB1 was a typical bacterium belonging to Bacillus mycoides and tolerated 125 mg L−1 Cr6+ (34 h-LC50 = 63.9 mg L−1). Although the strain preferred to grow at low pH without Cr6+ stress, it only grew well and removed Cr6+ from neutral or alkaline solutions. Similar to the pH experiment, the strain grew better at low temperature than at high temperature, regardless of Cr6+ amendment, but high temperature promoted Cr6+ removal. By analyzing the functional groups' change on the bacterial surface by Fourier transform infrared spectroscopy, our data indicated that N–H and O–H groups from N-methyl-glucamine were involved in Cr6+ adsorption. While Cr concentrations on the bacterial surface were 0.53–0.96 mg L−1, the cell Cr concentration was up to 273 mg kg−1, both of which might contribute to the efficient Cr6+ removal from aqueous solution. Our study demonstrated that B. mycoides strain 200AsB1 could provide new opportunities to remediate Cr-contaminated water through both biosorption and bioaccumulation processes.
 Please wait while we load your content...
Please wait while we load your content...

