
The in vivo estrogenic modulatory effect of bisphenol A (BPA) on Oreochromis mossambicus and prevention of early maturation of ovary by conjugates of intracellular laccase and silica nanoparticles†
Abstract
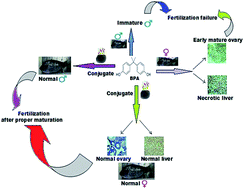
Bisphenol-A (BPA) is a well-known endocrine disrupting chemical (EDC). Employing Oreochromis mossambicus (Tilapia) as a model organism, this study aimed to analyse the estrogenic modulatory effect of BPA and to prevent the early maturation of ovary by treating with conjugates of intracellular laccase and silica nanoparticles. Intracellular laccase was extracted from Trametes versicolor and purification of laccase was performed using fast protein liquid chromatography after running through ion exchange (DEAE-cellulose) followed by a gel filtration (P-6) column. Sol gel method was used to synthesize silica nanoparticles which were characterized by transmission electron microscopy, atomic force microscopy, dynamic light scattering, Fourier-transform infrared spectroscopy and X-ray diffraction. Silica nanoparticles were amino-functionalized and conjugated with intracellular laccase. Isothermal titration calorimetry showed that conjugation of intracellular laccase with silica nanoparticles was stable and the reaction was exothermic and spontaneous. Maximum effective concentration of BPA was found to be 100 ppm after short term exposure of Oreochromis mossambicus to BPA, at this concentration it leads to deleterious effects on liver (disruption of tissues), ovary (early maturation) and testis (antagonistic effect) which was more prominent in histopathological studies. Vitellogenin protein level and gene level expression analysis using FPLC and quantitative real time PCR showed that conjugates of laccase and silica nanoparticle were more effective in reducing the BPA concentration when compared to free laccase enzyme. Thus from the result it clear that conjugates of intracellular laccase and silica nanoparticle represents a promising tool to reduce the estrogenic effect of BPA by preventing the early maturation of ovary in O. mossambicus.
 Please wait while we load your content...
Please wait while we load your content...

