
Biomimetic fabrication of biotinylated peptide nanostructures upon diatom scaffold; a plausible model for sustainable energy†

Abstract
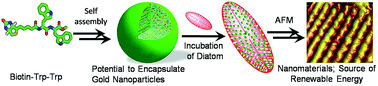
This study demonstrates the interactions of a biotinylated peptide with single cell eukaryotic microalgae known as diatoms, present in most natural water resources. The specifically designed biotinylated peptide consists of two key components, tryptophan along with the biotin, which helps us to unravel the interactions of this peptide with diatoms via spectroscopic tools owing to the high affinity biotin–avidin interactions and unique photophysical and photochemical properties of the Trp residue. The spectroscopic observations are further confirmed by atomic force microscopy (AFM), where the interactions of the biotinylated peptide afford a unique nanoarray over the diatom frustules. This peptide soft structure has potential for the encapsulation of gold nanoparticles which can be disrupted upon exposure of sunlight/radiation; hence this peptide can act as an adequate metal nanoparticle carrier for the diatom. Owing to thermoplasmonic heat of nanoparticles, the peptide can stimulate diatom frustules to open up, allowing oil to be extracted. Thus we hypothesize a most simple and parsimonious model for alternative/renewable energy resources and light-harvesting assemblies.
 Please wait while we load your content...
Please wait while we load your content...

