
Lipids influence the proton pump activity of photosynthetic protein embedded in nanodiscs†
Abstract
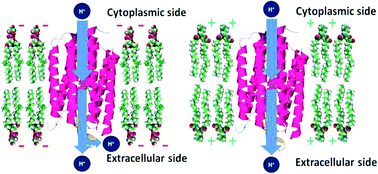
We report the lipid-composition dependent photocycle kinetics and proton pump activity of bacteriorhodopsin (bR) embedded in nanodiscs composed of different lipids. Using time-resolved spectroscopy and electrochemical methods, we were able to comprehensively understand the kinetics of the photocycle and the corresponding proton pumping activity as the composition of the charged lipids were systematically adjusted. We found that positively-charged lipids assist in repulsing protons from bR, thus increasing the concentration of the non-bounded protons in the bulk. In contrast, the negatively-charged lipids assist in entrapping the protons in the proximity of bR during the photocycle, preserving the electromotive force across the lipid bilayers which is essential for the vitality of the lateral proton transport and the bioenergetics.
 Please wait while we load your content...
Please wait while we load your content...

