
Interaction between dendrimers and regulatory proteins. Comparison of effects of carbosilane and carbosilane–viologen–phosphorus dendrimers
 a
K.
Milowska,
*a
a
K.
Milowska,
*a
Abstract
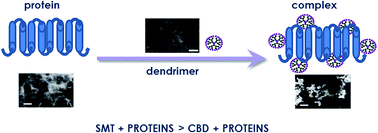
For nanoparticles to be used successfully in biomedical application, their interactions with biological fluids need to be investigated, in which they will react with proteins and other macromolecules. In this case, dendrimers might change their biological properties. Thus, the interaction of 2 generations of carbosilane–viologen–phosphorus and 2 types of carbosilane dendrimers with the 3 proteins – alkaline phosphatase from E. coli, human aspartate transaminase and L-lactate dehydrogenase from rabbit muscles, will be discussed. The techniques used included circular dichroism, zeta potential measurement, fluorescence quenching and transmission electron microscopy. The results show that positively charged hybrid and carbosilane dendrimers can interact with negatively charged regulatory proteins, and are able to form complexes. However, the spatial structure and the flexibility of dendrimer are as important as the protein configuration in complex formation.
 Please wait while we load your content...
Please wait while we load your content...

