
The effect of incubation conditions on the hemolytic properties of unmodified graphene oxide with various concentrations†
Yang Wang,
Baomei Zhang and
Guangxi Zhai*
Department of Pharmaceutics, College of Pharmaceutical Sciences, Shandong University, Jinan 250012, China. E-mail: professorgxzhai@126.com
First published on 5th July 2016
Abstract
The hemolysis assay is a common method used to evaluate the in vitro biocompatibility of biomaterials, but presently there is inadequate understanding of the interactions between RBCs and biomaterials, especially from the view of the incubation conditions and sample concentration. In this report, we used unmodified graphene oxide (GO) as a biomaterial model and measured the hemolytic properties of GO at a series of concentrations under multiple conditions, including medium type, red blood cell (RBC) species, RBC concentration, storage duration of the RBCs, animal health status, centrifugal speed, exposure time and incubation temperature. Additionally, a calculation method for the hemolytic index was proposed so as to objectively evaluate the hemolysis properties, distinct from frequently used methods. Compared to the hemolytic results under these conditions, we found that the hemolytic properties of GO had huge differences, and false-negative or false-positive errors might be produced by inappropriate incubation conditions. In our opinion, the feasible conditions for unmodified GO are: isotonic glucose solution as the medium type, fresh and healthy rabbit RBCs as the test object, 1% hematocrit as the RBC concentration, 3000 rpm for 5 min as the centrifugation conditions, 6 h as the exposure time and physiological temperature (37 °C) as the incubation temperature.
1. Introduction
Considering some of the advantages of graphene oxide (GO), such as a large specific surface area, strong physical adsorption capacity, its low cost of mass production and substantial functional oxygen-containing groups including hydroxyl, carboxyl, carbonyl and epoxy groups, GO has been widely applied to drug delivery,1 tumor imaging,2 photothermal therapy,3 bone tissue engineering4 and DNA manipulation.5 Since approximately 99% of the cells in the blood are red blood cells (RBCs),6 the hemocompatibility assay is an effective method in vitro to preliminarily estimate the biocompatibility of biomaterials using readily available RBCs from animal blood.7 In fact, hemolytic assays have been part of a currently momentous safety assessment of GO biomedical applications. The biocompatibility of GO has attracted considerable attention and has been summarized by several reviews,8–11 but these are still insufficient to understand the interactions between GO and RBCs, especially from the view of the incubation conditions and GO concentration. Likewise, the majority of contacted blood cells are RBCs after intravenous injection of GO suspensions. The destruction of RBC membranes and the leakage of intracellular hemoglobin (Hb) into the medium results from the contact and insertion of GO nanosheets by an alternation or distribution adjustment of the chemical components on the surface of the RBC membrane. Moreover, mature RBCs have no nucleus nor organelles such as mitochondria and ribosomes, and more than 95% intracellular proteins are hemoglobin, so the destruction degree can be evaluated by the hemolytic index (HI), which is a concentration ratio of the free Hb concentration divided by the total Hb concentration then multiplied by 100 in accordance with the American standard practice (ASTM F-756-00).12 HI values above the negative control fall into three classifications: (1) 0–2%, non-hemolytic; (2) 2–5%, slightly hemolytic; and (3) >5%, hemolytic.Some investigations have studied the hemolytic properties of unmodified GO, but the results have differed largely, even controversially. For instance, Liao et al.13 Singh et al.14 Cheng et al.15 Feng et al.16 He et al.17 and Wang et al.18 showed that hemolytic activity of GO was severe, while it was relatively low in the work of Wu et al.19 Zhang et al.20 Pinto et al.21 and Zhou et al.22 Why did the same biomaterials have distinct results? A general conclusion drawn from these investigations is that the experiment conditions varied from case to case in the hemolysis assay. Henkelman et al.23 found this issue and believed that there was no consensus on the procedures of the hemolysis assay in most research because of insufficiently comparative studies of the incubation conditions. These conditions include the species, quality and the storage duration of the RBCs, the medium type and osmolality, the incubation pattern (static, semi-static, or dynamic on a rotator with or without air), exposure time, incubation temperature, concentration ratio of RBCs and biomaterials, centrifugal speed and time. In the work of Henkelman,23 rabbit RBCs were deemed good choices due to the susceptibility to hemolysis of biomaterials after comparison with human RBCs. The washed RBCs should be used within 7 days with marginally increasing hemolysis after 7 days. The hemolysis percentages of most materials were similar with and without air during the test. However, the effects of the incubation conditions and sample concentration on the hemolytic properties have not yet been investigated in a prospective study. In this article, we chose GO as a biomaterial model and discussed the effects of incubation conditions on the hemolytic properties of GO so as to offer some suggestions on how to establish a normative procedure for the objective evaluation of the hemolysis of biomaterials. Additionally, a newly developed method was compared to two traditional methods.13,14,24
2. Experimental
2.1 Synthesis and characterization of GO
Unmodified GO was prepared by a modified Hummers method.25 Briefly, 1 g graphite powder (99%, D50 < 400 nm, Lot: A1402053, Aladdin Industrial Co., Ltd.) was soaked in 23 mL concentrated H2SO4 for 12 h to form peroxided graphite for a more uniform oxidization, then 3 g KMnO4 as an oxidizing agent was added in an ice bath. After the mixture was stirred at 40 °C for 30 min, 46 mL distilled water was slowly added and then mixture was kept at 100 °C for 30 min to produce GO. Subsequently, the mixture of 140 mL distilled water was blended with 10 mL 30% H2O2 solution and added into the GO suspensions: a mixture with a muddy color was obtained and stored at 4–8 °C overnight to form precipitates. The precipitates were repeatedly centrifuged (10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 rpm for 10 min), washed three times with 5% HCl and water in turn, suspended in water, ultra-sonicated for about 2 h, and dialyzed against water for 48 h.
000 rpm for 10 min), washed three times with 5% HCl and water in turn, suspended in water, ultra-sonicated for about 2 h, and dialyzed against water for 48 h.
UV spectra were determined by a UV–visible spectrophotometer (Beijing Purkinje General Instrument Co., Ltd., Beijing, China). The topography of the unmodified GO was investigated using an atomic force microscope (AFM, MultiMode IIId, Veeco Instruments Inc., Bruker AXC Inc., Madison, Wisconsin, USA). The images were flattened and plain fitted using NanoScope analysis software (version 1.7, Bruker AXC Inc., Madison, Wisconsin, USA). Raman spectra were obtained using a Horiba Jobin Yvon LabRAM HR-800 Raman microscope (λ = 532 nm). Fourier transform infrared (FTIR) transmittance spectra were obtained using a Nicolet 6700 Fourier transform infrared spectrometer (Thermo Nicolet Corp., Madison, WI, USA) and collected in the region 400–4000 cm−1 by OMNIC software (Thermo Nicolet Corporation, Madison, WI, USA).
2.2 Calculation of the hemolytic index
According to ASTM F-756-00, the hemolytic index is calculated using the free and total Hb concentrations. The fixed molar ratio between Hb and heme was 1:4 because one Hb molecule contains four heme molecules. So CHb was substituted by Cheme and the hemolytic index (HI) could be calculated by eqn (1).
 | (1) |
The molar extinction coefficient of heme (εheme) is known as 13.8 mM−1 cm−1,26 and the unit of heme concentration was irrelevant to the calculated results of hemolytic index regardless of molar concentration or mass concentration, so molar concentration was used in our work.
Csampleheme, Cnegheme, Ctotalheme were the heme concentrations of samples, the negative and total, respectively. Cheme was solved by one set of binary linear algebraic expressions, eqn (2) and (3). A540, A576 and A600 were the absorbance values of samples at 540, 576 and 600 nm, respectively. A540 − A600 and A576 − A600 were two-point absorbance difference values. a1, a2, b1, b2, c1 and c2 were various coefficients depending on the properties of the biomaterials and RBC species. CGO was GO concentration. Only Cheme was our interest in the hemolysis assay. The new method is called the 540-576-600 method henceforth.
| A540 − A600 = a1Cheme + b1CGO + c1 | (2) |
| A576 − A600 = a2Cheme + b2CGO + c2 | (3) |
In order to solve (2) and (3), the coefficients including a1, a2, b1, b2, c1 and c2 need to be determined first of all. Rabbits (2.50–3.50 kg), rats (200–250 g) and guinea pigs (200–250 g) were purchased from the Laboratory Animal Center of Medical School, Shandong University. The Shandong University Institutional Animal Care and Use Committee approved animal care and procedures. Animal use was in accordance with National Institutes of Health guidelines. First, the purified rabbit Hb was separated from the RBCs by previously described procedures.27 Briefly, blood drawn from healthy rabbits by intracardiac puncture was collected in polyethylene tubes containing 10 IU mL−1 heparin28 and centrifuged at 3000 rpm for 5 min to remove the plasma and buffy coat. The remaining cells were centrifuged and washed repeatedly with isosmotic 5% glucose solution until the supernatants were colorless. Then 1 mL of packed RBCs was suspended in 2 mL of deionized water at room temperature for 20 min, and then centrifuged at high speed (12000 rpm for 15 min) in order to separate it from the cellular debris. The blood from guinea pigs was drawn by intracardiac puncture and from rats by the jugular vein. The extracted Hb was obtained by the above approach. Secondly, the solutions containing a known heme concentration without GO were prepared, and another series of heme solutions with GO nanosheets were prepared. Each sample was repeated thrice. Third, these mixtures with the known CGO and Cheme were scanned in the region from 500 to 655 nm, and the absorbance data of three points at 540, 576 and 600 nm were collected. Finally, the linearity results were subjected to a least squares linear regression analysis (ESI S1†). The binary linear algebraic equations were fitted and solved by operating Matlab R2011a software (version 7.12.0.635, MathWorks Inc, Natick, Massachusetts, USA).
Ctotalheme was calculated using eqn (4). The mean corpuscular hemoglobin concentration (MCHC) was 324 ± 11 g L−1 for rabbits,29 370 ± 5 g L−1 for rats,30 and 336 ± 2 g L−1 for guinea pigs.31 The molecular weight of Hb was 69737.13 g mol−1 for rabbits (PDB ID: 2RAO, seen the website: http://www.rcsb.org/pdb/home/home.do), 64156.52 g mol−1 for rats (PDB ID: 3HF4), and 65317.24 g mol−1 for guinea pigs (PDB ID: 3AOG). HCT is the hematocrit in RBCs suspension; 4 refers to the fixed coefficient of molar concentration between Hb and heme; and Mr is the molecular weight of Hb.
 | (4) |
2.3 Medium type
Two types of isotonic medium were used, namely, 5% glucose solution and phosphate buffer solution (PBS, pH 7.4). The above packed RBCs from rabbits were washed repeatedly with isotonic glucose solution or PBS. 2 mL GO colloidal suspensions at a series of concentrations were dispersed in the respective medium and mixed with 2 mL RBC suspensions. Finally, the final suspensions contained 5, 10, 15, 20, 25, 30, 40, 50, 75 and 100 μg mL−1 GO with 1% RBCs, respectively. The negative and the positive control samples were prepared in a similar manner. The former had added 2 mL isotonic solution (5% glucose or PBS) without GO, and the latter had added 2 mL water. All mixtures were incubated in a water bath at 37 °C for 3 h, and centrifuged at 3000 rpm for 5 min. The supernatants were collected carefully to remove any large aggregates and were scanned from 500 to 655 nm by UV spectrophotometry.2.4 RBCs species
The healthy RBCs were from rabbits, guinea pigs and rats. According to the procedure for the 2.3 medium type, the RBC suspensions were blended with GO suspensions in equal volumes. The final suspensions were 1% RBC with 5% glucose for GO samples and the negative control. The positive control contained 1% RBC with 2.5% glucose.2.5 RBC concentration
The final suspensions contained 0.5, 0.75, 1, 1.5 and 2% RBC from healthy rabbits with 5% glucose, respectively.2.6 Storage duration of the RBCs
The 2% RBC suspensions from healthy rabbits with 5% glucose as the stock suspensions were divided into three groups (A, B and C) and stored at 4–8 °C. Group A was tested immediately, and groups B and C were tested on the second and fourth days, respectively.2.7 Animal health status
Healthy and anemic rabbit RBCs groups were used. The anemic group was made anemic by repeated bleeding for 7 days, which is a usual approach to acute anemia in an animal model.322.8 Centrifugal speed
All mixtures were centrifuged at 0, 1000, 2000, 3000, 4000, 6000 and 8000 rpm for 5 min, respectively. The calculated hemolysis percentages were based on the lowest GO concentration (10 μg mL−1) without centrifugation as a reference.2.9 Exposure time
Three GO concentrations were used including low (10 μg mL−1), middle (40 μg mL−1) and high (80 μg mL−1). The exposure time lasted 24 h at 37 °C and centrifuged at 3000 rpm for 5 min. A negative control without GO was conducted as a reference.2.10 Incubation temperature
The incubation temperatures were set at 27, 32, 37, 42 and 47 °C for 6 h. The data from before and after centrifugation were compared.3. Results and discussion
3.1 Synthesis and characterization of GO
Unmodified GO was manufactured and dispersed in water at a series of concentrations (Fig. S1†).The UV spectra of GO showed that the maximum absorption peak at about 230 nm was attributed to the π–π* transitions of the aromatic bonds (C![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) C) in sp2 hybrid regions, and a shoulder peak at 320 nm or so originated from the n–π* transitions of the carbonyl group (CO) in the sp3 hybrid regions (Fig. S2a†).14,33 The absorbance of GO steadily increased from 700 nm to 230 nm.
C) in sp2 hybrid regions, and a shoulder peak at 320 nm or so originated from the n–π* transitions of the carbonyl group (CO) in the sp3 hybrid regions (Fig. S2a†).14,33 The absorbance of GO steadily increased from 700 nm to 230 nm.
The Raman spectra of the GO nanosheets showed five characteristic vibrations of D, G, 2D D + G and 2G bands around 1362, 1585, 2724, 2937 and 3186 cm−1, respectively (Fig. S2b†).33 The three latter peaks appeared relatively weak, the two former peaks were higher at 1362 and 1585 cm−1. The increased intensity of the D band at 1362 cm−1 indicated that the planar structure of graphite had been destroyed and that GO subsequently formed.34 The intensity ratio of the D and G bands (ID/IG) was 0.80 for the GO nanosheets. This result is consistent with 0.79 in the previous study.35
The multiple peaks of the IR spectra of GO were observed in the region 800–1800 cm−1 (Fig. S2c†), which could be assigned to the oxygen-containing functional groups, including C–O (νC–O at 1090 cm−1), C–O–C (νC–O–C at 1275 cm−1), C–OH (νC–OH at 1365 cm−1), and CO in carboxylic acid and carbonyl groups (νCO at 1720 cm−1). The peak intensities at 2800–2950 cm−1 (–CH–, stretching vibrations) increased significantly for GO. A large and broadened peak at 3000–3700 cm−1 showed the stretching vibrations of hydroxyl groups due to the presence of absorbed water and hydroxyl moieties from the GO.15
The surface structure and morphology of the GO were characterized by AFM (Fig. S2d†). The irregular GO nanosheets had a large aspect ratio, since the data obtained by NanoScope analysis software showed that the average diameter was 156 ± 213 nm (SD) with a smallest diameter of 35 nm and a largest of 1005 nm, and that mean thickness was approximately 1 nm. Additionally, the mean surface area obtained by the software was 54991 nm2. The minimum and maximum were 977 and 793945 nm2, respectively.
3.2 Calculation of the hemolytic index
The sets of binary linear algebraic equations from the three species were fitted (ESI S1†). The calculation method for the hemolysis percentage was compared with two frequently used methods (ESI S2†).Obviously, the new method proposed in this work provided more accurate data compared with the two traditional methods. The first reason that false-positive errors are produced using traditional methods is the incomplete release of Hb for the positive control. RBCs cannot fully rupture in suspension after treatment with the current incubation conditions, so the measured values of total heme concentration could not reach the true values that originated from data published in the literature. The calculated true Ctotalheme was far more than that measured (in 1% RBC suspension, 185.84 ± 6.31 vs. 120.40 ± 1.39 μmol L−1 for rabbits, 230.69 ± 3.12 vs. 46.16 ± 3.56 μmol L−1 for rats, and 205.76 ± 1.22 vs. 93.83 ± 8.63 μmol L−1 for guinea pigs, in the section of 2.4 RBCs species). The second was strong background absorption for the colored GO from 500 to 655 nm. The background absorption was not deducted from the traditional methods and led to false-positive errors. In our new method, we considered the interference of GO, Csampleheme and Cnegativeheme were calculated by one set of binary linear algebraic equations. For good linear relationships between the absorption and concentration of heme or GO, two maxima (at 540 and 576 nm) and one minimum (at 600 nm) were selected. The low mean deviation of the fitted and actual heme concentrations showed that the new method was suitable for the calculation of Cheme (2.58% for rabbits, 9.21% for guinea pigs, and 18.83% for rats, see ESI S1†).
3.3 Medium type
The difference in the hemolysis properties of GO nanosheets was distinct in two types of medium (Fig. 1). In PBS, the hemolysis percentage of rabbit RBCs gradually reduced from 0 to −17% with increasing GO concentration, and Cfreeheme fell from 27 μmol L−1 to nearly zero when the GO concentration was over 30 μg mL−1. | ||
| Fig. 1 The effect of the medium on hemolysis caused by GO. (a) Photographs of hemolysis caused by GO in isotonic PBS and glucose solution. (b) Hemolysis curves from two incubation media. D.I. water (+) was used as the positive control. Concentration is abbreviated as “con.” in the figures, henceforth. | ||
For the negative control in two groups, a small amount of senescent RBCs were attributed to the free Hb released from the RBCs. Compared with the mature RBCs, the aged RBCs greatly changed in deformability, membrane elasticity, and the ratio of surface area to volume.36
In PBS, the successive decline of the hemolysis percentage was due to massive aggregation of GO nanosheets: when GO made contact with cations (e.g. Na+ and K+), the negative charges on the GO surface were neutralized, then the intersheet repulsion force between GO nanosheets sharply declined, and eventually GO precipitated at the bottom of the container after centrifugation. This is known as salting out37 and might be the primary cause of negative hemolysis. The stability of aqueous unmodified GO suspensions was determined by several factors including the surface zeta potential, hydration degree of the oxygen-containing groups and sheet size. The reduction of the effective surface area in contact with RBCs after precipitation of GO nanosheets led to a corresponding reduction of the hemolysis.
However, the hemolysis curve of GO presented a trend of sustained increase in glucose solution after a slight decrease during the initial stage (Fig. S3†). Interestingly, the hemolysis caused by GO with less than 15 μg mL−1 was below that of the negative control. The well-dispersed GO nanosheets were extremely thin in water suspension and were penetrating as a scalpel, only approximately 1 nm of thickness was observed in our experiment. Due to inorganic ions (e.g. Na+, 0.178 nm38) with smaller radii and Hb with a larger radius (3.17 nm39), the first released substances were intracellular ions (9.2 ± 2.7 and 112 ± 8.6 mmol L−1 for Na+ and K+ in rabbit RBCs, respectively40) after slight damage of the RBC membranes, and then Hb escaped from the RBCs. The negative hemolysis percentages demonstrated that GO with a low concentration has a weak disruptive effect on RBCs, which led to the liberation of a small amount of intracellular ions. After aggregation and precipitation of GO by these ions, the sharp edge of GO disappeared and the hemolysis properties subsequently dropped by a large margin. GO nanosheets had a strong absorption force and the adsorption capacity of GO was positively correlated with its concentration. The precipitated GO partly absorbed free Hb in suspension and then the Hb concentration in the supernatant declined to almost zero, which might be the reason for the negative hemolysis percentage.
GO could drastically extract a large quantity of phospholipid from the membranes of E. coli after insertion and cutting through the cell membranes.41 However, the adhesion energies obviously declined from 0.45 ± 0.02 to 0.31 ± 0.01 J m−2 for the monolayer and multilayer graphene membrane.42 The adsorption effect of GO nanosheets on the Hb molecules most likely kept a balance such that the adsorption capacity increased with GO concentration, while the effective surface area correspondingly declined.
The hemolysis curve showed that the hemolysis assay in isotonic glucose solution was successful, and reached our expected results that hemolysis increased with GO concentration. From the above discussion, it was crucial to choose the incubation medium and to set the GO concentration parameters for the hemolysis assay. The isotonic glucose solution as an incubation medium was reasonable, and the range of the GO concentration from 0 to 100 μg mL−1 was feasible for objective evaluation of the hemolysis properties.
3.4 RBC species
The RBC effects from three species (rabbits, rats and guinea pigs) on hemolysis were compared (Fig. 2 and S5†). High hemolysis occurred in rabbit RBC suspensions, especially at higher GO concentrations such as 50, 75 and 100 μg mL−1, which is consistent with previous reports.23 The rabbit RBCs underwent massive hemolysis under the influence of GO nanosheets and were more susceptible than human RBCs.23RBCs from rats showed almost no hemolysis at any GO concentration except 100 μg mL−1 (hemolysis with 3.08%). However, RBCs from guinea pigs exhibited negative hemolysis at every GO concentration. Similar results for RBCs from rats were reported by Zhang et al.20 and the hemolysis was below 5% at less than 80 μg mL−1 of GO concentration after incubation for 3 hours. The negative hemolysis indicated there was no damage or slight damage of the RBCs from rats and guinea pigs after incubation with GO nanosheets.
For RBCs from guinea pigs, the hemolysis caused by GO at a series of concentrations was lower than those of the negative control, and the pale yellow of the supernatants showed that almost all GO nanosheets had precipitated after centrifugation (Fig. 2a), which resulted from minor damage of RBCs membranes and abundant release of the intracellular inorganic ions with smaller radii. At the same time, most of the Hb molecules still remained inside the RBCs due to the slight destruction of the RBC membranes.
 | ||
| Fig. 2 The effect of RBC species on the hemolysis properties of GO after centrifugation. (a) Photographs of the hemolysis of RBCs from rabbits, rats and guinea pigs, respectively. (b) The hemolysis curves from three RBC species. The hemolysis increased steadily with GO concentration because of the strong interaction between GO and RBCs from rabbits after heat treatment for 3 h. For RBCs from rats, GO had nearly no hemolysis, while GO produced negative hemolysis for RBCs from guinea pigs owing to probable absorption of Hb and protection against RBCs. | ||
The hemolysis of rat RBCs was completely different from the other two types of RBCs. The brown color of the GO was deepened with its concentration, and the amount of precipitation was apparently unchanged, which showed that the GO nanosheets still existed in the supernatants and that the RBC membranes had not been destroyed by GO.
The different sensitivity of the RBCs from different animal species to the hemolysis properties of snake venom was reported over fifty year ago.43 Studies showed that the contents and distributions of the chemical compositions that included cholesterol, glycolipid, ganglioside and phospholipid on RBCs membranes were closely associated with the animal species.43 Furthermore, the phospholipid distributions of the RBCs were distinct for various species and phospholipids.44 These differences result in distinct characteristics of RBCs, such as surface charge, size, volume, and their lifetime in vivo. For example, the weight percentages of the total lipid and cholesterol on RBC membranes were different (28 ± 1.0% and 21 ± 2% for rabbits, 22 ± 1.0% and 28 ± 2% for rats45). The lower cholesterol percentage for rabbit RBCs was attributed to higher hemolysis since the introduction of cholesterol in the lipid bilayers normally reduces the permeability of RBC membranes.46,47 However, too high a cholesterol percentage also induced in vivo serious hemolysis because the membrane became highly fragile and had lower flexibility after the investigation on cholesterol-fed guinea pigs.48
The results illustrated that GO might lead to different hemolytic properties even with the same GO concentration due to the use of RBCs from different species. In order to lower the hemolysis risk of human RBCs, rabbits RBCs are a good model to evaluate the hemolysis properties of biomaterials including GO.
3.5 RBC concentration
RBC concentration greatly influenced the results of the hemolysis percentages (Fig. 3 and S6†). Although the free heme concentration for RBCs with lower hematocrit (e.g. 0.5% RBCs) was low, the total heme concentration (92.92 μmol L−1) was also low, so the calculated hemolysis percentages were relatively higher with the increase of GO concentration. The results illustrated that RBCs from rabbits with low concentration underwent wide and substantial destruction when the GO concentration was high.For a better understanding, the surface areas of the RBCs and GO molecules were calculated (ESI S3†). On the one hand, the surface area and volume of one rabbit RBC were 96.7 ± 10.2 μm2 and 60.9 ± 12 μm3, respectively.36 The calculated total RBC count and the total calculated surface area were about 1.64 × 108 cells mL−1 and 15.9 × 10−3 m2 mL−1 for 1% RBC, respectively. On the other hand, the specific surface area of the GO nanosheets, experimentally measured in diluted GO solutions, was 736.6 m2 g−1, and the estimated aggregation level was 15% for 50 μg mL−1 GO.49 The calculated surface area of 10 μg mL−1 GO was about 7.37 m2 mL−1.
The surface area of the total GO nanosheets over 10.79 μg mL−1 was larger than that of the 0.5% RBC suspension by quantitative calculation. RBCs in 0.5% suspensions had considerable interaction with GO molecules at a higher concentration (e.g. 40 μg mL−1). The interactions between GO and RBCs included absorption, cutting and insertion. Because the dielectric constant in the glucose solution was reduced50 and glucose molecules attenuated the interaction between the GO nanosheets, the critical aggregation concentration of GO might be above 35 μg mL−1. The hemolysis percentage reached a maximum (about 23%) at 40 and 50 μg mL−1 GO concentrations for 0.5% RBC (Fig. 3b). Moreover, the lower hematocrit in the RBC suspensions was unsuitable for hemolysis evaluation since larger operation errors originated from the incubation conditions with marginal variations.
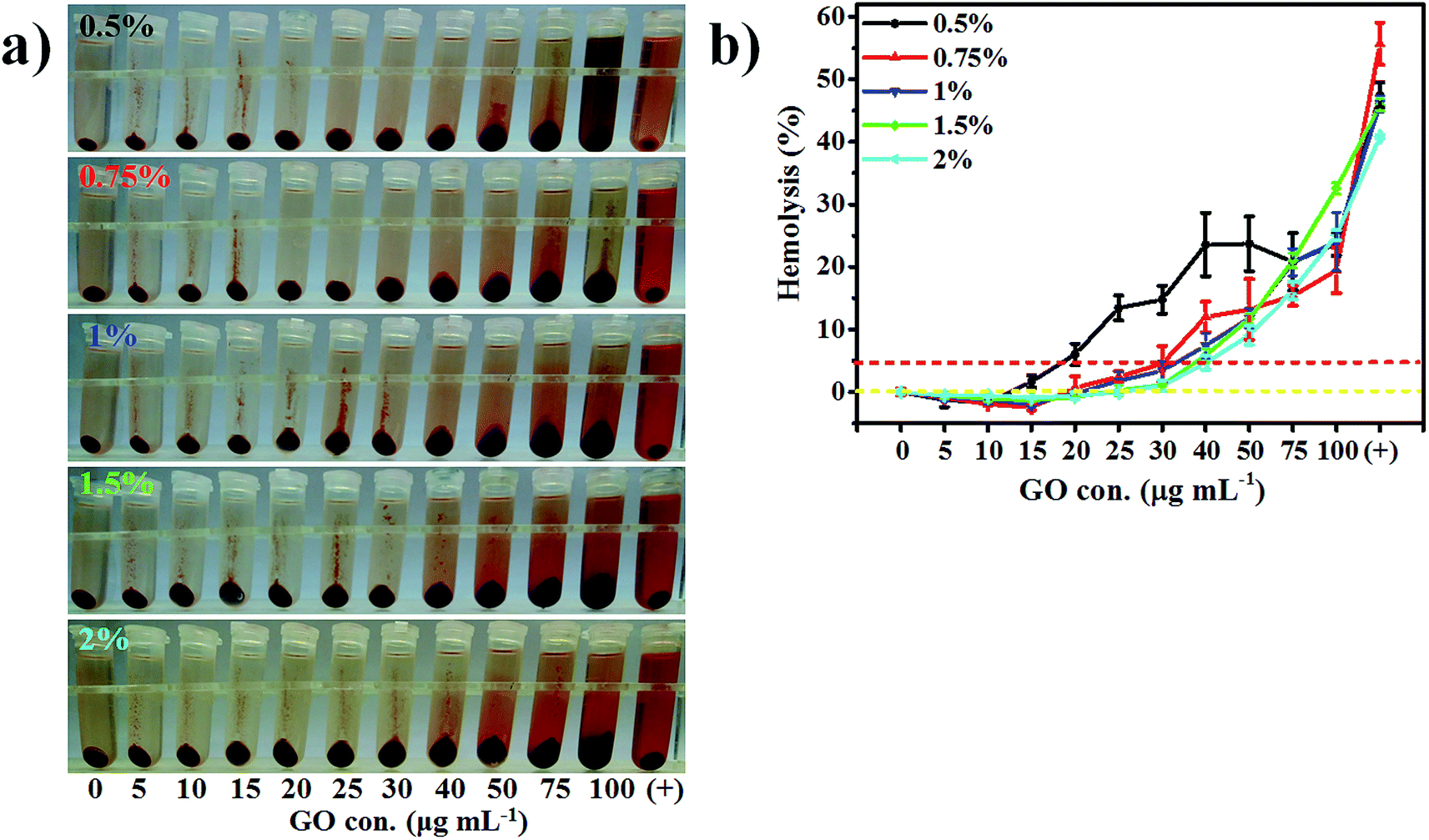 | ||
| Fig. 3 The effect of RBC concentration on hemolysis assays of GO. (a) Photographs of hemolysis of 0.5, 0.75, 1, 1.5 and 2% RBC, respectively. (b) The hemolysis curves from five RBC concentrations. The results exhibited relatively high hemolysis caused by GO nanosheets in the range of 15–75 μg mL−1 of GO concentration in 0.5% RBC, slightly high hemolysis in the range of 20–50 μg mL−1 of GO concentration in 0.75% RBC and similar hemolysis degrees for the positive control for five RBC concentrations except for 0.75% and 2% RBC. | ||
For higher RBC concentrations such as 1.5 and 2% the extent of interaction between GO and RBCs was low and partial, which caused the heme concentration values in the supernatants to be similar at the same GO concentration (Fig. S6b†), and the hemolysis curves were almost overlapping (Fig. 3b). These results indicate that the higher RBC concentration (more than 1%) was wasteful because excessive RBCs could not produce high hemolysis. Therefore, 1% RBC concentration was advisable and met the requirements of the experiments in which hemolysis gradually increased with the increase of GO concentration and the interaction between GO and RBCs increased continuously.
Additionally, in accordance with ASTM F-756-00, samples with a determinable surface area of 21 cm2 were used and 1 mL blood with a total Hb content of 10 ± 1 mg mL−1 was added. The corresponding GO concentration and RBC concentration were 0.356 μg mL−1 and 3.1% to evaluate hemolysis, respectively (ESI S3†). A low GO concentration (0.356 μg mL−1) caused low hemolysis. Furthermore, a 3.1% RBC concentration was too high in the assay and produced a relatively low hemolysis percentage. A high RBC concentration was used in the previous reports. 10% of the final RBC suspension was applied by Cai et al.51 and Zhang et al.,20 2.5% was used by Wang et al.,52 and 1.33% by Wang et al.18
3.6 Storage duration of the RBCs
The storage duration effect of the RBCs on hemolysis was investigated in a 5% glucose solution on the first, second and fourth days (Fig. 4 and S7†). The hemolysis caused by GO gradually crept up with the increase of GO concentration and storage duration. However, hemolysis of the positive control declined close to the value of the negative control with time, which showed the opposite trend. Furthermore, the hemolysis had larger errors for the negative control due to two probable accounts.Every RBC has a finite life span (approximately 50 days for rabbit RBCs53). Everyday about 2% of senescent RBCs loose their activity, easily destroyed by external conditions, and produce hemolysis. Secondly, when exposed to a glucose medium of high levels for a long time, RBC membranes acquire damage.54 The free Hb concentrations were 19.70 ± 6.02 and 49.02 ± 1.94 mg dL−1 for 24 h and 48 h,55 respectively, after incubation of human RBCs at 45 mmol L−1 of the final glucose concentration. The membrane changes related to glucose-based reactions originate from several mechanisms including glucose autoxidation, protein glycation, oxidative damage of some amino acid residues56 and formation of advanced glycation end-products.57 The oxidative extent of the RBC membrane is associated with glucose concentration and storage duration,55 although the normal biconcave shape of RBCs was not prominently altered after incubation with glucose media for three days in our assay. The RBC membrane became more fragile and hemolysis continually increased.55 Our findings were consistent with Buhler’s report.58
For the positive control, the hemolysis gradually declined with storage time for unknown reasons, perhaps explained from the view of molecular movement. The dynamic mass exchange on both sides of the lipid bilayer changes via the permeable membrane during storage of RBCs. Intracellular ions (e.g. K+ and Cl−) cross the membrane and leak into the extracellular medium, and glucose molecules enter into the RBCs via facilitated diffusion involving a specific glucose transport protein, GLUT-1.58 The diffusion rate of glucose is directly proportional to the glucose concentration gradient, temperature, and surface area of RBCs available for diffusion. The intracellular glucose underwent a non-enzymatic condensation, resulting in glycosylated Hb,59 which considerably influenced the physicochemical properties of Hb. Later RBC membrane deformability decreased and intracellular viscosity increased. These facts induced little change of hemolysis percentage on the fourth day (5.16 ± 1.73% vs. 63.82 ± 0.77%, Fig. 4) and caused false-negative results after RBCs were exposed to 2.5% glucose medium.
 | ||
| Fig. 4 The storage effect of RBCs on hemolysis assays of GO. (a) Photographs of hemolysis of RBCs stored on the 1st, 2nd and 4th days, respectively; (b) the hemolysis curves of the stored RBCs. The hemolysis properties of the GO nanosheets were relatively high for RBCs stored for three days. RBCs suffered relatively minor damage from GO nanosheets on the first day probably because the erythrocytes were most vibrant and the membranes were complete. | ||
For rising hemolysis with a change of GO concentration, decreased membrane deformability and massive protein damage of the RBC membrane were the main internal factors, while the sharpness of the planar GO structure and existence of substantial GO nanosheets were the external factors, which led to false-positive errors.
Fresh RBC suspensions in isotonic glucose solution should be used immediately so as to effectively avoid hemolysis with false-positive errors for GO or false-negative errors for the positive control.
3.7 Animal health status
The aim of this assay was to evaluate the relationship between hemolysis and animal health status. Two statuses of rabbit RBCs were used. The prevalently high hemolysis in the anemic group could be illustrated by the proportion and features of the reticulocytes (also called young RBCs) and mature RBCs (Fig. 5 and S8†). | ||
| Fig. 5 The health statue effect of RBCs on the hemolysis properties of GO. (a) Photographs of RBC hemolysis from healthy and anemic rabbits. (b) The two hemolysis curves from healthy and anemic rabbits. The increasing hemolysis from anemic rabbit blood was more intense than that from healthy blood with the increase of GO concentration. But the positive control had opposite results. These results illustrated that the different health statuses of rabbit RBCs were significant for the hemolytic properties of GO. | ||
The proportion of reticulocytes was relatively low in the whole blood (only 1–2%60 or 0.40 ± 0.15%61 in normal rabbit blood), which is closely related to duration of disease and the degree of anemia.60 The proportion increased to 15–20% after 9 days32 or 15–30% after 5 days60 for rabbit reticulocytes.
The distinct features of reticulocytes have many aspects. The low content of sialic acid on RBC membranes showed reduced surface charges,62 which meant more damage from external conditions, and led to high hemolysis. A larger cell volume (96.1 vs. 65.0 μm3,32 about 1.5-fold), and a lower concentration of Hb and cholesterol, and reduced density, led to increments of membrane flexibility and fluidity of the intracellular Hb.63 Moreover, a low level of cholesterol could enhance membrane permeability. These alterations made reticulocytes more susceptible to hemolysis than mature RBCs after attack by GO. For the anemic group that produced false-positive errors in hemolysis, the maximum hemolysis reached about 50 to 75 μg mL−1 GO. The subsequent decline mainly originated from precipitation and reduction of the effective surface area of most GO nanosheets.
For the positive control, since membrane flexibility and resistance to hypotonic hemolysis was enhanced, reticulocytes were influenced only by the osmotic pressure without GO and showed low hemolysis, which agrees with Chalfin et al.32 and Jaffe et al.64
Given the relationship between the health statuses of the RBCs and hemolysis, blood collection should be carried out without a notable reticulocyte count (less than 2%), and repeated bleeding of rabbits should be avoided.
3.8 Centrifugal speed
The purpose of setting the centrifugal time to 5 min is to simplify experimental system in the assay. In general, the lower the centrifugal speed, the weaker hemolysis caused by the GO (Fig. 6 and S9†). The fitted hemolysis curves were linear (Table 1) owing to three probable reasons. The first was the great repulsive interaction between GO and RBCs with the same negative charges. The carboxyl groups presented at the edge of the GO structure.11 A large number of sialic acid on the surface of the RBC membranes caused the negative zeta potential of the rabbit RBCs.65 The brown color of GO with 100 μg mL−1 was faint in the supernatants with centrifugal speed from 0 to 8000 rpm, which showed that GO had overcome repulsive forces and formed more precipitates. The second was the aggregation of GO nanosheets by cations that escaped from the ruptured RBCs. The centrifugal speed was correlated with the degree of membrane damage and hemolysis. The last was the thermal motion of GO nanosheets in the suspensions. RBCs suffered from random and severe attack by GO nanosheets at high concentration, which meant that hemolysis was positively related to GO concentration in the same centrifugal field. | ||
| Fig. 6 The centrifugation effect on hemolysis caused by GO. (a) Photographs of RBC hemolysis after 0, 1000, 2000, 3000, 4000, 6000 and 8000 rpm. (b) The seven hemolysis curves. Severe hemolysis was observed at high centrifugation speeds of above 4000 rpm. The centrifugation speed of 8000 rpm showed the maximum hemolysis percentage compared with the other speeds. | ||
| Centrifugation speed | Fitting equation | Regression coefficient |
|---|---|---|
| a The linear range was 10–100 μg mL−1. | ||
| 0 rpm | HI = 0.0771 × CGO − 0.3783 | 0.7946 |
| 1000 rpm | HI = 0.0962 × CGO − 0.4328 | 0.9857 |
| 2000 rpm | HI = 0.1271 × CGO − 1.1098 | 0.9932 |
| 3000 rpm | HI = 0.1225 × CGO − 0.2751 | 0.9825 |
| 4000 rpm | HI = 0.1297 × CGO + 0.5247 | 0.9614 |
| 6000 rpm | HI = 0.1739 × CGO + 0.4874 | 0.9508 |
| 8000 rpm | HI = 0.2109 × CGO + 2.4205 | 0.9141 |
For 2000, 3000 and 4000 rpm, the regression coefficients were superior to the others (more than 0.98), and the slope values were stable (0.12–0.13), which illustrated that the centrifugal speed from 2000 to 4000 rpm was almost unrelated to the hemolysis percentage and could objectively evaluate the hemolysis. The regression coefficient without centrifugation was worse than the others because of the existence of cellular debris in the supernatants that might interfere with the measurement. The slope for 8000 rpm reached a maximum (0.2109) and caused false-positive errors, which implied that the centrifugal speed strengthened RBC destruction and hemolysis.
How did the centrifugal speed influence the hemolysis of RBCs? The shear stress exerted on the RBC membranes was analyzed (ESI S4†). Briefly, assuming that the contact length of GO on the surface of an RBC membrane was 10 nm, then the calculated shear stress (from 0.009 to 462.182 Pa) partly exceeded the measured threshold (150 Pa66). In fact, the thermal motion of the GO nanosheets in the suspensions before centrifugation was the main factor and RBC damage was relatively slight in low centrifugal fields, while shear stress was predominant in high centrifugal fields. The hemolysis was 15.60 ± 0.55% at 8000 rpm for 40 μg mL−1 GO and surged to almost four times the original value as compared with 2000 rpm (4.04 ± 0.79%).
The centrifugal speed and time has been one of the most neglected factors and might cause false-positive errors due to high centrifugal speed and long times. Various conditions have been reported, including 1000 rpm for 10 min,67,68 1500 rpm for 10 min,22,69,70 1000 g for 10 min,71 3578 g for 1 min,23 4000 rpm for 10 min,20 4000 rpm for 15 min,72 4000 g for 5 min,73 10000 g for 3 min,51 10000 g for 10 min52 and 10016 g for 3 min.17,18 Considering false-positive errors, it is recommended to select 3000 rpm (equivalent to 603 × g) as the centrifugal speed in assays.
3.9 Exposure time
The exposure time was at least three hours in accordance with ASTM F-756-00, which implied that exposure time might have an important impact on hemolysis, although short exposure times have been reported, such as 0.5 h.22,70,72 Our assay found that hemolysis caused by GO was closely associated with GO concentration and exposure time (Fig. 7 and S10†). The higher the GO concentration and the longer the exposure time, the more severe the hemolysis. After 6 h, hemolysis was almost two times the original value for all GO concentrations, compared with 3 h. After about 12 h the hemolysis reached a maximum (13.29%, 25.64% and 42.38% for low, mid and high GO concentration, respectively). The number of GO nanosheets was calculated (ESI S5†). The ratio of maximum hemolysis was about 1:2:3. The ratio of GO concentrations was 1:4:8, disproportional to the ratio of maximum hemolysis owing to considerable aggregation of GO nanosheets with a concentration of more than 35 μg mL−1, although a high GO concentration contributed to an effect of amplified hemolysis. The regression coefficient was over 0.90 for all GO concentrations (Table 2), which showed that the damage extent to RBCs was a function of exposure time and GO concentration. For the negative control, hemolysis was independent of time as a result of the stability of RBCs in glucose solution. Generally, the hemolysis curves included at least two stages, namely, a rapid rise and a slow rise. These results agreed with the previous studies.18,20
 | ||
| Fig. 7 The relationship between exposure time and the hemolysis properties of GO. (a) Photographs of RBC hemolysis from 0 to 24 h at four GO concentrations including neg (0 μg mL−1), low (10 μg mL−1), mid (40 μg mL−1) and high (80 μg mL−1) concentrations. (b) The hemolysis curves of four GO concentrations. An obviously increasing trend was observed for the increasing GO concentrations. However, the hemolysis percentage was essentially constant with little fluctuation for the negative control. | ||
| GO concentration | Fitting linear equation | Regression coefficient | Linear range [h] |
|---|---|---|---|
| a t represented incubation time. | |||
| Negative control | HI = 0.018t − 0.4532 | 0.2049 | 0–24 |
| Low | HI = 1.3118t − 1.3665 | 0.9730 | 0–10 |
| Mid | HI = 2.4608t + 0.4525 | 0.9941 | 0–10 |
| High | HI = 4.6407t − 2.8564 | 0.9743 | 0–8 |
Since the incubation time was a significant parameter in the hemolysis assay, it was necessary to set a long time (such as 6 h) to objectively assess the hemolytic properties of GO.
3.10 Incubation temperature
The set incubation time was 6 h for all temperatures in this assay on the basis of the results in Section 3.9 Exposure time. The temperature effect was assessed (Fig. 8 and S11†), and the relationship between hemolysis and centrifugation was investigated (Fig. S12†). Hemolysis was correlated with GO concentration and incubation temperatures. | ||
| Fig. 8 The temperature effect on the hemolysis properties of GO after rabbit RBCs were incubated for 6 h. (a) Photographs of RBC hemolysis at 27, 32, 37, 42 and 47 °C. (b) The five hemolysis curves at the different temperatures. From these curves, the distinguishing features of hemolysis properties of GO were closely associated with temperature and GO concentration. The higher the temperature and GO concentration, the more severe the hemolysis. | ||
Temperature was a vital factor in determining RBC fragility, which led to a decline of osmotic fragility and an increase of hemolysis.74 On the one hand, when RBCs transiently expand, cholesterol molecules are excluded from the membrane in protein–lipid boundary regions, and the RBCs are inclined to rupture at this site.75 Substantial intracellular ion leakage has been shown to happen at higher temperature.76 On the other hand, high temperature caused furious thermal motion of the GO nanosheets in suspension, which finally led to an intense interaction between GO and RBCs via the insertion effect of GO into the RBC membranes.
The slope of the hemolysis curve at 47 °C was far more than that at 27 °C (5.11 vs. 0.65, Table 3). The upper limit of the linearity range decreased from 100 to 10 μg mL−1 after a temperature increase from 27 to 47 °C. The results showed that the incubation temperature and GO concentration are major factors, and the insertion effect of GO was rapidly enhanced with temperature.
| Incubation temperature [°C] | Fitting equation | Regression coefficient | Linear range [μg mL−1] |
|---|---|---|---|
| 27 | HI = 0.6512 × CGO + 0.1578 | 0.9331 | 10–100 |
| 32 | HI = 1.2126 × CGO − 1.538 | 0.9779 | 5–50 |
| 37 | HI = 1.7422 × CGO − 4.4399 | 0.9856 | 0–30 |
| 42 | HI = 2.6847 × CGO − 2.2932 | 0.9906 | 0–25 |
| 47 | HI = 5.1089 × CGO − 2.2253 | 0.9888 | 0–10 |
Centrifugation intensified the temperature effect via shear stress (Fig. S12†). The difference curves before and after centrifugation demonstrated that the difference of the hemolysis percentages first increased and then decreased along with GO concentration. The difference value reached a maximum at 27 °C for 50 μg mL−1 GO, and shifted to 25 μg mL−1 GO at 47 °C. The facts showed that centrifugation had a synergistic effect of incubation temperature as well as GO concentration. For the positive control, the hemolysis percentages were nearly irrelevant to incubation temperature. These findings illustrate that 37 °C as the incubation temperature is reasonable and should remain steady in assays.
4. Conclusion
In summary, the calculation method for the hemolytic index proposed in our paper was different from the two traditional methods that caused false-positive errors. The unmodified GO exhibited diverse hemolytic properties in multiple incubation conditions including medium type, RBC species, RBC concentration, storage duration of the RBCs, animal health status, centrifugal speed, exposure time and incubation temperature etc. We could be sure that completely opposite results would be obtained due to the employment of inappropriate incubation conditions. So incubation conditions should be investigated by detailed designs. In our opinion, for unmodified GO, it was optimal to choose isotonic glucose solution as the medium type, fresh and healthy rabbit RBCs as the test object, 1% hematocrit as the RBC concentration, 3000 rpm for 5 min as the centrifugation condition, 6 h as the exposure time and 37 °C as the incubation temperature. Our work was the first investigation of applying various incubation conditions and sample concentrations to intensively study the hemolytic properties of unmodified GO, and these properties have been well explained from the characteristics of GO nanosheets and RBCs. On the whole, unmodified GO nanosheets caused certain damage to RBCs.Conflict of interest
The authors declare no competing financial interest.Acknowledgements
This work was supported by the Natural Science Foundation of Shandong Province, China (No. ZR2015HM032).References
- S. Some, A. R. Gwon, E. Hwang, G. H. Bahn, Y. Yoon, Y. Kim, S. H. Kim, S. Bak, J. Yang, D. G. Jo and H. Lee, Sci. Rep., 2014, 4, 6314 CrossRef CAS PubMed.
- C. Guo, B. Book-Newell and J. Irudayaraj, Chem. Commun., 2011, 47, 12658–12660 RSC.
- H. Wan, Y. Zhang, Z. Liu, G. Xu, G. Huang, Y. Ji, Z. Xiong, Q. Zhang, J. Dong, W. Zhang and H. Zou, Nanoscale, 2014, 6, 8743–8753 RSC.
- X. Y. Xie, K. W. Hu, D. D. Fang, L. H. Shang, S. D. Tran and M. Cerruti, Nanoscale, 2015, 7, 7992–8002 RSC.
- H. Ren, C. Wang, J. Zhang, X. Zhou, D. Xu, J. Zheng, S. Guo and J. Zhang, ACS Nano, 2010, 4, 7169–7174 CrossRef CAS PubMed.
- E. P. Widmaier, H. Raff and K. T. Strang, in Vander’s Human Physiology: The Mechanisms of Body Function, ed. E. P. Widmaier, H. Raff and K. T. Strang, McGraw-Hill, New York, 2011, ch. 12, pp. 359–441 Search PubMed.
- M. A. Dobrovolskaia, J. D. Clogston, B. W. Neun, J. B. Hall, A. K. Patri and S. E. McNeil, Nano Lett., 2008, 8, 2180–2187 CrossRef CAS PubMed.
- S. F. Kiew, L. V. Kiew, H. B. Lee, T. Imae and L. Y. Chung, J. Controlled Release, 2016, 226, 217–228 CrossRef CAS PubMed.
- X. Zhou and F. Liang, Curr. Med. Chem., 2014, 21, 855–869 CrossRef CAS PubMed.
- A. B. Seabra, A. J. Paula, R. de Lima, O. L. Alves and N. Duran, Chem. Res. Toxicol., 2014, 27, 159–168 CrossRef CAS PubMed.
- A. M. Pinto, I. C. Gonçalves and F. D. Magalhães, Colloids Surf., B, 2013, 111, 188–202 CrossRef CAS PubMed.
- American Society for Testing and Materials, Standard practice for assessment of hemolytic properties of materials, ASTM, 2000, F756–00 Search PubMed.
- K. H. Liao, Y. S. Lin, C. W. Macosko and C. L. Haynes, ACS Appl. Mater. Interfaces, 2011, 3, 2607–2615 CAS.
- S. K. Singh, M. K. Singh, P. P. Kulkarni, V. K. Sonkar, J. J. Gracio and D. Dash, ACS Nano, 2012, 6, 2731–2740 CrossRef CAS PubMed.
- C. Cheng, S. Li, S. Nie, W. Zhao, H. Yang, S. Sun and C. Zhao, Biomacromolecules, 2012, 13, 4236–4246 CrossRef CAS PubMed.
- R. Feng, Y. Yu, C. Shen, Y. Jiao and C. Zhou, J. Biomed. Mater. Res., Part A, 2015, 103, 2006–2014 CrossRef CAS PubMed.
- A. He, RSC Adv., 2013, 3, 22120–22129 RSC.
- T. Wang, Toxicol. Res., 2015, 4, 885–894 RSC.
- H. Wu, H. Shi, Y. Wang, X. Jia, C. Tang, J. Zhang and S. Yang, Carbon, 2014, 69, 379–389 CrossRef CAS.
- X. Zhang, J. Yin, C. Peng, W. Hu, Z. Zhu, W. Li, C. Fan and Q. Huang, Carbon, 2011, 49, 986–995 CrossRef CAS.
- A. M. Pinto, C. Gonçalves, D. M. Sousa, A. R. Ferreira, J. A. Moreira, I. C. Gonçalves and F. D. Magalhães, Carbon, 2016, 99, 318–329 CrossRef CAS.
- N. Zhou, H. Gu, F. Tang, W. Li, Y. Chen and Y. Jiang, J. Mater. Sci., 2013, 48, 7097–7103 CrossRef CAS.
- S. Henkelman, G. Rakhorst, J. Blanton and W. Oeveren, Mater. Sci. Eng., C, 2009, 29, 1650–1654 CrossRef CAS.
- X. Pei, J. Wang and Q. Wan, ACS Nano, 2014, 8, 1966 CrossRef CAS PubMed.
- W. S. Hummers and R. E. Offeman, J. Am. Chem. Soc., 1958, 80, 1339 CrossRef CAS.
- M. Shamsipur, F. Molaabasi, M. Shanehsaz and A. A. Moosavi-Movahedi, Microchim. Acta, 2015, 182, 1131–1141 CrossRef CAS.
- G. S. Lemos, L. F. Marquez-Bernardes, L. R. Arvelos, L. F. Paraiso and N. Penha-Silva, Cell Biochem. Biophys., 2011, 61, 531–537 CrossRef CAS PubMed.
- D. Alarcon-Segovia, A. Ruiz-Arguelles and L. Llorente, J. Immunol., 1979, 122, 1855–1862 CAS.
- A. Bersényi, S. G. Fekete, Z. Szöcs and E. Berta, Acta Vet. Hung., 2003, 51, 297–304 CrossRef PubMed.
- K.-c. Chan, M.-c. Yin and W.-j. Chao, Food Chem. Toxicol., 2007, 45, 502–507 CrossRef CAS PubMed.
- L. A. Maginniss, J. M. Szewczak and C. M. Troup, Environ. Health Perspect., 1987, 72, 35–38 CrossRef CAS PubMed.
- D. Chalfin, J. Cell. Comp. Physiol., 1956, 47, 215–243 CrossRef CAS.
- J. Shang, L. Ma, J. Li, W. Ai, T. Yu and G. G. Gurzadyan, Sci. Rep., 2012, 2, 792 Search PubMed.
- K. N. Kudin, B. Ozbas, H. C. Schniepp, R. K. Prud’homme, I. A. Aksay and R. Car, Nano Lett., 2008, 8, 36–41 CrossRef CAS PubMed.
- S. C. Lee, S. Some, S. W. Kim, S. J. Kim, J. Seo, J. Lee, T. Lee, J. H. Ahn, H. J. Choi and S. C. Jun, Sci. Rep., 2015, 5, 12306 CrossRef PubMed.
- R. E. Waugh, M. Narla, C. W. Jackson, T. J. Mueller, T. Suzuki and G. L. Dale, Blood, 1992, 79, 1351–1358 CAS.
- R. L. Whitby, V. M. Gun’ko, A. Korobeinyk, R. Busquets, A. B. Cundy, K. Laszlo, J. Skubiszewska-Zieba, R. Leboda, E. Tombacz, I. Y. Toth, K. Kovacs and S. V. Mikhalovsky, ACS Nano, 2012, 6, 3967–3973 CrossRef CAS PubMed.
- M. Y. Kiriukhin and K. D. Collins, Biophys. Chem., 2002, 99, 155–168 CrossRef CAS PubMed.
- D. Arosio, H. E. Kwansa, H. Gering, G. Piszczek and E. Bucci, Biopolymers, 2002, 63, 1–11 CrossRef CAS PubMed.
- E. K. Smith, D. Farrington and L. Sydiuk, Can. J. Physiol. Pharmacol., 1972, 50, 791–797 CrossRef CAS PubMed.
- Y. Tu, M. Lv, P. Xiu, T. Huynh, M. Zhang, M. Castelli, Z. Liu, Q. Huang, C. Fan, H. Fang and R. Zhou, Nat. Nanotechnol., 2013, 8, 594–601 CrossRef CAS PubMed.
- S. P. Koenig, N. G. Boddeti, M. L. Dunn and J. S. Bunch, Nat. Nanotechnol., 2011, 6, 543–546 CrossRef CAS PubMed.
- E. Condrea, Z. Mammon, S. Aloof and A. de Vries, Biochim. Biophys. Acta, 1964, 84, 365–375 CAS.
- G. J. Nelson, Biochim. Biophys. Acta, 1967, 144, 221–232 CrossRef CAS.
- J. De Gier and L. Van Deenen, Biochim. Biophys. Acta, 1961, 49, 286–296 CrossRef.
- R. A. Demiel, W. S. Guerts van Kessel and L. L. van Deenen, Biochim. Biophys. Acta, 1972, 266, 26–40 CrossRef CAS.
- A. M. Forsyth, S. Braunmüller, J. Wan, T. Franke and H. A. Stone, Microvasc. Res., 2012, 83, 347–351 CrossRef CAS PubMed.
- C. Sardet, H. Hansma and R. Ostwald, J. Lipid Res., 1972, 13, 705–715 CAS.
- P. Montes-Navajas, N. G. Asenjo, R. Santamaria, R. Menendez, A. Corma and H. Garcia, Langmuir, 2013, 29, 13443–13448 CrossRef CAS PubMed.
- C. G. Malmberg and A. A. Maryott, J. Res. Natl. Bur. Stand., 1950, 45, 299–303 CrossRef CAS.
- B. Cai, K. Hu, C. Li, J. Jin and Y. Hu, Appl. Surf. Sci., 2015, 356, 844–851 CrossRef CAS.
- Y. Wang, A. G. El-Deen, P. Li, B. H. L. Oh, Z. Guo, M. M. Khin, Y. S. Vikhe, J. Wang, R. G. Hu, R. M. Boom, K. A. Kline, D. L. Becker, H. Duan and M. B. Chan-Park, ACS Nano, 2015, 9, 10142–10157 CrossRef CAS PubMed.
- G. N. Smith and P. L. Mollison, Scand. J. Haematol., 1973, 11, 188–194 CrossRef CAS PubMed.
- R. L. Jones and C. M. Peterson, Am. J. Med., 1981, 70, 339–352 CrossRef CAS PubMed.
- H. Resmi, H. A. A. Akhunlar and G. Guner, Cell Biochem. Funct., 2005, 23, 163–168 CrossRef CAS PubMed.
- K. J. Davies, J. Biol. Chem., 1987, 262, 9895–9901 CAS.
- J. W. Baynes, Diabetes, 1991, 40, 405–412 CrossRef CAS PubMed.
- I. Buhler, R. Walter and W. H. Reinhart, Eur. J. Clin. Invest., 2001, 31, 79–85 CrossRef CAS PubMed.
- L. Coppola, G. Verrazzo, C. La Marca, P. Ziccardi, A. Grassia, A. Tirelli and D. Giugliano, Diabetic Med., 1997, 14, 959–963 CrossRef CAS PubMed.
- V. P. Patel, A. Ciechanover, O. Platt and H. F. Lodish, Proc. Natl. Acad. Sci. U. S. A., 1985, 82, 440–444 CrossRef CAS.
- J. Balkan, S. Oztezcan, G. Aykac-Toker and M. Uysal, Biosci., Biotechnol., Biochem., 2002, 66, 2701–2705 CrossRef CAS PubMed.
- C. L. Balduini, G. Ricevuti, M. Sosso, E. Ascari and C. Balduini, Hoppe Seylers Z. Physiol. Chem., 1977, 358, 1143–1148 CrossRef CAS PubMed.
- L. M. Snyder, G. Fairbanks, J. Trainor, N. L. Fortier, J. B. Jacobs and L. Leb, Br. J. Haematol., 1985, 59, 513–522 CrossRef CAS PubMed.
- E. R. Jaffe, G. A. Vanderhoff, B. A. Lowy and I. M. London, J. Clin. Invest., 1958, 37, 1293–1297 CrossRef CAS PubMed.
- J. R. Durocher, R. C. Payne and M. E. Conrad, Blood, 1975, 45, 11–20 CAS.
- L. B. Leverett, J. D. Hellums, C. P. Alfrey and E. C. Lynch, Biophys. J., 1972, 12, 257–273 CrossRef CAS PubMed.
- M. Guo, D. Li, M. Zhao, Y. Zhang, D. Geng, A. Lushington and X. Sun, Carbon, 2013, 61, 321–328 CrossRef CAS.
- X. U. Dong, N. L. Zhou and J. Shen, Chem. Res. Chin. Univ., 2010, 31, 2354–2359 Search PubMed.
- Y. Xiao, Y. Fan, W. Wang, H. Gu, N. Zhou and J. Shen, Drug Delivery, 2014, 21, 362–369 CrossRef CAS PubMed.
- J. Tan, N. Meng, Y. Fan, Y. Su, M. Zhang, Y. Xiao and N. Zhou, Mater. Sci. Eng., C, 2016, 61, 681–687 CrossRef CAS PubMed.
- N. Meng, S. Q. Zhang, N. L. Zhou and J. Shen, Nanotechnology, 2010, 21, 185101 CrossRef PubMed.
- S. Thampi, V. Muthuvijayan and R. Parameswaran, J. Appl. Polym. Sci., 2015, 132, 41809 CrossRef.
- M. Papi, M. C. Lauriola, V. Palmieri, G. Ciasca, G. Maulucci and M. De Spirito, RSC Adv., 2015, 5, 81638–81641 RSC.
- G. V. Richieri and H. C. Mel, Biochim. Biophys. Acta, 1985, 813, 41–50 CrossRef CAS.
- B. Aloni, A. Eitan and A. Livne, Biochim. Biophys. Acta, 1977, 465, 46–53 CrossRef CAS.
- P. Seeman, T. Sauks, W. Argent and W. O. Kwant, Biochim. Biophys. Acta, 1969, 183, 476–489 CrossRef CAS.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c6ra13607a |
| This journal is © The Royal Society of Chemistry 2016 |