
Efficient and microbial communities for pollutant removal in a distributed-inflow biological reactor (DBR) for treating piggery wastewater†
Zhenxing Zhong,
Xiaohui Wu,
Lan Gao,
Xiejuan Lu and
Beiping Zhang*
School of Environmental Sciences and Engineering, Huazhong University of Science and Technology, Wuhan 430074, China. E-mail: probpzhang@126.com; Fax: +86 2787792155; Tel: +86 2787792155
First published on 26th September 2016
Abstract
To treat piggery wastewater with a low C/N ratio, a novel distributed-inflow biological reactor (DBR) was developed that assures the proper organic flow for an efficient denitrification process. The operation of the DBR featured the use of continuous feeding and bypass flow rates (x = 0.2–0.5). Results demonstrated that under low C/N ratio conditions (3.8–5.4), bypass rates of 0.3–0.4 were desirable and resulted in COD, NH4+–N, and TN removal efficiencies of 89.2%, 97.4%, and 86.2%, respectively. Track studies elucidated the removal and transformation of organic matter and nitrogen in the DBR. High-throughput sequencing showed that Proteobacteria and Bacteroidetes were dominant in both the DBR and the sludge obtained from a wastewater treatment plant (WWTP) for seeding the DBR. Compared with the seed sludge, the populations of aerobic heterotrophic bacteria (Filimonas and Chryseolinea) and heterotrophic denitrifiers (Thauera, Halomonas, and Ottowia) were apparently enriched in DBR, and were mainly responsible for the removal of COD and NO3−–N. In contrast, the populations of ammonium-oxidizing bacteria (AOB) and nitrite-oxidizing bacteria (NOB) decreased sharply. Although stable nitrification was achieved, autotrophic nitrifiers such as AOB and NOB were unexpectedly present as minor species, probably due to the synergistic effects of autotrophic AOB and heterotrophic nitrifiers.
1. Introduction
In the past three decades, the increasing demand for meat has resulted in a tremendous increase in pig breeding in many countries such as China. In China at the end of 2010, there were approximately 963![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 pig farms (each producing ≥100 heads per year).1 Correspondingly, development of the pig industry has resulted in a considerable increase in the production and discharge of highly concentrated wastewater. Piggery wastewater is characterized by high concentrations of COD, ammonia, and suspended solids, as well as significant amount of salts and heavy metals, but a low ratio of C/N.2,3 If not treated safely, piggery wastewater can be a major source of serious environmental tribulations.
000 pig farms (each producing ≥100 heads per year).1 Correspondingly, development of the pig industry has resulted in a considerable increase in the production and discharge of highly concentrated wastewater. Piggery wastewater is characterized by high concentrations of COD, ammonia, and suspended solids, as well as significant amount of salts and heavy metals, but a low ratio of C/N.2,3 If not treated safely, piggery wastewater can be a major source of serious environmental tribulations.
To reduce the negative impacts of swine wastewater on the environment, various techniques have been used to treat piggery wastewater prior to discharge. These include the up-flow anaerobic sludge blanket reactor (UASB), internal circulation reactor (IC), anaerobic sequencing batch reactor (ASBR), anaerobic–anoxic–aerobic process (A2/O) and membrane bioreactor (MBR).2,4–7 However, the effluent from anaerobic digestion contains high COD and high nitrogen concentrations, and far exceeds the discharge standards. In addition, the complex control systems and expensive running costs impede the broad application of techniques such as A2/O and MBR. The SBR and modified SBR, as engineered treatment technologies with robust adaptability and small size, have been paid more attention by researchers compared to other techniques.8–10 Because swine wastewater with a low C/N ratio has a small amount of organic matter, which limits the denitrification process, sufficient organic matter (as a carbon source) must be provided for proper denitrification. There are two possible ways to enhance the denitrification process. The first way is to dose the organic matter with external carbon sources, for which glucose, acetate and methanol are often chosen.8,10 Unfortunately, this approach increases the treatment cost sharply. The other approach is to use the piggery wastewater as an electron donor for denitrification in SBR. This technique has been suggested by some researchers and the external carbon source is a viable choice for enhancing the nitrogen removal.9,10 Because any excess external carbon added over the amount required for the denitrification process to proceed efficiently would raise the operational cost unnecessarily, the addition of the external carbon source should be optimized. However, this optimization in SBR remains largely unstudied. Furthermore, SBR and modified SBR are generally operated in an intermittent mode and need a hydraulic retention time (HRT) as long as 9–16 d,8,11 resulting in a large construction investment, high power consumption and a complex control system.12 Cost-effective treatment of swine wastewater therefore remains a great challenge.
Based on the results of previous studies and aiming to optimize the utilization of organic matter, a novel distributed-inflow biological reactor (DBR) composed of two reaction compartments (DBR-1 and DBR-2) was developed to treat piggery wastewater. To ensure enough organic matter for denitrification in the DBR system, a strategy of distributed and continuous feeding was adopted, and aeration and agitation were alternately implemented in the DBR system to promote the nitrification and denitrification processes. Compared with a conventional SBR that operates with intermittent feeding, the DBR could operate with a continuous inflow (and outflow), and directly eliminate the time-consuming process of reactor filling, decanting and idling that characterizes a conventional SBR. Importantly, these changes increase the treatment capability of the DBR beyond what is achievable in a conventional SBR.
Previous research demonstrated that organic matter was removed by heterotrophic bacteria in a biological system, and ammonia and nitrate were eliminated by nitrifiers and denitrifiers,13,14 respectively. However, the precise biological mechanism of these processes during wastewater treatment remains indeterminate due to a lack of detailed information about the microbial community structure in a bioreactor. In addition, bioreactor performance relies on functional bacteria,15 yet information on efficiency and functional bacterial populations in industrial WWTPs is scarce, especially in piggery wastewater treatment systems. Fortunately, high throughput sequencing techniques have become a highly efficient tool for identifying the entire profile of bacterial communities and have been applied extensively to analyze the bacterial communities in various environments.16,17
In this study, a novel DBR was operated to treat piggery wastewater using a strategy of distributed and continuous feeding. The objectives of this research were: (1) to investigate the long-term performance of the DBR system in simultaneously eliminating carbon and nitrogen from piggery wastewater, (2) to optimize the influent bypass rates in the DBR system, and (3) to reveal (using Illumina high throughput sequencing technology) the microbial community structure and the relationship between the key microbial populations and the removal of the pollutants.
2. Materials and methods
2.1. Laboratory equipment, wastewater quality and seeding sludge
Fig. 1 shows the experimental system, which includes a feed tank and the two compartments DBR-1 and DBR-2. DBR-1 and DBR-2 has a working volume of 31.25 L and 30.625 L, respectively. The DBR-2 compartment consists of two zones, i.e., a biological reaction zone and a settling zone. In the biological reaction zone, aeration was achieved using an air pump, and agitation was achieved using a mechanical mixer. The air pump and mixer were operated alternately under the control of an automatic timer. The settling zone was separated from the biological zone by a vertical clapboard. Two layers of guide plates were installed at the lower end of the settling zone for separating the activated sludge from the mixed liquor, and there was a gap (3–5 cm) between the guide plates of each layer. The separated sludge returned to the bottom of the biological zone by moving along the gaps between the guide plates in response to gravity. Then the separated liquid and the non-separated sludge flowed into the inclined plate settler, where the non-separated sludge would be further polished. Notably, the compartment DBR-1 had identical structures as DBR-2, except for the inclined plate settler in the settling zone. | ||
| Fig. 1 Schematic diagram of proposed DBR system. | ||
Because the organic matter of piggery wastewater with a low C/N ratio is generally insufficient to support effective denitrification, a strategy of distributed and continuous feeding was adopted in the DBR system. The influent (Q = 10 mL min−1) was divided into two parts: one part (Q1 = Q*(1 − x)) was fed into the DBR-1 compartment, while the other part (Q2 = Q*x) bypassed DBR-1 and was fed directly into the DBR-2 to provide sufficient organic matter for denitrification. The bypass flow rate (x), which varied from 0.2 to 0.5, was introduced in this study.
Wastewater fed to the DBR was the raw piggery wastewater collected from a commercial pig farm in Wuhan, China. The wastewater was sieved through a 0.8 mm diameter mesh to remove large solids prior to use in the bioreactor. A summary of the sieved wastewater characteristics is given in Table 1. The COD and TN of the wastewater samples used in this study varied from 2220 mg L−1 to 4610 mg L−1 and from 459.1 mg L−1 to 1193.6 mg L−1, respectively. This variation was due to differences in the growth stage of animals, frequency and water volume in the flush, environmental factors and other managerial factors.2 Notably, NH4+–N dominated the TN and the average COD/TN ratio was as low as 4.5.
| Period | Days | Bypass rates (x)a | COD (mg L−1) | TN (mg L−1) | NH4+–N (mg L−1) | COD/TN | OLRb (kg per (m3 per d)) | NLRc (kg per (m3 per d)) | pH |
|---|---|---|---|---|---|---|---|---|---|
| a The inflow ratio (x) of Q2 to Q1 in terms of volume.b Organic loading rate in terms of COD.c Nitrogen loading rate in terms of TN. | |||||||||
| Startup | 0–68 | — | 4070 ± 843 | 893.7 ± 169.4 | 811.3 ± 159.9 | 4.7 ± 1.2 | 0.95 ± 0.20 | 0.21 ± 0.04 | 7.2 ± 0.3 |
| SET 1 | 69–84 | 0.5 | 4610 ± 531 | 977.7 ± 64.0 | 891.7 ± 67.5 | 4.7 ± 0.6 | 1.07 ± 0.12 | 0.23 ± 0.02 | 7.3 ± 0.2 |
| SET 2 | 85–100 | 0.4 | 4754 ± 442 | 1193.6 ± 268.0 | 1021.8 ± 183.8 | 4.2 ± 0.8 | 1.11 ± 0.10 | 0.28 ± 0.06 | 7.4 ± 0.2 |
| SET 3 | 101–116 | 0.3 | 2220 ± 593 | 623.2 ± 235.8 | 564.8 ± 225.7 | 3.8 ± 0.7 | 0.52 ± 0.14 | 0.15 ± 0.06 | 7.2 ± 0.2 |
| SET 4 | 117–132 | 0.2 | 2297 ± 175 | 459.1 ± 19.2 | 405.8 ± 23.0 | 5.0 ± 0.6 | 0.53 ± 0.04 | 0.11 ± 0.01 | 7.3 ± 0.1 |
2.2. Acclimation of the inoculated sludge
The DBR was inoculated with seed sludge collected from Longwangzui Wastewater Treatment Plant (WWTP) (Wuhan, China) that operates with an AAO process. The sludge load rate of the WWTP aerobic tank is 0.16–0.2 kg BOD5 per (kg MLSS per d). After inoculation, the DBR was operated for 68 d, which was divided into two periods. In the first period from day 0 to 21, in order to acclimate the inoculated sludge to the wastewater, piggery wastewater fed into the DBR was diluted using tap water and the dilution ratios were 10, 5, 3, and 1. Following a decline in ammonia and TN removal during the first 8 d, the removal of ammonia and TN increased (although with fluctuation) and became steady. In the second period from day 22 to 68, the piggery wastewater was not diluted and was directly fed into the DBR system. Although the quality of the wastewater fluctuated moderately (Table 1), the elimination of nutrients in the DBR system was relatively constant, and the system achieved COD, NH4+–N, and TN removal efficiencies of 88.5%, 95.8%, and 81.7%, respectively. The TN removal moderately fluctuated, which may have been related to the obvious fluctuation of nitrogen concentration and the low COD/TN ratio of 2.3–7.6 in influent piggery wastewater. The concentrations of mixed liquor suspended solids (MLSS) in the DBR-1 and DBR-2 compartments were 6120–7850 mg L−1 and 3390–4690 mg L−1, respectively. The sludge retention time (SRT) was maintained at approximately 25 d by controlling sludge wastage.2.3. Experimental procedure
After acclimation of the inoculated sludge, the DBR system was continuously operated for approximately two months at a hydraulic retention time (HRT) of 4.3 d. To identify the proper bypass flow rate in the DBR system, four different rates ranging from 0.5 to 0.2 were tested, corresponding to four independent operating conditions, i.e., SET 1, 2, 3 and 4, respectively. The feed and operating parameters of the four SETs are presented in Table 1. The performance of the DBR system was evaluated according to the efficiencies with which COD, NH4+–N, and TN were removed. Based on results from a pre-experiment, all SETs were operated with an identical cyclic period of 480 min, including 240 min for aeration and 240 min without aeration. Air was supplied at the rate of 2 L min−1 by two air compressors through tube-type diffusers installed at the bottom of the DBR. Complete mixing was provided by two mechanical stirrers rotating at a speed of 70 rpm. During the aerobic phase of in the DBR system, the target DO concentration was set for the range 2–4 mg L−1, while the target was 0–0.3 mg L−1 in the anoxic period. The DBR was operated at room temperature (17.1–32.3 °C).Additionally, track studies during one complete cycle of the optimized bypass flow rate were implemented to identify the removal of organic matter and nitrogen in the DBR.
2.4. Analytical methods
During the experiments, samples were regularly collected at various parts of the DBR system (feeding tank, influent, effluent of the DBR-1, and DBR-2). Chemical oxygen demand (COD), total nitrogen (TN), ammonia (NH4+–N), nitrite (NO2−–N), nitrate (NO3−–N), mixed liquid suspended solids (MLSS) and MLVSS (mixed liquor volatile suspended solids) concentrations were analyzed according to standard methods.18 The temperature and DO were monitored using a portable DO meter (3410, WTW Company, Germany), pH was measured using Sartorius pH meter (PB-10, Sartorius Company, Germany), ORP (Oxidation–Reduction Potential) was monitored by Shanghai Rex PHS-3C meter (China).2.5. Bacterial community study
723 (DBR-1), 57024 (DBR-2) and 51951 (WWTP) sequences.3. Results and discussion
3.1. Performance of the DBR system
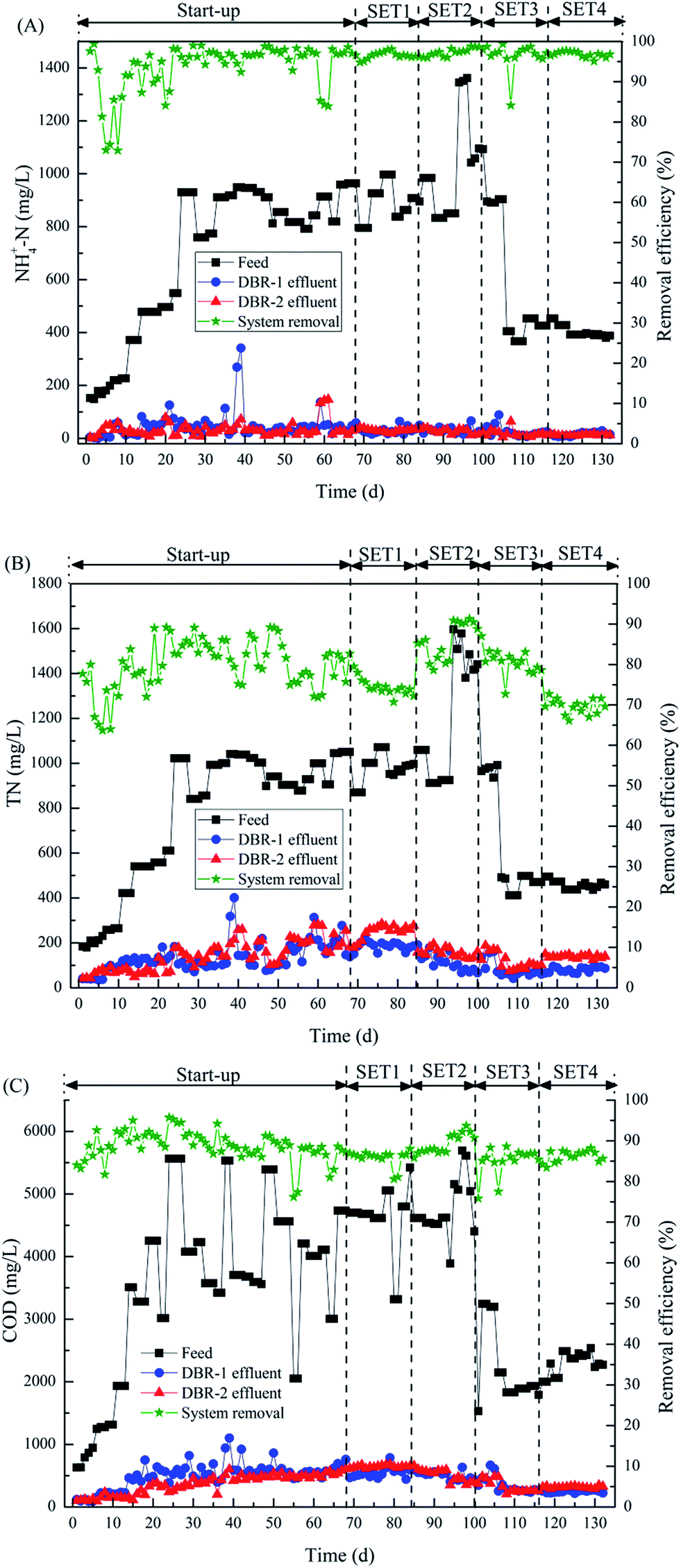 | ||
| Fig. 2 Removal efficiencies of ammonia (A), total nitrogen (B) and COD (C) in DBR system during the experimental period. | ||
Fig. 2(B) shows the temporal variations of influent and effluent TN concentrations in the DBR system. In the four SETs, the TN concentration in the influent fluctuated dramatically in the range 459.1–1193.6 mg L−1, while the average removal of TN was 74.4% (SET 1), 86.2% (SET 2), 80.6% (SET 3) and 69.6% (SET 4). Although TN removal in DBR was good, it was noticeably lower than the ammonia removal and was significantly affected by the bypass flow rate. These results may be attributed to the organic matter distribution in the four SETs due to the different bypass flow rates and the composition of TN in piggery wastewater. As known from previous research and this study, TN was primarily comprised of ammonia (85–92%) and organic nitrogen (8–15%). Ammonia could be transferred into nitrite and nitrate by nitrifiers, but organic nitrogen could not be eliminated by the DBR.2,4 Under the low C/N ratios, the nitrate removal by denitrification was closely related to the carbon source,23,26 hence, the removal of nitrate and TN was apparently influenced by the bypass flow rate. Similar results were obtained in previous studies. Kim et al. found the effluent NOx−–N concentration clearly increased from 107 mg L−1 to 219 mg L−1 as the bypass flow rates decreased from 1 to 0.2.4 Rajagopal et al.5 found that TN removal increased from 66% to 70–73% in A2/O process when the influent bypass rates decreased from 50% to 30–40%. Regarding the high effluent concentration of TN (approximately 100 mg L−1) in the current study, similar results have been typically reported for various biological processes due to non-degradable organic nitrogen and residual nitrates.2,24 It’s reported that a fraction of organic nitrogen was non-biodegradable and could not be eliminated by biological processes.5 Meanwhile, a certain concentration of nitrate (40–219 mg L−1) presented in the effluent of various biological processes due to incomplete denitrification and the low C/N ratios (3.2–10) in piggery wastewater.4,5,24 In SET 1–4, the nitrogen loading rate (NLR) and TN removal rate of the DBR process (Table 2) were 0.11–0.28 kg per (m3 per d) and 0.07–0.24 kg per (m3 per d), respectively, which might be a little low for bacteria in the DBR.
| Period | COD removal (kg per (m3 per d)) | TN removal (kg per (m3 per d)) | CODremoved/TNremoved ratio | COD removal (%) | TN removal (%) | NH4+–N removal (%) |
|---|---|---|---|---|---|---|
| Start-up | 0.85 ± 0.19 | 0.17 ± 0.03 | 5.1 ± 1.4 | 88.7 | 79.5 | 93.7 |
| SET 1 | 0.92 ± 0.12 | 0.17 ± 0.01 | 5.5 ± 0.8 | 85.8 | 74.4 | 96.5 |
| SET 2 | 0.99 ± 0.11 | 0.24 ± 0.07 | 4.3 ± 1.0 | 89.2 | 86.2 | 97.4 |
| SET 3 | 0.44 ± 0.12 | 0.11 ± 0.05 | 4.1 ± 0.9 | 85.1 | 80.6 | 96.4 |
| SET 4 | 0.46 ± 0.04 | 0.07 ± 0.01 | 6.2 ± 0.9 | 86.2 | 69.6 | 96.8 |
The data in Table 2 and Fig. 2(C) show that in all four SETs, despite dramatic fluctuations in the influent COD concentrations (2220–4754 mg L−1), the average COD removal in the DBR was almost constant (85–89%) regardless of the bypass flow rate. This result is similar to that for ammonia removal. A similar COD removal efficiency (83–94%) from swine wastewater was also obtained in other biological processes,5,23 and the residual organic matter in effluent may be inert soluble compounds.2,4,8 Moreover, in all four SETs, although the average organic loading rates were 0.52–1.11 kg COD per m3 per d, the average organic removal rates (ORRs) of the DBR reached 0.44–0.92 kg COD per m3 per d (Table 2). Compared with traditional SBR and the modified SBR, the DBR system in this study presented some incomparable advantages in the aspects of nutrient removal and simplified control systems.
There are some probable reasons for the above results. In SET 2 (x = 0.4), 60% of the influent was fed directly into the DBR-1 (40% was fed into DBR-2), and the influent ammonia was converted to nitrite and nitrate during nitrification. However, the nitrite and nitrate were not completely removed by denitrification in the subsequent non-aeration period because the carbon source was insufficient.5 Instead, any remaining nitrite and nitrate were further polished in DBR-2 where 40% of the influent organics were continuously fed in the flow that bypassed DBR-1. In SET 3 (x = 0.3), the TN removal is slightly decreased compared to that in SET 2. The possible reason for this result was that the available carbon source correspondingly decreased with the decreased bypass rate (from 0.4 to 0.3). In SET 1, however, the highest bypass rate (x = 0.5) was used and 50% of influent was continuously fed into the DBR-2. Thus, the HRT of the influent in DBR-2 was relatively short (compared to that in SET 2 and SET 3) due to the continuous inflow and outflow of the higher quantity of wastewater. As a consequence, a fraction of TN was directly discharged from DBR-2 before it could be removed, causing the effluent TN concentration of DBR-2 to increase rather obviously. In addition, the average effluent COD in SET 1 (approximately 645 mg L−1) was the highest among all SETs, which also confirmed that a fraction of organic matter and nitrogen in the influent were directly discharged before processes within the reactor could remove them. Rajagopal et al.5 developed a process that combined anaerobic digestion with an anoxic/oxic system to treat piggery wastewater. These researchers found that the TN removal was only 66% when 50% of the influent was bypassed into the anoxic/aerobic reactor, while the TN elimination increased to 70–73% when the bypass flow rates decreased to 30–40%, which is in agreement with the results achieved in the current study.
Moreover, the removal of TN in SET 4 (x = 0.2) was clearly lower than that in SET 2 and SET 3. The probable explanation for this result is that only 20% of the influent organics were fed into the DBR-2 and the corresponding carbon source was grossly insufficient to support effective denitrification. As a consequence, NO3−–N and TN concentrations in the effluent from DBR-2 increased sharply compared to those in SET 2 and SET 3. Besides, the MLSS of DBR-2 was remarkably reduced from approximately 4690 mg L−1 to 3390 mg L−1 in SET 4, indicating that the carbon source in DBR-2 was insufficient for the growth and metabolism of heterotrophic bacteria. Additionally, data in Fig. 2(B) showed that the effluent TN of DBR-2 was sometimes slightly higher than that of DBR-1, which indicated that the non-biodegradable nitrogen in the effluent of DBR-2 was probably higher than that of DBR-1. Finally, though the TN removal was enhanced by the distributed-inflow mode, and excellent removal of nitrogen was achieved in the DBR process, a certain amount of TN (approximately 150 mg L−1) remained in the effluent from DBR-2 due to the presence of non-biodegradable nitrogen and the low COD/TN ratio in swine wastewater.2,4,5 These results illustrated that the bypass flow rates ranging from 0.3 to 0.4 could be regarded as optimal for the DBR process, which promoted the efficacious removal of TN in piggery wastewater.
3.2. Track study of the SET 2 (x = 0.4) in DBR
To identify the real-time variations of COD and nitrogen in DBR, track studies of SET 2 and SET 3 were conducted during a full operational cycle of the reactor. Fig. 3 represents the typical profiles of nitrogen, organic matter, ORP, DO and pH during one complete cycle of SET 2. As shown in Fig. 3(A), the average COD concentration in DBR-1 was approximately 554 mg L−1 and the COD remained stable during one complete cycle. There are two primary reasons for this phenomenon. Firstly, most of the organic matter in swine wastewater was readily biodegradable and easily removed by aerobic heterotrophic bacteria in the aeration phase of the DBR operation.2,8 During the non-aeration period, the organic matter was mainly utilized by heterotrophic denitrifiers, as suggested by the low CODremoved/TNremoved ratios of 4.1–6.2 (Table 2).5 Secondly, the effective removal of COD may have been related to the low organic loading rate. The sludge loading rates in DBR-1 were about 0.06–0.21 kg COD per kg MLSS per d, which might be a little low for the heterotrophic bacteria to consume. | ||
| Fig. 3 Variations of nitrogen, organic, DO, pH and ORP during one typical cycle of DBR-1 (A) and DBR-2 (B) in SET 2 (x = 0.4); (A) ORP break point, (B) ammonia valley point, (C) DO break point. | ||
Similar results were also observed in DBR-2 of SET 2 and in SET 3. As shown in Fig. 3(B) and S1(B),† the average COD concentrations in DBR-2 were approximately 573 mg L−1 and 269 mg L−1, respectively. These results demonstrated that the effluent COD of DBR may be closely related to the initial concentration of organic material and the inert organic matter in the piggery wastewater as aforementioned. Similar or higher concentrations of COD in the effluent were also found by previous researchers and attributed to the recalcitrant compounds.2,4,24,25
In the nitrification process within DBR-1, ammonia was oxidized to nitrite and then nitrate by nitrifiers (Fig. 3(A)). Then nitrite and nitrate were converted to nitrogen gas by heterotrophic denitrifiers in the denitrification process. Notably, the NO2−–N concentration during nitrification reached 48.4 mg L−1, which implied a high ammonia content in the influent may be favorable to achieving partial nitrification. During the nitrification and denitrification processes, several characteristic points were identified, namely the “ammonium valley”, the “DO breakout point”, and the “ORP break point”, which appeared at the end of nitrification.10,24 However, both the “nitrite knee” and the “nitrite apex” disappeared during the denitrification process. This perhaps was due to the insufficient amount of organic matter and the low ratio of COD/TN in the influent.9 At the end of the non-aeration phase, the presence of a fraction of nitrite and nitrate in the effluent of DBT-1 also verified that the phenomena was present in this study.
During the initial phase of aeration, the pH consistently declined due to the nitrification process and the consumption of the alkalinity. Conversely, the pH consistently increased in the rest of the cycle because nitrification was completed and the alkalinity was consistently increased due to the continuous feed of raw influent. As for DBR-1, although the pH initially decreased (from 8.3 to 7.9) due to nitrification and then rose to approximately 8.4, the pH in this compartment during a typical operation cycle of the DBR was relatively steady and suitable for nitrifiers; furthermore, the effluent ammonia concentration remained stable. Similar results were also obtained in DBR-2. However, Kishida et al., observed a sharp decrease in the pH and activity of nitrifying bacteria in an SBR used to treat swine wastewater (with a BOD5/TN ratio of 2.6) and noted that the effluent ammonia concentration increased and eventually exceeded 200 mg L−1.24 Additionally, with respect to the DBR-2 of SET 2 (x = 0.4), the profiles of physical parameters, organic matter and nitrogen were similar to those obtained in DBR-1 (Fig. 3(B)).
The track studies of DBR-1 and DBR-2 during SET 3 (x = 0.3) are plotted in Fig. S1.† These data show that the nitrite concentrations were less than 10 mg L−1 and 1.6 mg L−1 in DBR-1 and DBR-2, respectively, and that the COD varied between 244 and 301 mg L−1 during an entire cycle. The results may be attributed to the relatively low influent concentrations of ammonia and COD in SET 3. The variation in other parameters and nitrogen were similar to those observed in SET 2 (x = 0.4). Finally, it should be noted that track analysis is available to reveal the removal and transformation of organic and nitrogen in DBR, optimize the operational conditions of nitrification and denitrification, prevent the occurrence of excess aeration, enhance the nitrogen removal and reduce energy cost.
3.3. Overall analysis of Illumina Miseq sequencing
Three 16S rRNA gene libraries were constructed from Illumina high-throughput sequencing of microbial communities in seed sludge (WWTP), DBR-1 and DBR-2, for which 52723, 57024 and 51951 high quality sequence tags, respectively, were identified and 2964 (WWTP), 2127 (DBR-1) and 2411 (DBR-2) operational taxonomic units (OTUs) were clustered at a 3% distance. The total number of OTUs estimated by the Chao1 estimator was 6383 (WWTP), 4477 (DBR-1) and 5304 (DBR-2) at a 3% distance (Table 3), indicating that the microbial species in the three samples were very rich. These results also confirmed that the full-scale WWTP fed with municipal wastewater exhibited greater microbial richness than the lab-scale DBR system fed with piggery wastewater. Similar results have been verified by previous research and surveys.27–30 The Shannon diversity index denotes not only the species richness but also the species abundance in a microbial community. Considering that the Shannon diversity index of DBR-1 (4.43/4.29) was lower than WWTP (5.84/5.57) and DBR-2 (5.10/4.89), it could be inferred that the enriched OTUs in WWTP and DBR-2 communities were distributed more evenly than those in DBR-1. The value of Good’s coverage indicated that all samples were sequenced at high depth (Table 3).
| Sample | Sequence | Reads | α = 0.03 | α = 0.05 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| OTU | Chao1 | Shannon | Coverage | OTU | Chao1 | Shannon | Coverage | |||
| WWTP | 52723 |
43693 |
2847 | 5923 | 5.84 | 0.96 | 2107 | 3988 | 5.57 | 0.98 |
| DBR-1 | 57024 |
47065 |
1945 | 4056 | 4.43 | 0.97 | 1458 | 2896 | 4.29 | 0.98 |
| DBR-2 | 51951 |
40070 |
2411 | 5304 | 5.10 | 0.97 | 1602 | 3084 | 4.89 | 0.98 |
3.4. Microbial distribution analysis in the DBR system
 | ||
| Fig. 4 Abundances of different phyla in samples from WWTP and DBR systems (A), and a Venn diagram of the bacterial communities of WWTP, DBR-1 and DBR-2 based on OTUs at 3% distance (B). | ||
In all three communities, the sum of total observed OTUs was 6213, but only 135 OTUs (2.2%) were shared by the communities (Fig. 4(B)). Notably, DBR-1 and DBR-2 shared more OTUs (994, 16.0% of total) with each other than either of them shared with WWTP (DBR-1/WWTP, 202, 3.3%; DBR-2/WWTP, 228, 3.7%). Moreover, the majority (72.1%) of the shared OTUs of DBR-1 and DBR-2 were Proteobacteria and Bacteroidetes, accounting for 52.0% and 20.1%, respectively, of the total observed sequences. The number of OTUs unique to an individual community was 2669 (WWTP), 1066 (DBR-1) and 1324 (DBR-2), accounting for 81% of the total number of observed OTUs.
The order level identification of the bacterial communities in three samples is plotted in Fig. 5. The dominant orders in DBR-1 and DBR-2 were Sphingobacteriales, Burkholderiales, Alteromonadales, and Flavobacteriales, while WWTP primarily consisted of Burkholderiales, Rhodocyclales, Nitrosomonadales, Sphingobacteriales, and Planctomycetales. These results show close similarity between the bacterial composition of samples from DBR-1 and DBR-2. However, a few differences between DBR-1 and DBR-2 were also observed, which is possibly due to the different influent. The influent of DBR-1 was only piggery wastewater (Q1) while the influent of DBR-2 consisted of piggery wastewater (Q2) and the effluent from DBR-1. Earlier analyses of activated sludge performed in full-scale and lab-scale WWTPs indicated that wastewater characteristics made the greatest contribution to the bacterial community variance, and that bioreactor scale, water temperature, DO, pH, SRT, and MLSS also made contributions.27,32,33
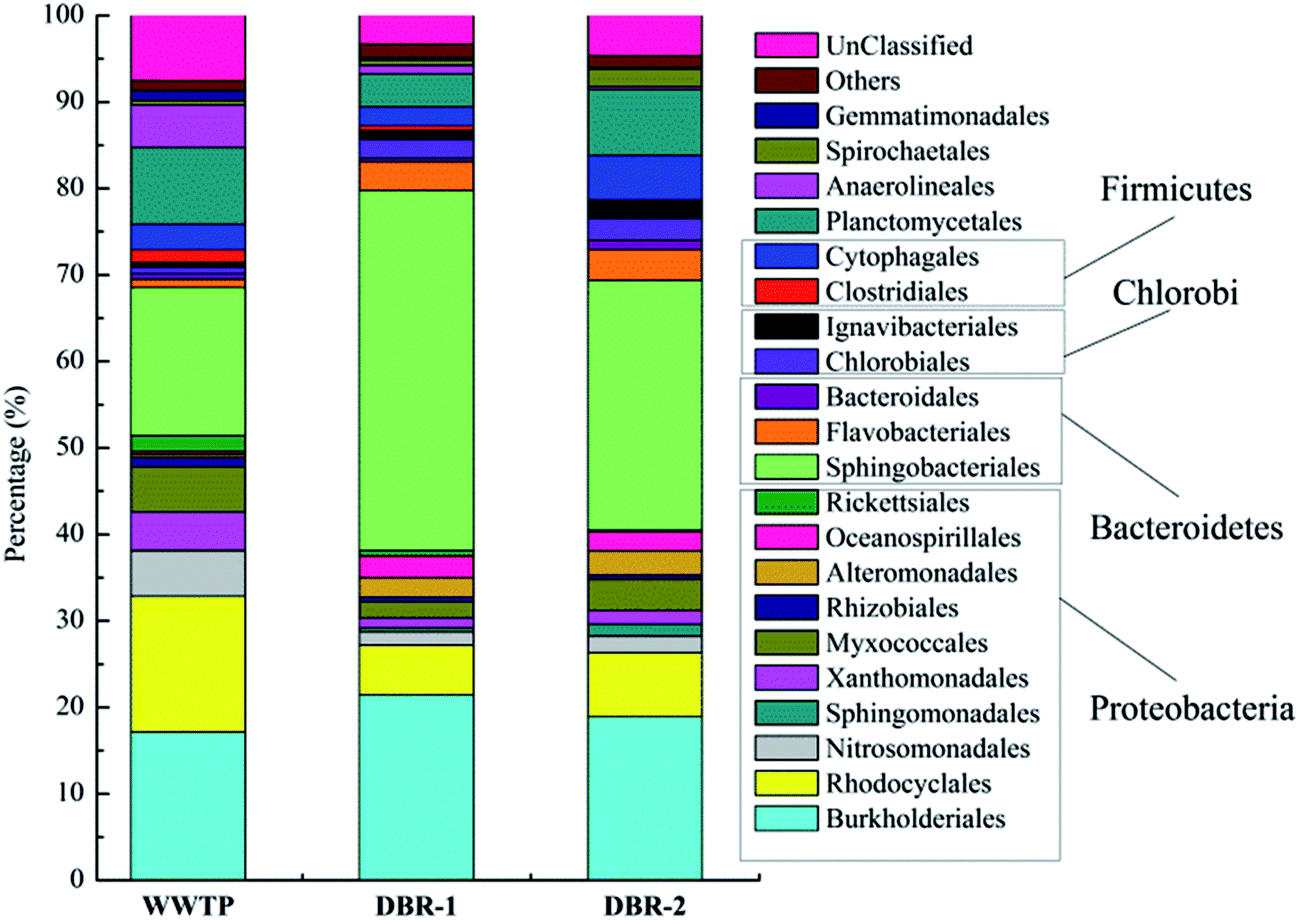 | ||
| Fig. 5 The relative abundances of the predominant phylogenetic groups of WWTP, DBR-1 and DBR-2 at the order level. | ||
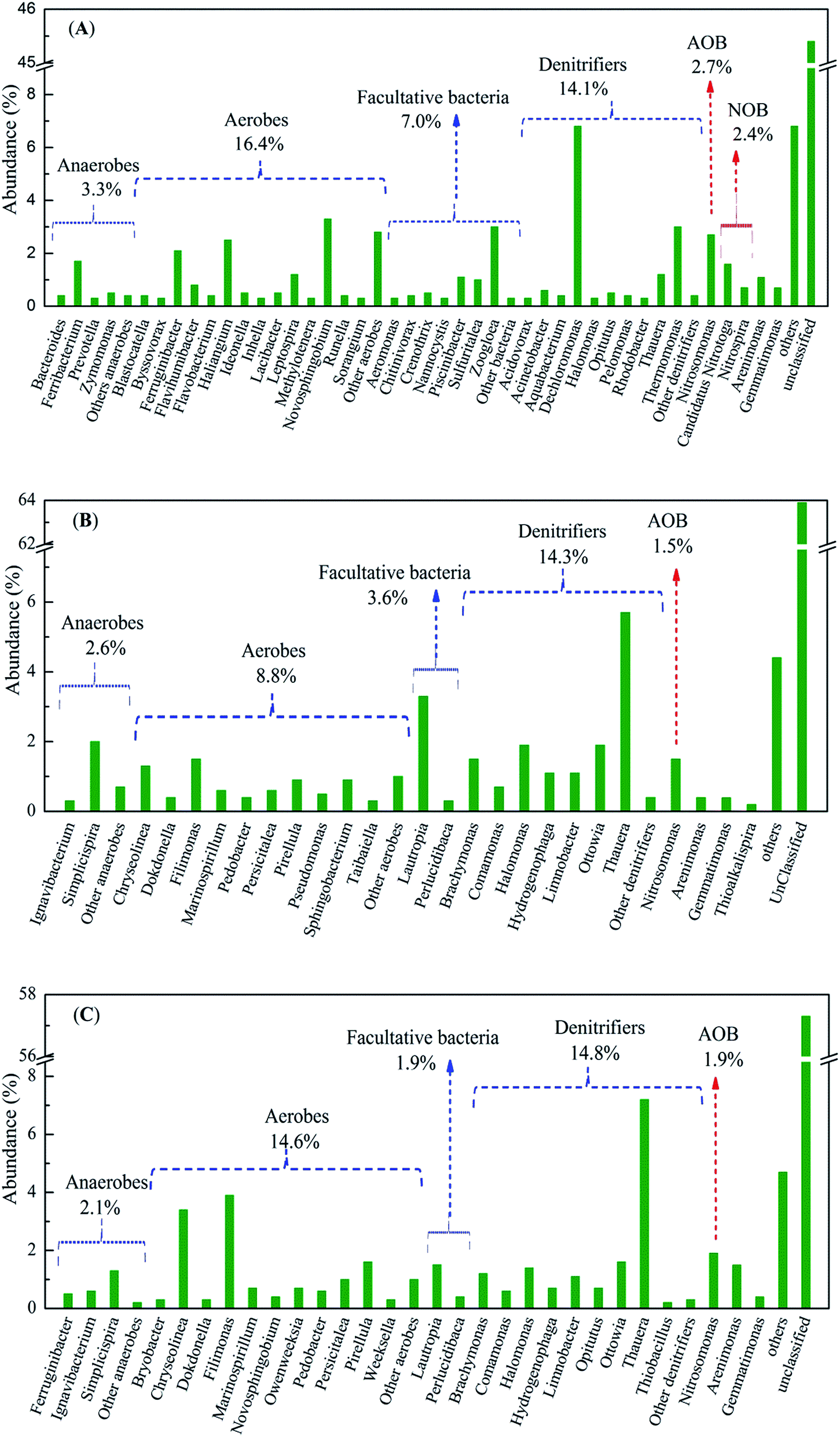 | ||
| Fig. 6 The relative abundances of the functional bacterial populations of WWTP, DBR-1 and DBR-2 at the genus level. | ||
Aerobic heterotrophic bacteria, which were mainly responsible for COD removal in the aeration phase of the DBR process, were the foremost populations in WWTP (16.4%), but were less abundant in DBR-1 (8.8%) and DBR-2 (14.6%). Novosphingobium (3.3%), Ferruginibacter (2.1%) and Flavihumibacter (0.8%), all versatile organic degraders for carbohydrates and other organics, were the major genera in WWTP (Fig. 6(A)).36,37 However, as in DBR-1 and DBR-2, Chryseolinea, Filimonas, and Persicitalea were the primary aerobic heterotrophic genera and exhibited a relatively high abundance (0.6–3.9%). These genera were responsible for the process of oxidization and the utilization of diverse mono- and disaccharides as well as some polysaccharides and a few organic acids, and may have taken part in the removal of organic matter in piggery wastewater.38–40 Meanwhile, some other genera (such as Pirellula and Marinospirillum) that were absent from WWTP were enriched in the DBR system. This result indicated that these genera grew well in DBR fed with swine wastewater. Furthermore, some facultative bacteria only dominated in certain samples, such as Piscinibacter (1.1%, WWTP), Zoogloea (3.0%, WWTP), and Lautropia (3.3% in DBR-1, and 1.5% in DBR-2). The heterotrophic growth of these bacteria occurs on saccharides such as galactose, sucrose agar, and propionic acid under aerobic conditions, while the bacteria can also ferment various carbohydrates and types of macromolecular organic matter under anaerobic conditions.37,41,42 Enrichment of these aerobic and facultative genera presumably contributed to the efficacious removal of organic matter.43,44
Nitrosomonas was the dominant AOB in all three samples (Fig. 6), but its abundance in WWTP (2.7%) was significantly greater than in DBR-1 (1.5%) and DBR-2 (1.9%). Likewise, Nitrospira (0.7%) and Candidatus Nitrotoga (1.6%) were the dominant species of NOB in WWTP, but were not detected in the DBR system. These results indicated that AOB and NOB may not be resistant to external changes. Ma et al.45 demonstrated that the relative abundances of Nitrosomonas and Nitrospira decreased dramatically with an increase in aeration intensity. Based on the competition for nitrite between heterotrophic bacteria and NOB, Jenni and co-workers proved that NOB activity decreased steadily as the influent C/N ratio increased from 0.2 to 1.4.46 That NOB disappeared in the DBR probably due to the influent C/N ratios of 3.8–5.0 and the external changes. On the other hand, the unexpectedly low percentage of AOB and NOB in DBR seemed to contradict the successful nitrification that was achieved. However, a similar phenomenon was also observed in earlier studies. Figuerola and Erijman could hardly detected any AOB in an oil refinery industrial WWTP that achieved high nitrification efficiency.47 Zhao et al.,48 found that at high COD and nitrogen loading rates, heterotrophic and autotrophic nitrification coexisted and that the heterotrophic nitrification contributed more to nitrogen removal in a lab-scale aerobic granule sludge reactor that successfully treated modified piggery wastewater. Based on the results of previous studies and this study, nitrification is considered to be the result of synergistic effects of AOB and NOB, autotrophic and heterotrophic bacteria.49,50 Furthermore, Thauera and Comamonas (both promising denitrifiers) were also proven to harbor perfect nitrification capacity in industrial wastewater treatment.48–50 In the DBR system, a high concentration of organic matter can favor potential heterotrophic nitrifiers. Hence, the relative abundance of Thauera and Comamonas reached up to 5.7% and 0.7%, respectively, in DBR-1, and 7.2% and 0.6% in DBR-2, respectively. These microorganisms may out-compete autotrophic bacteria and contribute more to ammonia elimination from swine wastewater. However, the nitrification contribution of heterotrophic nitrifiers needs further research.
Heterotrophic denitrifiers, which are mainly responsible for nitrogen removal, were the foremost populations in the DBR system (Fig. 6(B) and C). Dechloromonas (6.8%), Thermomonas (3.0%), and Thauera (1.2%) and represent genera that potentially contribute to the denitrification process in WWTP.17 However, except for Thauera and Halomonas, the relative abundance of other denitrifiers sharply decreased in the DBR compared to the WWTP. As for the DBR, Thauera (5.7–7.2%), Halomonas (1.4–1.9%), and Ottowia (1.6–1.9%) are functionally important denitrifiers. The abundance of these microorganisms implied that a selective enrichment of the three genera in DBR might be due to their flexibility in piggery wastewater. Earlier surveys pointed out that Thauera was the most abundant potential denitrifier in 17 nitrogen removal plants and nine coking WWTPs.50,51 Halomonas and Ottowia, which also participated in denitrification, were detected in industrial WWTPs.30,46 Furthermore, although the dominant denitrifiers were identical in DBR-1 and DBR-2, each biological reaction zone contained some unique populations. For instance, the three denitrifiers Thiobacillus (0.2%), Aquabacterium (0.1%) and Azoarcus (0.1%) were identified only in DBR-2, while Steroidobacter (0.1%) was only detected in DBR-1. Notably, autotrophic denitrifiers (Thiobacillus) were observed in DBR-2 with a proportion of 0.2%; importantly, this population can accomplish denitrification without the need for a supplemental carbon source.36 Members of these lineages seemed to be tolerant and well-adapted to piggery wastewater as evidenced by their increased occurrence upon exposure to this complex wastewater. In summary, the enriched abundance of a diverse denitrifying community enabled the DBR process to achieve satisfactory TN removal.
In this study, using distributed and continuous feeding, both DBR-1 and DBR-2 alternately worked in aerobic and anoxic phases. Based on the Illumina high-throughput sequencing analyses, there was a clear distinction in the compositions and structures of the bacterial communities in the DBR and WWTP, despite the fact that sludge from the WWTP was used to seed the DBR. High relative abundance of aerobes, facultative bacteria and heterotrophic denitrifiers coexisted and cooperated to eliminate pollutants during the DBR process. Chryseolinea, Filimonas, and Persicitalea were the primary aerobic heterotrophic genera for removing COD in the DBR. Thauera, Halomonas, and Ottowia are functionally important denitrifiers for the removal of organic matter and nitrate in the anoxic phase of the DBR process. Moreover, a high influent concentration of ammonia was removed effectively, probably by the synergistic effects of autotrophic AOB and heterotrophic nitrifiers (Comamonas and Thauera) in the biological system. Additionally, some functional genera, such as polyphosphate accumulating bacteria (PAOs) (0.4–0.7% Gemmatimonas) and desulfurizing bacteria (0.4–1.5% Arenimonas) were also found in three sludge samples.
4. Conclusions
In this study, a novel DBR was developed for the simultaneous removal of organic matter and nitrogen from piggery wastewater with a low C/N ratio. The DBR was operated using a mode of continuous and distributed feeding. Results demonstrated that bypass flow rates of 0.3–0.4 were optimal for facilitating the elimination of nutrients in the DBR. The average COD, NH4+–N, and TN removal reached 89.2%, 97.4%, and 86.2%, respectively. Track studies elucidated the removal and transformation of organic matter and nitrogen in the DBR. High-throughput sequencing showed that both Proteobacteria and Bacteroidetes were dominant in the DBR and WWTP. Compared with the seed sludge in WWTP, populations of aerobic heterotrophic bacteria (Filimonas and Chryseolinea) and heterotrophic denitrifiers (Thauera, Halomonas, and Ottowia) were enriched in DBR, whereas the populations of AOB and NOB decreased sharply. The stable and high nitrification efficiency in DBR may be related to the synergistic effects of autotrophic AOB, NOB and heterotrophic nitrifiers (Comamonas and Thauera).Acknowledgements
This research was supported by the National High Technology Research and Development Program of China (863 Program) (Grant No. 2012AA06A304). The authors would like to thank Dr Liu ZQ, Dr Zhou T, Dr Mao J (HUST) for their scientific contribution and Dr Wang LL, Dr Chen J (HUST) for their technical contribution in this study.References
- H. Gao, China animal industry yearbook, Chinese agricultural press, Beijing, 2011 Search PubMed.
- N. Prado, J. Ochoa and A. Amrane, Water Res., 2009, 43, 1549–1558 CrossRef CAS PubMed.
- P. Tulayakul, A. Boonsoongnern, S. Kasemsuwan, S. Wiriyarampa, J. Pankumnoed, S. Tippayaluck, H. Hananantachai, R. Mingkhwan, R. Netvichian and S. Khaodhiar, J. Environ. Sci., 2011, 23, 991–997 CrossRef CAS.
- D. Kim, E. Choi, Z. Yun and S. Kim, Water Sci. Technol., 2004, 49, 165–171 CAS.
- R. Rajagopal, P. Rousseau, N. Bernet and F. Beline, Bioresour. Technol., 2011, 102, 2185–2192 CrossRef CAS PubMed.
- L.-W. Deng, P. Zheng and Z.-A. Chen, Process Biochem., 2006, 41, 965–969 CrossRef CAS.
- P. Li, Y. Wang, Y. Wang, K. Liu and L. Tong, Water Sci. Technol., 2010, 61, 243–252 CrossRef CAS PubMed.
- G. Bortone, S. Gemelli, A. Rambaldi and A. Tilche, Water Sci. Technol., 1992, 26, 977–985 CAS.
- Z. Han, W. Wu, J. Zhu and Y. Chen, Biosystems Eng., 2008, 99, 273–281 CrossRef.
- J. H. Kim, M. Chen, N. Kishida and R. Sudo, Water Res., 2004, 38, 3340–3348 CrossRef CAS PubMed.
- A. Tilche, E. Bacilieri, G. Bortone, F. Malaspina, S. Piccinini and L. Stante, Water Sci. Technol., 1999, 40, 199–206 CrossRef CAS.
- S. Piccinini, F. Verzellesi and P. Mantovi, Biological nutrient removal in a full sequencing batch reactor treating pig slurry, Proceedings of the 10th International Conference of the RAMIRAN Network, Strbske Pleso, High Tatras, Slovak Republic, 2002, pp. 391–395 Search PubMed.
- L. Ye, M. F. Shao, T. Zhang, A. H. Tong and S. Lok, Water Res., 2011, 45, 4390–4398 CrossRef CAS PubMed.
- S. Yang, W. Guo, Y. Chen, X. Zhou, H. Zheng, X. Feng, R. Yin and N. Ren, RSC Adv., 2014, 4, 52892–52897 RSC.
- W. Zhi, Z. Ge, Z. He and H. Zhang, Bioresour. Technol., 2014, 171, 461–468 CrossRef CAS PubMed.
- Y. Yang, K. Yu, Y. Xia, F. T. Lau, D. T. Tang, W. C. Fung, H. H. Fang and T. Zhang, Appl. Microbiol. Biotechnol., 2014, 98, 5709–5718 CrossRef CAS PubMed.
- T. Zhang, M. F. Shao and L. Ye, ISME J., 2012, 6, 1137–1147 CrossRef CAS PubMed.
- APHA, American Public Health Association (APHA): Washington, DC, USA, 2005.
- G. C. Baker, J. J. Smith and D. A. Cowan, J. Microbiol. Methods, 2003, 55, 541–555 CrossRef CAS PubMed.
- Z. Liu, T. Z. DeSantis, G. L. Andersen and R. Knight, Nucleic Acids Res., 2008, 36, e120 CrossRef PubMed.
- Y. Wang and P.-Y. Qian, PLoS One, 2009, 4(10), e7401 Search PubMed.
- MEPPRC, MEPPRC (Ministry Environmental Protection of People’s Republic of China), Discharge Standard of Pollutants for Livestock and Poultry Breeding, Beijing, CHN, 2001 Search PubMed.
- L. Deng, P. Zheng, Z. Chen and Q. Mahmood, Bioresour. Technol., 2008, 99, 3136–3145 CrossRef CAS PubMed.
- N. Kishida, J.-H. Kim, M. Chen, H. Sasaki and R. Sudo, J. Biosci. Bioeng., 2003, 96, 285–290 CrossRef CAS PubMed.
- Z. Han, W. Wu, Y. Ding, J. Zhu and Y. Chen, J. Environ. Sci. Health, Part A: Toxic/Hazard. Subst. Environ. Eng., 2008, 43, 161–170 CrossRef CAS PubMed.
- D. Obaja, S. Mace, J. Costa, C. Sans and J. Mata-Alvarez, Bioresour. Technol., 2003, 87, 103–111 CrossRef CAS PubMed.
- R. Hai, Y. Wang, X. Wang, Y. Li and Z. Du, PLoS One, 2014, 9, e90175 Search PubMed.
- C. J. van der Gast, B. Jefferson, E. Reid, T. Robinson, M. J. Bailey, S. J. Judd and I. P. Thompson, Environ. Microbiol., 2006, 8, 1048–1055 CrossRef PubMed.
- P. W. Ethan, B. A. Peter, K. L. William, A. G. Richard, G. David, M. K. Dawn, R. Andrew, A. R. James, D. S. Melinda and R. S. John, Oikos, 2006, 112, 185–195 CrossRef.
- F. M. Ibarbalz, E. L. Figuerola and L. Erijman, Water Res., 2013, 47, 3854–3864 CrossRef CAS PubMed.
- M. Wagner and A. Loy, Curr. Opin. Biotechnol., 2002, 13, 218–227 CrossRef CAS PubMed.
- G. F. Wells, H. D. Park, B. Eggleston, C. A. Francis and C. S. Criddle, Water Res., 2011, 45, 5476–5488 CrossRef CAS PubMed.
- Q. Niu, T. Kobayashi, Y. Takemura, K. Kubota and Y. Y. Li, Bioresour. Technol., 2015, 193, 110–118 CrossRef CAS PubMed.
- S. Lu, S. H. Ryu, B. S. Chung, Y. R. Chung, W. Park and C. O. Jeon, Int. J. Syst. Evol. Microbiol., 2007, 57, 31–34 CrossRef CAS PubMed.
- T. Iino, K. Mori, Y. Uchino, T. Nakagawa, S. Harayama and K. Suzuki, Int. J. Syst. Evol. Microbiol., 2010, 60, 1376–1382 CrossRef CAS PubMed.
- J. Meng, J. Li, J. Li, K. Sun, P. Antwi, K. Deng, C. Wang and G. Buelna, Bioresour. Technol., 2016, 201, 166–173 CrossRef CAS PubMed.
- G. Garrity, J. T. Staley, D. R. Boone, P. De Vos, M. Goodfellow, F. A. Rainey, K.-H. Schleifer, D. J. Brenner and N. R. Krieg, Bergey’s Manual® of Systematic Bacteriology: Volume Two: The Proteobacteria, Springer Science & Business Media, 2006 Search PubMed.
- J. J. Kim, M. Alkawally, A. L. Brady, W. I. Rijpstra, J. S. Sinninghe Damste and P. F. Dunfield, Int. J. Syst. Evol. Microbiol., 2013, 63, 654–660 CrossRef CAS PubMed.
- H. Shiratori, Y. Tagami, T. Morishita, Y. Kamihara, T. Beppu and K. Ueda, Int. J. Syst. Evol. Microbiol., 2009, 59, 1137–1142 CrossRef CAS PubMed.
- J. Yoon, S. Ishikawa, H. Kasai and A. Yokota, Int. J. Syst. Evol. Microbiol., 2007, 57, 1014–1017 CrossRef CAS PubMed.
- Y. Shao, B. S. Chung, S. S. Lee, W. Park, S. S. Lee and C. O. Jeon, Int. J. Syst. Evol. Microbiol., 2009, 59, 526–530 CrossRef CAS PubMed.
- E. Stackebrandt, S. Verbarg, A. Fruhling, H. J. Busse and B. J. Tindall, Int. J. Syst. Evol. Microbiol., 2009, 59, 2552–2560 CrossRef CAS PubMed.
- K. Nath and D. Das, Appl. Microbiol. Biotechnol., 2004, 65, 520–529 CrossRef CAS PubMed.
- J. A. Field, A. J. M. Stams, M. Kato and G. Schraa, Antonie Van Leeuwenhoek, 1995, 67, 47–77 CrossRef CAS PubMed.
- J. Ma, Z. Wang, Y. Yang, X. Mei and Z. Wu, Water Res., 2013, 47, 859–869 CrossRef CAS PubMed.
- S. Jenni, S. E. Vlaeminck, E. Morgenroth and K. M. Udert, Water Res., 2014, 49, 316–326 CrossRef CAS PubMed.
- E. L. Figuerola and L. Erijman, Environ. Microbiol., 2007, 9, 1780–1789 CrossRef CAS PubMed.
- Y. Zhao, J. Huang, H. Zhao and H. Yang, Bioresour. Technol., 2013, 143, 439–446 CrossRef CAS PubMed.
- T. Felfoldi, A. J. Szekely, R. Goral, K. Barkacs, G. Scheirich, J. Andras, A. Racz and K. Marialigeti, Bioresour. Technol., 2010, 101, 3406–3414 CrossRef CAS PubMed.
- Q. Ma, Y. Qu, W. Shen, Z. Zhang, J. Wang, Z. Liu, D. Li, H. Li and J. Zhou, Bioresour. Technol., 2015, 179, 436–443 CrossRef CAS PubMed.
- T. R. Thomsen, Y. Kong and P. H. Nielsen, FEMS Microbiol. Ecol., 2007, 60, 370–382 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Fig. S1. See DOI: 10.1039/c6ra20777d |
| This journal is © The Royal Society of Chemistry 2016 |