
Efficient access to L-phenylglycine using a newly identified amino acid dehydrogenase from Bacillus clausii†
Jun Cheng,
Guochao Xu,
Ruizhi Han,
Jinjun Dong and
Ye Ni*
The Key Laboratory of Industrial Biotechnology, Ministry of Education, School of Biotechnology, Jiangnan University, Wuxi 214122, Jiangsu, China. E-mail: yni@jiangnan.edu.cn
First published on 16th August 2016
Abstract
An amino acid dehydrogenase from Bacillus clausii (BcAADH) was identified and overexpressed in Escherichia coli BL21(DE3) for the preparation of L-phenylglycine from benzoylformic acid. Recombinant BcAADH was purified to homogeneity and characterized. BcAADH could catalyse reductive amination and oxidative deamination at optimum pHs of 9.5 and 10.5. Furthermore, BcAADH has a broad substrate spectrum, displaying activities toward various aromatic and aliphatic keto acids. When coexpressed with glucose dehydrogenase from Bacillus megaterium, the potential application of BcAADH in the preparation of L-phenylglycine was investigated at a high substrate loading and low biocatalyst addition. As much as 400 mM benzoylformic acid could be fully reduced into L-phenylglycine within 6 h at >99.9% ee. With merely 0.5 g DCW L−1, 200 mM benzoylformic acid was completely reduced, resulting in a substrate to biocatalyst ratio of 60 g g−1, environmental factor of 4.7 and 91.7% isolation yield at gram scale. This study provides guidance for the application of BcAADH in the synthesis of chiral non-natural amino acids.
Introduction
Optically active non-natural amino acids are important compounds, and are widely used as vital building blocks for pharmaceuticals and agrochemicals.1–4 Due to their multifunctional structure, non-natural amino acids could also be applied in the synthesis of structural motifs of peptides and peptidomimetics.5–7 There has been an increasing demand for the non-natural amino acids, especially L-phenylglycine.8 Ascribing this to their various functional groups (aromatic, carboxylic and chiral amine groups), chiral L-phenylglycine and derivatives are important blocks of antibiotics such as penicillin,9 pristinamycin I,10 and the antitumor Taxol,11 antiplatelet inhibitors Clopidogrel.12Various chemical routes have been developed for the synthesis of non-natural amino acids using asymmetric reactions. However, the utilization of environmentally harmful organic solvents and toxic reagents such as cyanides are disadvantageous for industrial production.13,14 Moreover, the relatively low enantioselectivity could not satisfy the optical purity requirement for pharmaceuticals.15 In comparison, biological methods, using microbes and enantioselective enzymes, have also been established and are preferable for the production of non-natural amino acid.15,16 Through metabolic engineering and pathway recombination, L-phenylalanine and L-phenylglycine could be produced through fermentation.17 However, the metabolic pathways is complex which might influence the normal growth of microbes, and the product L-phenylglycine is hard to be recovered from natural α-amino acids.18,19 Enzymatic synthesis approach is a green alternative for the preparation of optically active non-natural amino acids considering its environmental benignity and high enantioselectivity.20 Amino acid dehydrogenases are regarded as a group of enzymes capable of producing amino acids. Catalytic tandem reaction participated by various biocatalysts in one pot is an elegant way for the synthesis of enantiomeric pure compounds.21,22 Asymmetric synthesis of chiral alcohols, amines and amino acids, as well as non-natural amino acid have been successfully accomplished using these multi-enzymatic cascades.23–27 A three-enzyme cascade reaction has been developed by Fan and coworkers employing D-mandelate dehydrogenase from Lactobacillus brevis, mandelate racemase from Pseudomonas and leucine dehydrogenase from Exiguobacterium sibircum for the synthesis of L-phenylglycine.24 Under optimized condition, 0.2 M mandelic acid was converted into L-phenylglycine, with 96.4% conversion and >99% ee. However, this bioprocess requires large loading of biocatalysts. Consequently, there is a constant demand for enzymes with high efficiency, stability and enantioselectivity for scale-up production of amino acids.28–30
In this study, a novel leucine dehydrogenase (BcAADH) was identified and cloned from Bacillus clausii, with 50.6% identity to phenylalanine dehydrogenase from Rhodococcus sp. Recombinant BcAADH was purified and the enzymatic properties such as optimum temperature and pH, substrate specificity and kinetic parameters were studied. To our knowledge, few AADHs have been applied in the preparation of L-phenylglycine. The potential of this newly identified BcAADH in the synthesis of L-phenylglycine was also investigated. Our results suggest that BcAADH could not only tolerate high substrate loading, but also display high substrate to biocatalyst ratio.
Results and discussion
Genome mining for amino acid dehydrogenases
To find efficient aryl amino acid dehydrogenases (AADHs), genome data mining was adopted. Four potential AADHs were selected from genome database using the amino acid sequence of phenylalanine dehydrogenase from Rhodococcus sp. (PheDH, Q59771.2) as probe. After heterogeneously expressed in E. coli BL21(DE3), the reductive amination activity of AADHs toward benzoylformic acid was investigated. One AADH from Bacillus clausii NRRL B-23342 (BcAADH) under accession no. Q5WF72 was proved to be efficient in the asymmetric reduction reaction of benzoylformic acid. This newly identified AADH displays 50.6% identity with the amino acid sequence of PheDH. Sequence and phylogenetic analysis reveal that BcAADH belongs to the Glu/Leu/Phe/Val dehydrogenase family and displays high similarity with leucine dehydrogenase from Exiguobacterium sibiricum31 and valine dehydrogenase from Bacillus badius33 as shown in Fig. 1. Alignment of BcAADH and other amino acid dehydrogenases from different origins indicate that conserved residue and domains were also found in BcAADH (Fig. S1†), including catalytic K80 and NADH binding domain of GVGNVAY (180–186). | ||
| Fig. 1 Phylogenetic analysis of BcAADH and other amino acid dehydrogenases. | ||
Purification and characterization of BcAADH
Recombinant BcAADH with N-terminal His-taq was purified by nickel affinity chromatography. After desalting and concentration, purified BcAADH was obtained and analyzed by SDS-PAGE. As shown in Fig. 2, BcAADH was migrated a single band at about 45 kDa, in agreement with its theoretical value. In addition, gel exclusion chromatography revealed that the molecular weight was 90 kDa, indicating BcAADH is a homodimeric enzyme consisting of two identical subunits. Specific activity of purified BcAADH toward benzoylformic acid was 17.7 U mg−1, 2.1 folds higher than that of the crude extract.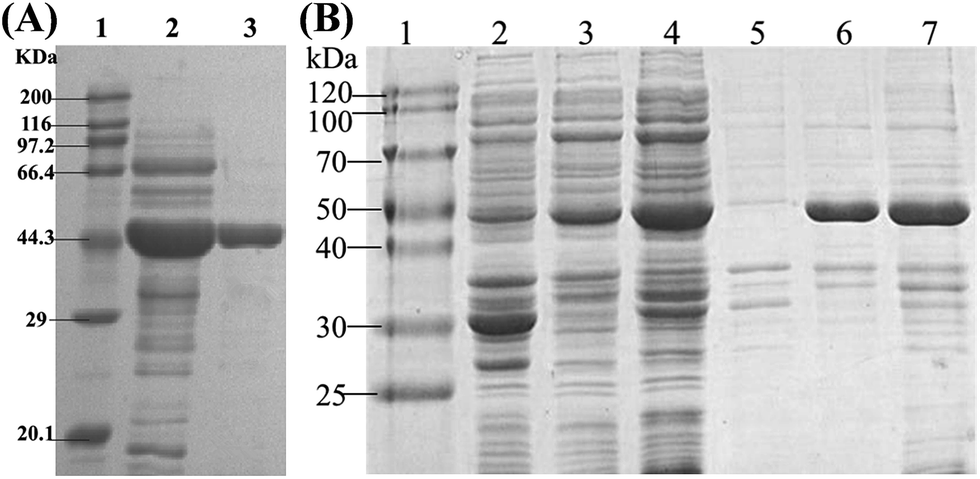 | ||
| Fig. 2 SDS-PAGE of analysis of purified BcAADH and coexpression of BcAADH and BmGDH. (A) Lane 1: protein molecular marker, lane 2: crude extract, lane 3: purified BcAADH; (B) lane 1: protein molecular marker, lanes 2 & 5, 3 & 6, 4 & 7: supernatant and precipitant of recombinant E. coli BL21(DE3) harboring pACYCDuet-BmGDH, pET28-BcAADH and pACYCDuet-BmGDH/pET28-BcAADH. | ||
Enzyme activities at different temperatures of 20–50 °C were investigated. As illustrated in Fig. 3A, BcAADH shows the highest activity at around 30 °C. Half-lives of BcAADH were 315, 85.6 and 4.5 h at 30, 40 and 50 °C (Fig. S2†), respectively, indicating BcAADH was stable than at operational temperature.31 The half-lives of amino acid dehydrogenases from Sporosarcina psychrophila DSM3 and Exiguobacterium sibircum ECU9271 at 50 °C were 0.5 h and 11 h.31,33 Effect of pH ranging from 7.5 to 11.0 on the reductive amination and oxidative deamination activities of BcAADH were also investigated (Fig. 3B). For reductive amination, the most suitable pH was pH 9.5. While for the oxidative deamination, BcAADH displays the maximum activity at pH 10.5. Competing reductive and oxidative activities of BcAADH indicate the dual-function of amino acid dehydrogenases in the synthesis of amino acids.
 | ||
| Fig. 3 Effects of temperature and pH on the activity of BcAADH. (A) Temperature profile of BcAADH. (B) pH profiles of the reductive and oxidative activities of BcAADH, blue line: reductive profile, green line: oxidative profile. | ||
Effects of various metal ions on the activity of BcAADH were also studied (Table S3†). The enzyme activity was obviously inhibited by the addition of 1 mM Li+, Fe3+ and Ag+, while Mg2+ and Co+ could promote the activity of BcAADH to 108.3% and 113.5% respectively. EDTA is a metal ions chelator, which could eliminate the metal ions from enzyme. As a result, the metal ion dependent enzyme could be deactivated or lose some activity. However, EDTA had none influence on the activity of BcAADH. Consequently, BcAADH is presumed to be a metal ion-independent amino acid dehydrogenase.
Substrate spectrum of BcAADH toward various keto acids and L-amino acids with different substituents was investigated. As shown in Table 1, BcAADH preferred aliphatic to aromatic keto acids in the asymmetrically reductive amination reaction. Moreover, most of the tested aromatic keto acids could be reduced by BcAADH, especially benzoylformic acid (17.7 U mg−1). The highest activity was observed with 4-methyl-2-oxopentanoic acid (96.1 U mg−1), which could be reduced into L-leucine by BcAADH. For the oxidative deamination, BcAADH could also catalyse the oxidative deamination reaction of aromatic amino acids, such as L-phenylglycine (0.11 U mg−1). However, higher activity was found with aliphatic amino acids, especially L-leucine (0.59 U mg−1), which might be ascribed to its lower steric hindrance and similarity to leucine dehydrogenase from Bacillus sphaericus ATCC4525.37 Our results indicate BcAADH has a wide substrate spectrum.
| Reductive amination | Oxidative deamination | ||
|---|---|---|---|
| Substrate | Relative activity [%] | Substrate | Relative activity [%] |
| a Specific activity toward benzoylformic acid was regarded as 100%, 17.7 U mg−1.b Specific activity toward L-phenylglycine was regarded as 100%, 0.11 U mg−1. | |||
 |
100a | 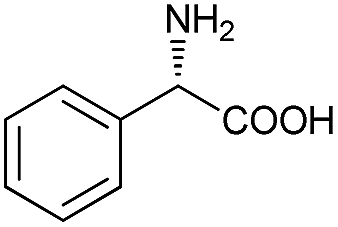 |
100b |
 |
5.88 |  |
<0.1 |
 |
<0.1 |  |
346 |
 |
0.23 |  |
318 |
 |
543 | 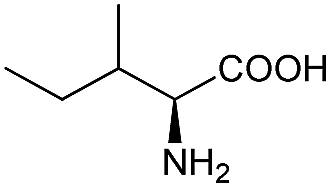 |
536 |
 |
322 |  |
<0.1 |
 |
2.86 | ||
 |
8.96 | ||
Kinetic parameters of purified BcAADH toward several preferred substrates were investigated (Table 2). The Km and kcat of BcAADH toward aromatic amino acids, benzoylformic acid and 2-chlorobenzoylformic acid, were 13 mM and 65.2 s−1, 6.33 mM and 1.56 s−1, respectively. BcAADH displays the highest affinity and catalytic efficiency on 4-methyl-2-oxopentanic acid (2.74 mM and 242 s−1). However, with regard to the substrates with smaller side-chains, such as pyruvic acid and trimethylpyruvic acid, poor binding affinity was observed, resulting in low kcat/Km (0.088 s−1 and 0.423 s−1 respectively), which were similar to other amino acid dehydrogenases, such as EsAADH.31
| Substrate | Km [mM] | kcat [s−1] | kcat/Km [s−1 mM−1] |
|---|---|---|---|
| Benzoylformic acid | 13.0 | 65.2 | 5.02 |
| 2-Chlorobenzoylformic acid | 6.33 | 1.56 | 0.246 |
| 4-Methyl-2-oxopentanoic acid | 2.74 | 242 | 88.3 |
| 2-Ketobutyric acid | 13.9 | 153 | 11.0 |
| Pyruvic acid | 37.2 | 3.27 | 0.088 |
| Trimethylpyruvic acid | 25.7 | 10.9 | 0.423 |
| NADH | 0.373 | 100 | 268 |
Optimization of the asymmetric preparation of L-phenylglycine
To evaluate the potential of BcAADH in the asymmetric preparation of L-phenylglycine, the reaction condition was optimized. Firstly, to drive the reaction, cofactor regeneration system was usually introduced to generate NADH for the reduction reaction. Glucose dehydrogenase (GDH) is usually used as cofactor regeneration system in the asymmetric bioreduction, due to its high stability and activity, low side reaction, low price and favorable thermodynamic.38,39 By the reduction of NAD+ along with the oxidation of glucose, NADH and gluconic acid are produced. Among various GDHs, the GDH from Bacillus megaterium (BmGDH) displays excellent activity and high soluble expression level in E. coli BL21(DE3).32 Consequently, BmGDH was chosen for the regeneration of NADH. BmGDH was coexpressed with BcAADH in E. coli BL21(DE3) harboring both plasmids pET28-BcLeuDH and pACYCDuet-BmGDH. As illustrated in Fig. 2B, GDH and BcAADH were successfully coexpressed in E. coli BL21(DE3). Activities of BcAADH and BmGDH in dry cells were determined to be 5.53 and 3.84 U mg−1 DCW, respectively.To establish an efficient biocatalytic process, various influential factors, including temperature, substrate/biocatalyst loading and NAD+ addition, were investigated. The reaction was optimized to improve the substrate loading and productivity as shown in entries 1–6 in Table 3. Within 0.5 h, 50 mM (7.5 g L−1) benzoylformic acid could be fully reduced into L-phenylglycine using 10 g L−1 biocatalyst (dry cells) at 30 °C and 120 rpm with substrate to biocatalyst ratio of 0.75 g g−1 and >99.9% ee. Under the same condition, when substrate loading was increased to 200 mM (30 g L−1), the conversion could only reach 88.9% even after 24 h (entry 3). Additionally, much L-phenylglycine was crystallized and attached to the reactor. Lower temperature is therefore presumed to be more suitable for the biocatalytic reaction, due to the higher stability of the enzymes. At 25 °C, 200 mM benzoylformic acid could be fully reduced. To test the maximum efficacy of BcAADH, the substrate loading was further increased to 400 mM (60 g L−1). Although 400 mM substrate could be completely reduced in 6 h under assistance of 0.3 mM NAD+, too much L-phenylglycine crystals were attached to the reactor, leading to poor mass transfer. Consequently, further optimization was conducted aiming at improving the efficiency of BcAADH and substrate to biocatalyst ratio (S/B) as shown in entries 7–11 of Table 3. Under 5 g L−1 biocatalyst, 200 mM benzoylformic acid could not be fully reduced even after 12 h, with 92.7% conversion. Addition of 0.3 mM NAD+ could however accelerate the reaction, and 200 mM substrate could be reduced within 1 h (entry 8). The amount of biocatalyst was further reduced to 3, 1 and 0.5 g L−1, the substrate could be fully reduced within 2, 6 and 12 h respectively (entries 9–11). The S/B reached as high as 60 g g−1 with >99.9% ee, ranking the highest level for the bioreductive preparation of L-phenylglycine.34 Environmental factor (E factor) was calculated to be 4.7, demonstrating a green bioprocess catalyzed by BcAADH.35 Enzyme consumption number (e.c.n.) was calculated to be 0.018, illustrating an extremely low amount of enzyme used in the bioprocess.36 In addition, the crystallized product L-phenylglycine could easily be isolated by adjusting the pH to dissolve the crystals, followed by boiling to remove the cells due to the low biocatalyst loading, and then subjected to rotary evaporation. To the best of our knowledge, this BcAADH catalyzed bioprocess for L-phenylglycine is ranking one of most efficient bioprocess, considering its high substrate loading and biocatalysts utilization efficiency.24,40 Therefore, this newly mined BcAADH is highly potential in the asymmetric preparation of L-phenylglycine.
|
||||||||
|---|---|---|---|---|---|---|---|---|
| Entry | Substrate [g L−1] | Biocatalyst [g L−1] | S/Ba [g g−1] | Temperature [°C] | NAD+ [mM] | Time [h] | Conversionb [%] | eec [%] |
| a S/B: substrate to biocatalyst ratio, g g−1.b Conversion was analysed by HPLC equipped with Diamonsil C18 column at 220 nm and flow rate of 0.6 mL min−1 with 5% methanol, 95% KH2PO4 and 0.08% trifluoroacetic acid as mobile phase.c ee was determined by HPLC equipped with Chirobiotic T column at 220 nm and flow rate of 0.5 mL min−1 with 20% methanol, 80% ddH2O and 0.1% TFA as mobile phase. | ||||||||
| 1 | 7.5 | 10 | 0.75 | 30 | 0 | 0.5 | >99 | >99.9 |
| 2 | 15 | 10 | 1.5 | 30 | 0 | 2 | >99 | >99.9 |
| 3 | 30 | 10 | 3 | 30 | 0 | 24 | 88.9 | >99.9 |
| 4 | 30 | 10 | 3 | 25 | 0 | 4 | >99 | >99.9 |
| 5 | 60 | 10 | 6 | 25 | 0 | 24 | 62.8 | >99.9 |
| 6 | 60 | 10 | 6 | 25 | 0.3 | 6 | >99 | >99.9 |
| 7 | 30 | 5 | 6 | 25 | 0 | 12 | 92.7 | >99.9 |
| 8 | 30 | 5 | 6 | 25 | 0.3 | 1 | >99 | >99.9 |
| 9 | 30 | 3 | 10 | 25 | 0.3 | 2 | >99 | >99.9 |
| 10 | 30 | 1 | 30 | 25 | 0.3 | 6 | >99 | >99.9 |
| 11 | 30 | 0.5 | 60 | 25 | 0.3 | 12 | >99 | >99.9 |

Preparation of L-phenylglycine at gram scale
A gram-scale preparation of L-phenylglycine was conducted using the recombinant whole cells of E. coli BL21(DE3)/pET28-BcAADH/pACYCDuet-BmGDH. In a 100 mL reaction system, 3.0 g benzoylformic acid, 50 mg DCW of whole cells, 0.3 mM NAD+ (final concentration) and 7.2 g glucose were added. The reaction was performed at 25 °C with 60 rpm mechanical agitation. The conversion of benzoylformic acid reached 99% in 12 h. After rotary evaporation and crystallization, 2.73 g optically pure L-phenylglycine was harvested, with 91% isolation yield and >99.9% ee. The product was also confirmed to be L-phenylglycine by LC-MS, 1H-NMR and 13C-NMR as follows (Fig. S3–S5†): 1H-NMR (400 MHz, D2O), δ/ppm: 4.36 (s, 1H), 7.16–7.42 (m, 5H); 13C-NMR (100 MHz, D2O), δ/ppm: 60.5126.8, 127.5, 128.8, 181.0.Experimental
Strains and chemicals
Escherichia coli DH5α and BL21(DE3) in this study were preserved in our laboratory and used as the cloning and expression hosts respectively. Plasmids pET28a and pACYCDuet-1 were commercially obtained from Novagen (Madison, WI, USA). Benzoylformic acid, D/L-phenylglycine and other substrates were all purchased from Shanghai CIVI Chemical Technology co. Ltd (shanghai, China).Cloning, expression and purification of BcAADH
Gene coding for BcAADH was amplified from the genomic DNA of Bacillus clausii NRRL B-23342 by PCR using primers (F: CGC![[G with combining low line]](https://www.rsc.org/images/entities/char_0047_0332.gif)
![[A with combining low line]](https://www.rsc.org/images/entities/char_0041_0332.gif)
![[T with combining low line]](https://www.rsc.org/images/entities/char_0054_0332.gif)
![[C with combining low line]](https://www.rsc.org/images/entities/char_0043_0332.gif) ATGGAATTATTTGCAAAGA, R: CCGTTACTTTTTCCTCGAA) with BamHI and XhoI restriction sites. PCR product and pET28a were double digested with BamHI and XhoI. The resultant DNA fragments were recovered and ligated at 4 °C for 12 h. Then the constructed plasmid pET28-BcAADH was transformed into E. coli DH5α. After verified by colony PCR, the recombinant plasmid was extracted and further transferred into E. coli BL21(DE3).
ATGGAATTATTTGCAAAGA, R: CCGTTACTTTTTCCTCGAA) with BamHI and XhoI restriction sites. PCR product and pET28a were double digested with BamHI and XhoI. The resultant DNA fragments were recovered and ligated at 4 °C for 12 h. Then the constructed plasmid pET28-BcAADH was transformed into E. coli DH5α. After verified by colony PCR, the recombinant plasmid was extracted and further transferred into E. coli BL21(DE3).
Recombinant strain E. coli BL21(DE3)/pET28-BcAADH cell was cultivated in 30 mL LB medium containing 50 μg mL−1 kanamycin at 37 °C and 180 rpm, until optical density at 600 nm reached 0.6–0.8. Then 0.1 mM IPTG was added and the recombinant strain was further cultivated at 25 °C for 7 hours. Cells were collected by centrifuging at 8000 × g for 10 min. Then the cells were disrupted by a ATS engineering AH-BASIC II homogenizer (Shanghai, China). And the recombinant BcAADH with N terminal his tag was purified as previously described.31 Purified BcAADH was added with 30% (v/v) glycerol after desalting and concentration, and subsequently stored at −80 °C.
Activity assays
The reductive amination and oxidative deamination activities of BcAADH were determined by monitoring the change of NADH at 340 nm (ε = 6220 L M−1 cm−1) using spectrophotometer. For the reductive amination activity, 200 μL reaction mixture was consisted of 0.5 mM NADH, 5 mM benzoylformic acid and appropriate amount of BcAADH in NH4Cl–NH3·H2O (pH 9.5, 0.5 M). For the oxidative deamination activity, 200 μL reaction mixture was consisted of 0.5 mM NAD+, 5 mM L-phenylglycine and appropriate amount of BcAADH in NH4Cl–NH3·H2O (pH 9.5, 0.5 M). One unit of AADH or glucose dehydrogenase (GDH) was defined as the amount of enzyme that catalyzes the reduction of phenylglycine or oxidation of glucose to produce 1.0 μmol NAD+ or 1.0 μmol NADH per minute. Activity of GDH was defined and determined as previously reported.32 All the assay was performed in triplicate.Enzymatic characterization of purified BcAADH
Co-expression of BcAADH and GDH
Glucose dehydrogenase coding gene was cloned from Bacillus megaterium and inserted into the BamHI and SalI restriction sites of pACYCDuet using ClonExpressII one step clone kit (Vazyme Inc., Nanjing). The resultant plasmids pACYCDuet-BmGDH and pET28-BcAADH were simultaneously transformed into E. coli BL21(DE3) cells and spread on LB plates supplemented with kanamycin and chloramphenicol. E. coli BL21(DE3) harboring pET28-BcAADH and pACYCDuet-BmGDH was identified by colony PCR. Recombinant coexpression cells were cultured and induced as mentioned above. The cells was lyophilized to dry cells and stored at 4 °C for further use.Optimization of the asymmetrically reductive amination of benzoylformic acid
Optimization of the asymmetric amination of benzoylformic acid was conducted in a 20 mL reactor and magnetically stirred at 120 rpm and 30 °C. Firstly, benzoylformic acid was dissolved by addition of diluted NaOH (0.5 M), then NH4Cl and glucose were added into the reactor. The pH of reaction mixture was adjusted to 8.5 with 0.5 M NH3·H2O. Finally the lyophilized cells of the recombinant E. coli BL21(DE3) harboring pET28-BcAADH and pACYCDuet-BmGDH were added to start the reaction. Amount of enzyme, reaction temperature and NAD+ were varied as shown in Table 3. Samples were withdrawn from the reaction mixture to determine the conversion and enantiomeric excess (ee) by HPLC analysis. Conversion was monitored by HPLC equipping with Diamonsil C18 column (250 mm × 4.6 mm, ID 5 mm, DIMKA), detecting at UV 220 nm and 30 °C in the mobile phase including methanol 5%, KH2PO4 95% and trifluoroacetic acid 0.08% at a flow rate of 0.6 mL min−1. The ee of the L-phenylglycine was measured by HPLC with a chiral Astec column (Chirobiotic T, 150 mm × Φ 4.6 mm) at 220 nm, using mobile phase consisted of methanol, ddH2O and TFA with a volume ratio of 20![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :80:0.001. The flow rate was 0.5 mL min−1. Retention times of L- and D-phenylglycine were 6.1 min and 8.6 min, respectively.
:80:0.001. The flow rate was 0.5 mL min−1. Retention times of L- and D-phenylglycine were 6.1 min and 8.6 min, respectively.
Gram-scale synthesis of L-phenylglycine
In a 100 mL reaction system, 3.0 g benzoylformic acid was dissolved in deionized water and adjusted to pH 8.5 with 0.5 M NH3·H2O, then 2.675 g NH4Cl, 0.19 g NAD+, 7.2 g glucose and 0.05 g dry cells were added. The reaction pH was maintained at 8.5 by titrating 0.5 M NH3·H2O. The reaction was conducted at 120 rpm and 30 °C. Samples were withdrawn to analyze the conversion and ee as described above. Finally, reaction mixture was centrifuged (8000 × g, 15 minutes). The precipitation was dissolved with 1.0 M NH3·H2O and subsequently boiled for 10 min to remove the biocatalysts. The product was collected by rotary evaporation and crystallization. The synthesized L-phenylglycine was verified by LC/MS, 1H-NMR and 13C-NMR.Conclusions
In summary, a novel amino acid dehydrogenase (BcAADH) was identified from Bacillus clausii NRRL B-23342 with high activity and enantioselectivity toward aromatic keto acids. BcAADH belongs to Glu/Leu/Phe/Val dehydrogenase family and could catalyse reductive amination and oxidative deamination. The optimum pH for reduction and oxidation were 9.5 and 10.5. Specific activities of BcAADH toward benzoylformic acid and L-phenylglycine were 17.7 and 0.11 U mg−1. The Km and kcat were 13 mM and 65.2 s−1 toward benzoylformic acid. BcAADH displays high efficiency in the asymmetric preparation of L-phenylglycine. When coexpressed with BmGDH from Bacillus megaterium, 200 mM benzoylformic acid could be fully reduced using merely 0.5 g L−1 dry cells, with 91% isolation yield and >99.9% ee. The substrate to biocatalyst ratio and Environmental factor were 60 g g−1 and 4.7. Our results indicate that BcAADH is a highly potential and robust enzyme for the industrial production of valuable non-natural amino acids.Acknowledgements
We are grateful to National Natural Science Foundation of China (21276112, 21506073), Natural Science Foundation of Jiangsu Province (BK20150003), the Fundamental Research Funds for the Central Universities (JUSRP51409B), the Program of Introducing Talents of Discipline to Universities (111-2-06), and a project funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions for the financial support of this research.Notes and references
- S. J. Zuend, M. P. Coughlin, M. P. Lalonde and E. N. Jacobsen, Nature, 2009, 461, 968–970 CrossRef CAS PubMed
.
- T. Lazar, ChemBioChem, 2005, 6, 1127–1128 CrossRef CAS
- S. C. Stinson, Chem. Eng. News, 2001, 79, 79–97 Search PubMed
- J. Altenbuchner, M. Siemann-Herzberg and C. Syldatk, Curr. Opin. Biotechnol., 2001, 12, 559–563 CrossRef CAS PubMed
- S. Servi, D. Tessaro and G. Pedrocchi-Fantoni, Coord. Chem. Rev., 2008, 252, 715–726 CrossRef CAS
- T. K. Oost, C. Sun, R. C. Armstrong, A. S. Al-Assaad, S. F. Betz, T. L. Deckwerth, H. Ding, S. W. Elmore, R. P. Meadows, E. T. Olejniczak, A. Oleksijew, T. Oltersdorf, S. H. Rosenberg, A. R. Shoemaker, K. J. Tomaselli, H. Zou and S. W. Fesik, J. Med. Chem., 2004, 47, 4417–4426 CrossRef CAS PubMed
- R. Williams, Synthesis of optically active R-amino acids, Pergamon Press, Oxford, 1989 Search PubMed
- A. Perdih and M. Sollner Dolenc, Curr. Org. Chem., 2007, 11, 801–832 CrossRef CAS
- L. M. V. Langen, F. V. Rantwijk, V. K. Švedas and R. A. Sheldon, Tetrahedron: Asymmetry, 2000, 11, 1077–1083 CrossRef
- Y. Zhao, R. Feng, G. Zheng, J. Tian, L. Ruan, M. Ge, W. Jiang and Y. Lu, J. Bacteriol., 2015, 197, 2062–2071 CrossRef CAS PubMed
- R. Croteau, R. E. B. Ketchum, R. M. Long, R. Kaspera and M. R. Wildung, Phytochem. Rev., 2006, 5, 75–97 CrossRef CAS PubMed
- M. W. van der Meijden, M. Leeman, E. Gelens, W. L. Noorduim, H. Meekes, W. J. P. van Enckevort, B. Kaptein, E. Vieg and R. M. Kellogg, Org. Process Res. Dev., 2009, 13, 1195–1198 CrossRef CAS
- D. J. Ager, Curr. Opin. Drug Discovery Dev., 2002, 5, 892–905 CAS
- M. A. Wegman, M. H. A. Janssen, F. van Rantwijk and R. A. Sheldon, Adv. Synth. Catal., 2001, 343, 559–576 CrossRef CAS
- J. B. Wang and M. T. Reetz, Nat. Chem., 2015, 7, 948–949 CrossRef CAS PubMed
- D. J. Ager and I. G. Fotheringham, Curr. Opin. Drug Discovery Dev., 2001, 4, 800–807 CAS
- P. P. Taylor, D. P. Pantalenone, R. F. Senkpeil and I. G. Fotheringham, Trends Biotechnol., 1998, 16, 412–418 CrossRef CAS PubMed
- R. McKenna and D. R. Nielsen, Metab. Eng., 2011, 13, 544–554 CrossRef CAS PubMed
- S. P. Liu, R. X. Liu, A. A. EI-Rotail, Z. Y. Ding, Z. H. Gu, L. Zhang and G. Y. Shi, J. Biotechnol., 2014, 186, 91–97 CrossRef CAS PubMed
- B. K. Hubbard, M. G. Thomas and C. T. Walsh, Chem. Biol., 2000, 7, 931–942 CrossRef CAS PubMed
- F. G. Mutti, T. Knaus, N. S. Scrutton, M. Breuer and N. J. Turner, Science, 2015, 349, 1525–1529 CrossRef CAS PubMed
- A. S. Goldman, A. H. Roy, Z. Huang, R. Ahuja, W. Schinski and M. Brookhart, Science, 2006, 312, 257–261 CrossRef CAS PubMed
- D. Enders, M. R. Hüttl, C. Grondal and G. Raabe, Nature, 2006, 441, 861–863 CrossRef CAS PubMed
- C. W. Fan, G. C. Xu, B. D. Ma, Y. P. Bai, J. Zhang and J. H. Xu, J. Biotechnol., 2015, 195, 67–71 CrossRef CAS PubMed
- V. Resch, W. M. F. Fabian and W. Kroutil, Adv. Synth. Catal., 2010, 352, 993–997 CrossRef CAS
- J. H. Sattler, M. Fuchs, K. Tauber, F. G. Mutti, K. Faber, J. Pfeffer, T. Haas and W. Kroutil, Angew. Chem., Int. Ed., 2012, 51, 9156–9159 CrossRef CAS PubMed
- M. Hall and A. S. Bommarius, Chem. Rev., 2011, 111, 4088–4110 CrossRef CAS PubMed
- T. Rojanarata, D. Isarangkul, S. Wiyakrutta, V. Meevootisom and J. M. Woodley, Biocatal. Biotransform., 2004, 22, 195–201 CrossRef CAS
- W. M. Liu, H. M. Ma, J. X. Luo, W. H. Shen, X. Xu, S. Li, Y. Hu and H. Huang, Biochem. Eng. J., 2014, 91, 204–209 CrossRef CAS
- T. Ema, S. Ide, N. Okita and T. Sakai, Adv. Synth. Catal., 2008, 350, 2039–2044 CrossRef CAS
- J. Li, J. Pan, J. Zhang and J. H. Xu, J. Mol. Catal. B: Enzym., 2014, 105, 11–17 CrossRef CAS
- G. C. Xu, H. L. Yu, Y. P. Shang and J. H. Xu, RSC Adv., 2015, 5, 22703–22711 RSC
- Y. Zhao, T. Wakamatsu, K. Doi, H. Sakuraba and T. Ohshima, J. Mol. Catal. B: Enzym., 2012, 83, 65–72 CrossRef CAS
- G. W. Zheng, J. Pan, H. L. Yu, M. T. Ngo-Thi, C. X. Li and J. H. Xu, J. Biotechnol., 2010, 150, 108–114 CrossRef CAS PubMed
- C. Wandrey, A. Liese and D. Kihumbu, Org. Process Res. Dev., 2000, 4, 286–290 CrossRef CAS
- R. A. Sheldon, Green Chem., 2007, 9, 1273–1283 RSC
- H. M. Li, D. M. Zhu, B. A. Hyatt, F. M. Malik, E. R. Biehl and L. Hua, Appl. Biochem. Biotechnol., 2009, 158, 343–351 CrossRef CAS PubMed
- J. C. Moore, D. J. Pollard, B. Kosjek and P. N. Devine, Acc. Chem. Res., 2007, 40, 1412–1419 CrossRef CAS PubMed
- S. Broussy, R. W. Cheloha and D. B. Berkowitz, Org. Lett., 2009, 11, 305–308 CrossRef CAS PubMed
- S. P. Liu, R. X. Liu, A. A. M. M. El-Rotail, Z. Y. Ding, Z. H. Gu, L. Zhang and G. Y. Shi, J. Biotechnol., 2014, 30, 91–97 CrossRef PubMed
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c6ra17683f |
| This journal is © The Royal Society of Chemistry 2016 |