
Punica granatum (pomegranate) peel extract exerts potent antitumor and anti-metastasis activity in thyroid cancer
Abstract
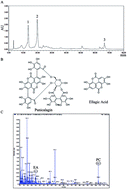
The incidence of thyroid carcinoma has obviously been rising throughout the world during the past ten years. However, over-treatment has usually occurred in thyroid carcinoma without new and effective approaches being explored. In this study, Punica granatum (pomegranate) peel extract (PoPx), a kind of herb, was evaluated for its anticancer activity to thyroid carcinoma in vitro and in vivo. PoPx potently suppressed proliferation in two kinds of thyroid cancer cell lines, and induced cancer cell apoptosis. PoPx could also decrease the mitochondrial membrane potential (ΔΨm), indicating that PoPx may induce apoptosis via a mitochondria-mediated apoptotic pathway. In addition, PoPx markedly impaired thyroid cancer cell migration and invasion by down-regulating the expression of matrix metalloproteinase-9 (MMP-9). More importantly, PoPx significantly inhibited tumor growth in the BCPAP-bearing mice model by reducing cell proliferation and inducing apoptosis. These findings suggest that PoPx could be an effective phytochemical agent.
 Please wait while we load your content...
Please wait while we load your content...

