
Insight into the short-term effect of titanium dioxide nanoparticles on active ammonia oxidizing microorganisms in a full-scale wastewater treatment plant: a DNA-stable isotope probing study
Jing-Feng Gao*,
Xiao-Yan Fan,
Xin Luo and
Kai-Ling Pan
College of Environmental and Energy Engineering, Beijing University of Technology, Beijing 100124, China. E-mail: gao.jingfeng@bjut.edu.cn; gao158@gmail.com; Fax: +86-10-67391983; Tel: +86-10-67391918
First published on 28th July 2016
Abstract
Ammonia-oxidizing bacteria (AOB) and archaea (AOA) are two distinct ammonia-oxidizing microorganisms (AOMs) responsible for nitrification in wastewater treatment plants (WWTPs). However, their relative contributions to ammonia oxidation in WWTPs and short-term responses to titanium dioxide nanoparticles (TiO2 NPs) remain unclear. In this study, DNA-based stable isotope probing (DNA-SIP), quantitative polymerase chain reaction (qPCR), PCR, cloning and sequencing were applied to investigate the in situ activity of AOMs in a full-scale WWTP and to evaluate their responses to TiO2 NPs. An environmentally relevant concentration (1 mg L−1) and a higher concentration (50 mg L−1) of TiO2 NPs were chosen for laboratory microcosms. The laboratory microcosms revealed that TiO2 NPs (1 or 50 mg L−1) caused slight or no short-term inhibitions on nitrification activity and nitrification rate of activated sludge. The successful Na213CO3 assimilation of AOB rather than AOA in original sludge provided compelling evidence for the strong contribution of AOB to the in situ nitrification of the WWTP. The main AOB in original sludge, Nitrosomonas oligotropha cluster and Nitrosomonas europaea cluster, were the dominant active AOB in the WWTP. However, compared with the original sludge, the abundance and distribution of active AOB changed under 1 and 50 mg L−1 of TiO2 NPs. In particular, AOA within Nitrososphaera cluster were the active AOMs under 50 mg L−1 TiO2 NPs. These results suggested that TiO2 NPs had a potential impact on the abundance and composition of active AOMs in the WWTP. This study further provided direct evidence of the autotrophic growth of AOB within N. oligotropha cluster and N. europaea cluster, and AOA within the general Nitrososphaera cluster. Overall, AOB played an important role in the in situ nitrification of the full-scale WWTP investigated, while AOA showed a strong contribution to the active nitrification under higher concentration of TiO2 NPs.
1. Introduction
In general, ammonia-oxidizing bacteria (AOB) as well as ammonia-oxidizing archaea (AOA) discovered in 2005 (ref. 1 and 2) are responsible for ammonia oxidation, the first and rate-limiting step of nitrification. The phylogenetic diversity and abundance of the two ammonia-oxidizing microorganisms (AOMs) have been extensively investigated in terrestrial systems,3 estuarine and ocean sediments,2,4 groundwater systems,5 drinking water treatment plants6 and wastewater treatment plants (WWTPs).7–9 Especially, AOB and AOA are important for nitrogen removal in WWTPs. Phylogenetic analyses based on 16S rRNA and amoA genes demonstrate that AOB in WWTPs mainly belong to Beta-proteobacteria, including Nitrosomonas and Nitrosospira.10–12 AOA in WWTPs are predominated by Nitrososphaera cluster, while Nitrosopumilus cluster occurs only sporadically in these systems, except the saline nitrification reactors.11,13–15 Moreover, the previous studies have provided quantitative data on AOB and AOA amoA genes in municipal and industrial WWTPs, and the results suggest that there is a variation in their abundances.11–13,16,17 Nevertheless, in general, most of the studies indicate that AOB are numerically dominant in the municipal WWTPs.11,12,16 However, the current knowledge of phylogenetic diversity and abundance of AOMs provides few direct evidences regarding their metabolic capacities. Therefore, new molecular technologies should be applied to gain a deep insight into the activity of AOMs in WWTPs.DNA based stable-isotope probing (DNA-SIP) is a powerful technique to link the active microorganisms with a specific metabolic process.18 Microcosms are used to incubate the environmental samples with stable-isotope-labeled substrates.19 13C-labeled substrates have been most commonly used in microcosms. After incubation, DNA is isolated from the environmental samples, and isopycnic centrifugation is performed in a CsCl gradient to separate 12C- and 13C-DNA for downstream analyses.20 13C-DNA-SIP approach can provide direct evidence for active AOMs based on the fact that both AOA and AOB can use inorganic carbon as carbon source to perform ammonia oxidation. So far, 13C-DNA-SIP has been applied to identify the active AOMs in sediments,21 agriculture soils,22,23 acid soils,24–26 paddy soils,27,28 dry subhumid ecosystems29 and granular activated carbon used for drinking water purification.30 However, DNA-SIP has not been employed to investigate the activity of AOMs and to assess their relative contributions to ammonia oxidation in full-scale WWTPs.
Nanoparticles (NPs) are materials within the dimension of 1 to 100 nm with novel physicochemical characteristics, such as composition, size, shape, surface charge and capping molecules, etc. These properties may be toxic for microorganisms, and therefore the large utilization of NPs will induce the potential risks to human and other living microorganisms.31 The commonly mechanisms of toxicity of NPs include oxidative stress via the generation of reactive oxygen species on crystal surfaces of NPs, the release of metal ions from the dissolution of metals from the surface of the NPs, the penetration of the cell envelope, and disorganization of bacterial membrane.32–34
Moreover, the extensive applications of NPs, especially titanium dioxide nanoparticles (TiO2 NPs) in various consumer and industrial products have caused an observation of TiO2 NPs in full-scale WWTPs.35 The prediction of TiO2 NPs concentrations in wastewater effluents is at μg L−1 levels (2–20 μg L−1),35 but the concentrations may continuously increase due to their large production and application. Adsorption to activated sludge seems to be the major removal mechanism for NPs.36 Usually, 90–95% of TiO2 NPs could be accumulated in activated sludge in WWTPs.37
Recent years, the growing concerns have been raised on long-term and short-term effects of TiO2 NPs on the performance of WWTPs.38–41 It was reported in the literature that after long-term exposure (70 days) to 50 mg L−1 TiO2 NPs, the nitrogen removal efficiency is significantly inhibited.38,39 Nevertheless, the previous study demonstrates that long-term (24 days) exposures to TiO2 NPs (1, 10 and 100 mg L−1) do not adversely affect the nitrogen removal.42 The gradually increase of TiO2 NPs concentrations from 10 to 50 mg L−1 was observed to have limit adverse effect on nitrification efficiency after long-term exposure (100 days).41 These results suggest that the long-term effect of TiO2 NPs on nitrogen removal is different. However, the previous studies suggest that TiO2 NPs have no acute effects on wastewater nitrogen removal or ammonia oxidation after short-term exposure. Short-term (1 day) exposure to TiO2 NPs (1, 10, 50 and 100 mg L−1) do not adversely affect the nitrogen removal.38,42 The increase of TiO2 NPs concentrations from 2 to 50 mg L−1 also do not affect the nitrogen removal after short-term exposure (7 days).40 Moreover, the potential toxicity of TiO2 NPs to communities of activated sludge has been investigated. Long-term exposure to 50 mg L−1 TiO2 NPs significantly reduce the microbial diversity of activated sludge, the abundance of functional genera (especially AOB) and the catalytic activity of essential enzymes.38,39 However, the microbial community did not significantly vary after short-term exposure.42 A previous study indicated that the difference caused by NPs largely depends on exposure time rather than NPs concentrations.42 Overall, after short-term exposure to TiO2 NPs, although the ammonia oxidation or nitrogen removal are not or slightly inhibited, the active microorganisms responsible for this process may be different. Until now, the short-term effect of TiO2 NPs on the active AOA and AOB in full-scale WWTPs remains uncertain.
Given the above arguments, one objective of this study was to investigate the in situ activity of AOA and AOB in a full-scale WWTP. Another objective was to evaluate the short-term effects of different TiO2 NPs concentrations on nitrification activity and abundance and diversity of active AOMs. The environmentally relevant concentration (1 mg L−1) and a higher concentration of TiO2 NPs (50 mg L−1) were chosen for the study. By setting up short-term laboratory microcosms of activated sludge under different TiO2 NPs concentrations, 13C-DNA-SIP technique would provide direct evidences to the in situ activity of AOA and AOB and their responses to TiO2 NPs. Quantitative PCR (qPCR), PCR, cloning and sequencing were applied to track the active AOB and AOA. Moreover, this study might provide evidence for autotrophic growth of the active AOMs with Na213CO3 as the labeled substrate in the microcosms.
2. Materials and methods
2.1. Nanoparticle suspension
Commercially produced TiO2 NPs used in this study were purchased from Sigma-Aldrich (St. Louis, MO, USA). The specific surface area of TiO2 NPs was 323 m2 g−1 according to the BET method. The stock suspensions (100 and 1000 mg L−1) were prepared by dispersing 10 mg and 500 mg of TiO2 NPs in 100 mL and 500 mL Milli-Q water, respectively, followed by 1 h of sonication (25 °C, 250 W, 40 kHz).432.2. DNA-SIP microcosms at different TiO2 NPs concentrations
Fig. 1 depicts the experimental flow diagram of this study. Activated sludge sample was collected from the outer channel of a Carrousel® 3000 process in a full-scale WWTP (40.05° N, 116.26° E) in Beijing, China during July. The relevant parameters and descriptions are as follows: treatment capacity, 2 × 104 m3 d−1; dissolved oxygen (DO), 0.2–0.3 mg L−1 in outer channel, 0.5–1.5 mg L−1 in middle channel and 1.5–2.5 mg L−1 in inner channel; influent NH4+-N, 20 mg L−1; influent chemical oxygen demand (COD), 154 mg L−1; effluent NH4+-N, 2 mg L−1; influent COD, 49 mg L−1; average mixed liquid suspended solids (MLSS), 5000 mg L−1; sludge retention time (SRT), 15 d; pH, 7.4–8.2. Here, the values of NH4+-N, COD and MLSS were the monthly average. AOB and AOA were confirmed to be present in the WWTP, and their abundance and diversity have been investigated in the previous study.11 AOB (3.96 × 108 copies per g sludge) outnumber AOA (1.73 × 105 copies per g sludge) in original sludge. The detected AOB lineages in original sludge are Nitrosomonas oligotropha cluster (70.01%) Nitrosomonas-like cluster (19.93%) and Nitrosomonas europaea cluster (10.06%). AOA in the original sludge are general Nitrososphaera (76.78%) and Nitrososphaera subcluster 4.1 (23.22%). After sampling, the activated sludge was immediately kept inside of icebox during transportation to the lab. | ||
| Fig. 1 Experimental flow diagram of this study. Duplicate microcosms were conducted in the presence of different concentrations of TiO2 NPs. | ||
Before incubation of laboratory microcosms, the activated sludge was pre-incubated in a 2 L beaker to remove organic matter and ammonia at 25 °C (in situ temperature of sampling). The DO was controlled at 2 mg L−1. During the pre-incubation, supernatant was collected for every 30 min and filtered through 0.2 μm pore size polytetrafluoroethylene membranes. The concentration of ammonia was analyzed immediately using UV-vis spectrophotometry (UV-2802PC, Unico, Shanghai, China) in accordance with standard methods.44 The activated sludge showed good nitrification activity, and after about one hour of pre-incubation, the ammonia concentration decreased to zero. The activated sludge was immediately used for the laboratory microcosms.
The TiO2 NPs concentrations chosen for DNA-SIP microcosms were 0, 1 and 50 mg L−1 according to the literature.38 12C-Na2CO3 and 13C-Na2CO3 were used as the control and labeled substrates in DNA-SIP microcosms. The 13C-Na2CO3 with 99% purity was purchased from Cambridge Isotope Laboratories, Inc. DNA-SIP microcosm was conducted in duplicate to investigate the in situ activity of AOA and AOB in the WWTP. In total, there were 12 activated sludge microcosms. The laboratory microcosms were performed in 250 mL conical flasks containing 25 mL activated sludge and 100 mL synthetic wastewater at 25 °C under agitation at 100 rpm. The conical flasks were named as 12-0, 13-0, 12-1, 13-1, 12-50 and 13-50 according to the different TiO2 NPs concentrations (Fig. 1). The DO and MLSS of different microcosms were controlled at 2 and 5000 mg L−1, respectively. In this study, in order to keep the activated sludge in the in situ condition, eight cycles were selected for all the microcosms. At the first cycle, the incubation time of each microcosm was determined by measuring the ammonia, nitrite and nitrate concentrations of supernatant for every 30 min. Indicator of the ending of one cycle was that the ammonia concentration of the supernatant was close to zero. Therefore, the target concentration of ammonia could be maintained at constant level over the entire incubation period. Aeration of the microcosms was stopped at the end of each cycle. After setting, 100 mL supernatant was removed from the conical flasks and replaced with 100 mL new synthetic wastewater at the beginning of next microcosm. For the cycles ranged from two to seven, the ammonia, nitrite and nitrate concentrations of influent and effluent of each microcosm were regularly measured. Finally, in order to investigate the potential impact of TiO2 NPs on nitrification rate, the ammonia, nitrite and nitrate concentrations of the eighth cycle were measured every 30 min.
The synthetic wastewater contained: NH4Cl (0.075 g L−1, 1.4 mM), NaCl (0.585 g L−1, 10 mM), KH2PO4 (0.054 g L−1, 0.4 mM), KCl (0.075 g L−1, 1 mM), CaCl2·2H2O (0.147 g L−1, 1 mM), MgSO4·7H2O (0.049 g L−1, 0.2 mM), Na2CO3 (0.1484 g L−1, 1.4 mM) and trace element (1 mL L−1).45 The trace element contained: Na2EDTA (4292 mg L−1, 11.5 mM), FeCl2·4H2O (1988 mg L−1, 10 mM), MnCl2·2H2O (81 mg L−1, 0.5 mM), NiCl2·6H2O (24 mg L−1, 0.1 mM), CoCl2·6H2O (24 mg L−1, 0.1 mM), CuCl2·2H2O (17 mg L−1, 0.1 mM), ZnCl2 (68 mg L−1, 0.5 mM), Na2MoO4·2H2O (24 mg L−1, 0.1 mM), Na2WoO4·2H2O (33 mg L−1, 0.1 mM), H3BO3 (62 mg L−1, 1 mM).46
2.3. DNA extraction and SIP gradient fractionation
Activated sludge samples were collected at the end of DNA-SIP microcosms. Samples collected from the duplicate microcosms were mixed prior to be freeze-dried by using Labconco Freezone 1 L (Labconco, USA). Therefore, six samples were obtained for subsequent experiments. DNA was extracted from 0.05–0.10 g of dried activated samples with a Fast DNA® SPIN Kit for soil (Qiagen, CA, USA) following the manufacturer's protocol. Concentrations of the DNA extracted were measured by Nanodrop-1000 (Thermo Fisher Scientific, USA).Isopycnic centrifugation was performed in a CsCl gradient for the separation of 12C- and 13C-DNA in the microcosms followed the study of Neufeld et al.20 with slight modifications. Briefly, extracted DNA (5 μg) was combined into a CsCl solution (7.163 M) and a gradient buffer (0.1 M Tris–HCl, pH 8.0; 1 mM EDTA; 0.1 M KCl) to achieve an initial buoyant density of 1.72 g mL−1. Ultracentrifugation of the mixed solution was performed in 5.1 mL OptiSeal polyallomer tubes with a VTi 90 vertical rotor (Optima L-100XP Ultracentrifuge; Beckman Coulter, Palo Alto, CA, USA) at 190![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 × g for 44 h at 20 °C. Fifteen equal DNA fractions (∼300 μL) were obtained by displacing the gradient medium with sterile water from the top of the ultracentrifuge tube using a syringe pump (LSP01-1A, Baoding Longer Precision Pump Co., Ltd). After fractionation, 70 μL aliquot of each fraction was used to measure the buoyant density with an AR200 digital hand-held refractometer (Reichert, Inc., Buffalo, NY, USA). Then, the buoyant densities of the 15 fractions were calculated by their refractive indices. The fractionated DNA was separated from CsCl solution by polyethylene glycol 6000 precipitation at 37 °C for 1 h followed by centrifugation at 13000 × g for 30 min. The precipitated DNA was purified with 70% ethanol for two times and then dried at room temperature for about 1 h. Finally, the precipitated DNA was dissolved in 30 μL Tris–EDTA buffer solution and stored at −20 °C. The DNA obtained from fraction 4 to 7 and 10 to 13 was defined as “heavy” and “light” DNA, respectively.
000 × g for 44 h at 20 °C. Fifteen equal DNA fractions (∼300 μL) were obtained by displacing the gradient medium with sterile water from the top of the ultracentrifuge tube using a syringe pump (LSP01-1A, Baoding Longer Precision Pump Co., Ltd). After fractionation, 70 μL aliquot of each fraction was used to measure the buoyant density with an AR200 digital hand-held refractometer (Reichert, Inc., Buffalo, NY, USA). Then, the buoyant densities of the 15 fractions were calculated by their refractive indices. The fractionated DNA was separated from CsCl solution by polyethylene glycol 6000 precipitation at 37 °C for 1 h followed by centrifugation at 13000 × g for 30 min. The precipitated DNA was purified with 70% ethanol for two times and then dried at room temperature for about 1 h. Finally, the precipitated DNA was dissolved in 30 μL Tris–EDTA buffer solution and stored at −20 °C. The DNA obtained from fraction 4 to 7 and 10 to 13 was defined as “heavy” and “light” DNA, respectively.
2.4. qPCR of AOA and AOB amoA genes
The abundances of AOA and AOB amoA genes in all the fractions were quantified by qPCR on a Stratagene MX3005p thermocycler (Agilent Technologies, USA) using a Fast-Plus EvaGreen Master Mix (Biotium, USA). The qPCR was performed in triplicate. The primer sets for AOA and AOB were Arch-amoAF/Arch-amoAR2 and amoA-1F/amoA-1R,47 respectively. The 20 μL qPCR mixture contained aseptic water (7.2 μL), each primer (0.4 μL, 10 μM), qPCR mix (10 μL) and template DNA (2 μL, ∼5 ng). For AOA amoA gene, the qPCR conditions were as follows: 96 °C for 2 min, 40 cycles of 5 s at 96 °C, 20 s at 56 °C and 25 s at 72 °C. For AOB amoA gene, the qPCR conditions were the same as AOA amoA gene except the different annealing temperature (57 °C). The standard curve was generated by using 10-fold serial dilutions of plasmid DNA. For AOA and AOB, plasmids containing the target amoA gene fragments with accession number JX233564 and JX233554 were selected as the standard DNA. For standard curve of AOA, the amplification efficiencies of 93.8% to 97.9% were obtained, with R2 values of 0.972 to 1.000. Amplification efficiencies for the standard curve of AOB were between 93.4% and 104.6%, with R2 values of 0.987 to 0.998.2.5. PCR amplification of AOA and AOB amoA genes
Each fraction of all the samples collected from DNA-SIP microcosms was used to amplify AOA and AOB amoA genes with primer sets Arch-amoAF/Arch-amoAR and amoA-1F/amoA-2R, respectively. The 25 μL PCR reaction mixture comprised of 12.5 μL of GoTaq® Green Master Mix (Promega, USA), 0.5 μL of each primer (10 μM), 9.5 μL of aseptic water and 2 μL of template DNA (∼5 ng). For AOA amoA gene, PCR amplification was conducted under the following conditions: an initial 2 min at 95 °C, followed by 36 cycles of 45 s of denaturation at 95 °C, 1 min of annealing at 55 °C and 1 min of extension at 72 °C, with a final extension at 72 °C for 10 min. The PCR conditions for AOB were the same as AOA except the different annealing temperature (57 °C). The PCR products were detected by 1.5% of agarose gel electrophoresis.2.6. Cloning, sequencing and phylogenetic analyses of AOA and AOB amoA genes
PCR products of the “heavy” fractions of each microcosm were purified and ligated to pGEM®-T Easy Vector System (Promega, Madison, WI, USA). The Escherichia coli JM109 competent cells (Takara, Dalian, China) were used for clone library construction. For AOA amoA gene, only one library was successfully constructed and 16 sequences were obtained. For AOB amoA gene, two libraries were constructed and 52 sequences were obtained. The sequences obtained were aligned with MEGA 5.0 software and then were compared with those in the database of the National Center for the Biotechnology Information (NCBI). The identified AOA or AOB amoA gene sequences were then divided into different operational taxonomic units (OTUs) with the 97% sequence similarity using Mothur 1.28.48 The neighbor-joining (NJ) phylogenetic tree was constructed using MEGA 5.0 based on the sequences obtained and the reference sequences.2.7. Accession numbers
All amoA gene sequences obtained were deposited in GenBank. For AOA, the accession numbers were KC700852 to KC700855 and KX113362 to KX113373. For AOB, the accession numbers were KC700931 to KC700964 and KC700984 to KC701001.3. Results and discussion
3.1. Nitrification activity in activated sludge under different TiO2 NPs concentrations
To assess nitrification activity, ammonia, nitrite and nitrate concentrations in the influents and effluents of each cycle in all the activated sludge microcosms were measured. As shown in Fig. 2, for all the control and labeled microcosms, the average ammonia concentration in the synthetic wastewater was (20.42 ± 2.74) mg L−1 and most of ammonia was transformed into nitrate. Concentrations of nitrate ranged from (16.29 ± 4.86) to (18.34 ± 3.95) mg L−1. Productions of nitrite and nitrate were close for all the control and labeled microcosms, indicating the similar active nitrification activity under environmentally relevant concentration (1 mg L−1) and a higher concentration (50 mg L−1) of TiO2 NPs. The results suggested that TiO2 NPs caused a slight or no short-term inhibition on nitrification activity.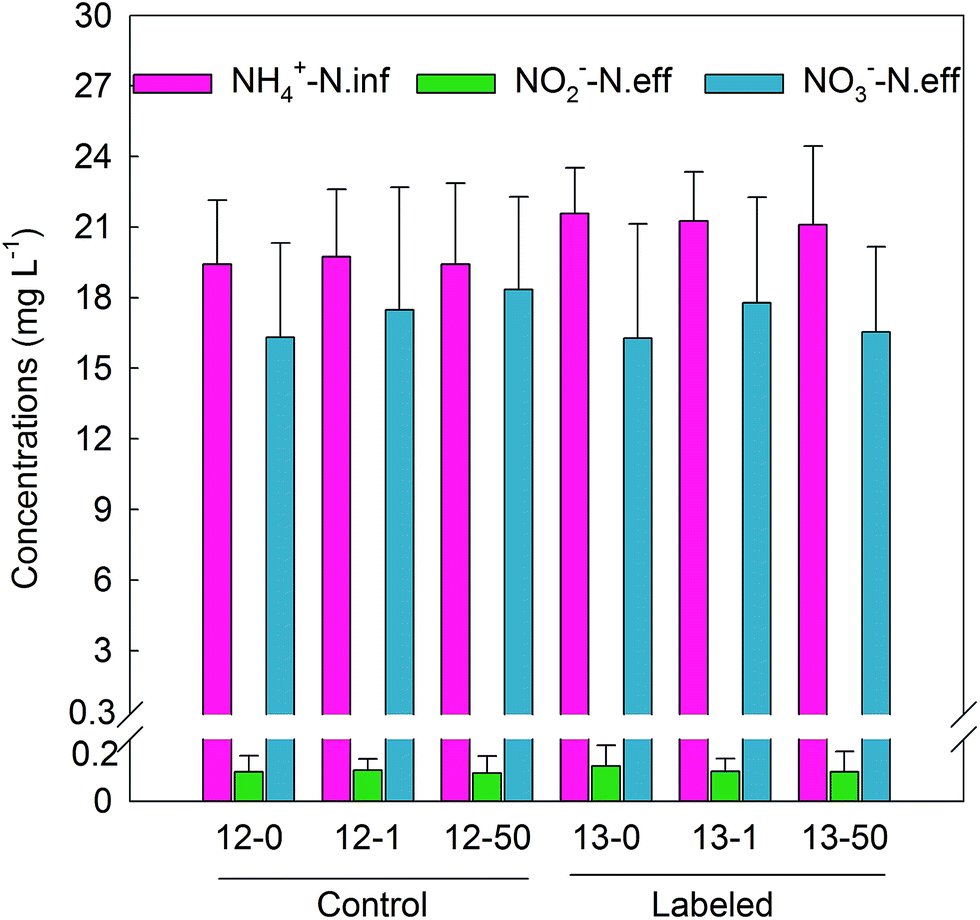 | ||
| Fig. 2 Changes in ammonia, nitrite and nitrate concentrations under different TiO2 NPs concentrations for control and labeled microcosms. In order to keep the activated sludge in the in situ condition, eight cycles were selected for all the microcosms. Data plotted are means and standard deviations of influent ammonia and effluent nitrite and nitrate concentrations of eight cycles for each microcosm. | ||
In order to investigate the potential impact of TiO2 NPs on nitrification rate, the variations of ammonia, nitrite and nitrate among the first and eighth cycle of the control microcosms were measured (Fig. 3). As shows in Fig. 3, for both cycle 1 and cycle 8, similar variation trends of ammonia concentration were observed in the presence of 0, 1 and 50 mg L−1 TiO2 NPs. Namely, the ammonia concentrations decreased slowly for the first 30 min, sharply decreased from 30–60 min and kept slightly decrease for the last 30 min. Also, the variation trends of nitrite and nitrate concentrations were similar under 0, 1 and 50 mg L−1 TiO2 NPs for both cycle 1 and cycle 8. The nitrate concentrations increased slowly for the first 30 min, and which showed a rapid increase for the next 30 min and a slight increase for the last 30 min. Furthermore, the nitrification rates of each treatment were calculated. Close nitrification rates were obtained, and the average nitrification rates of the two cycles were 0.044, 0.042 and 0.044 (NH4+-N) [g(MLSS) min]−1 at 0, 1 and 50 mg L−1 TiO2 NPs, respectively. These results suggested that nitrification rates were slightly or not inhibited by TiO2 NPs of the environmentally relevant concentration (1 mg L−1) and higher concentration (50 mg L−1) after short-term exposure. In summary, TiO2 NPs had slight or no short-term effects on nitrification activity, which were in accordance with previous studies in activated sludge.38,40 TiO2 NPs with the same concentrations (1 and 50 mg L−1) as this study were confirmed to have no effects of nitrogen removal after only one day exposure in a SBR.38 The 2–50 mg L−1 of TiO2 NPs also did not affect the nitrogen removal after seven days exposure in two SBRs.40 One possible reason for this phenomena was related to the dispersion and aggregation of TiO2 NPs with different concentrations. Sonication was used for the preparation of TiO2 NPs suspensions, which is an effective method for eliminating aggregation of NPs and could ensure TiO2 NPs in good dispersion state.49 However, TiO2 NPs have a high tendency for aggregation, and their aggregations would become more in quantity and larger in size with the increase of TiO2 NPs concentrations.50 Therefore, compared with 1 mg L−1 of TiO2 NPs, despite the higher concentration, the aggregation of TiO2 NPs in the presence of 50 of mg L−1 TiO2 NPs would largely reduce the effective surface area, surface reactivity and critical adverse impacts on cells of TiO2 NPs, leading to the similar nitrification activity and nitrification rates. However, a previous study suggest that nitrification functionality was not impacted by dosing NPs (TiO2 NPs) or corresponding bulk material (bulk TiO2), indicating that the state of NPs has no effect on nitrification.51 Further investigations about the effect of dispersion and aggregation of TiO2 NPs on nitrification are needed. Moreover, a previous study suggests that the effects caused by NPs are largely depend on exposure time rather than NPs concentration.42 Therefore, in this study, the short-term incubation (eight cycles) was probably one reason for that TiO2 NPs (1 and 50 mg L−1) did not inhibit the nitrification activity and nitrification rates.
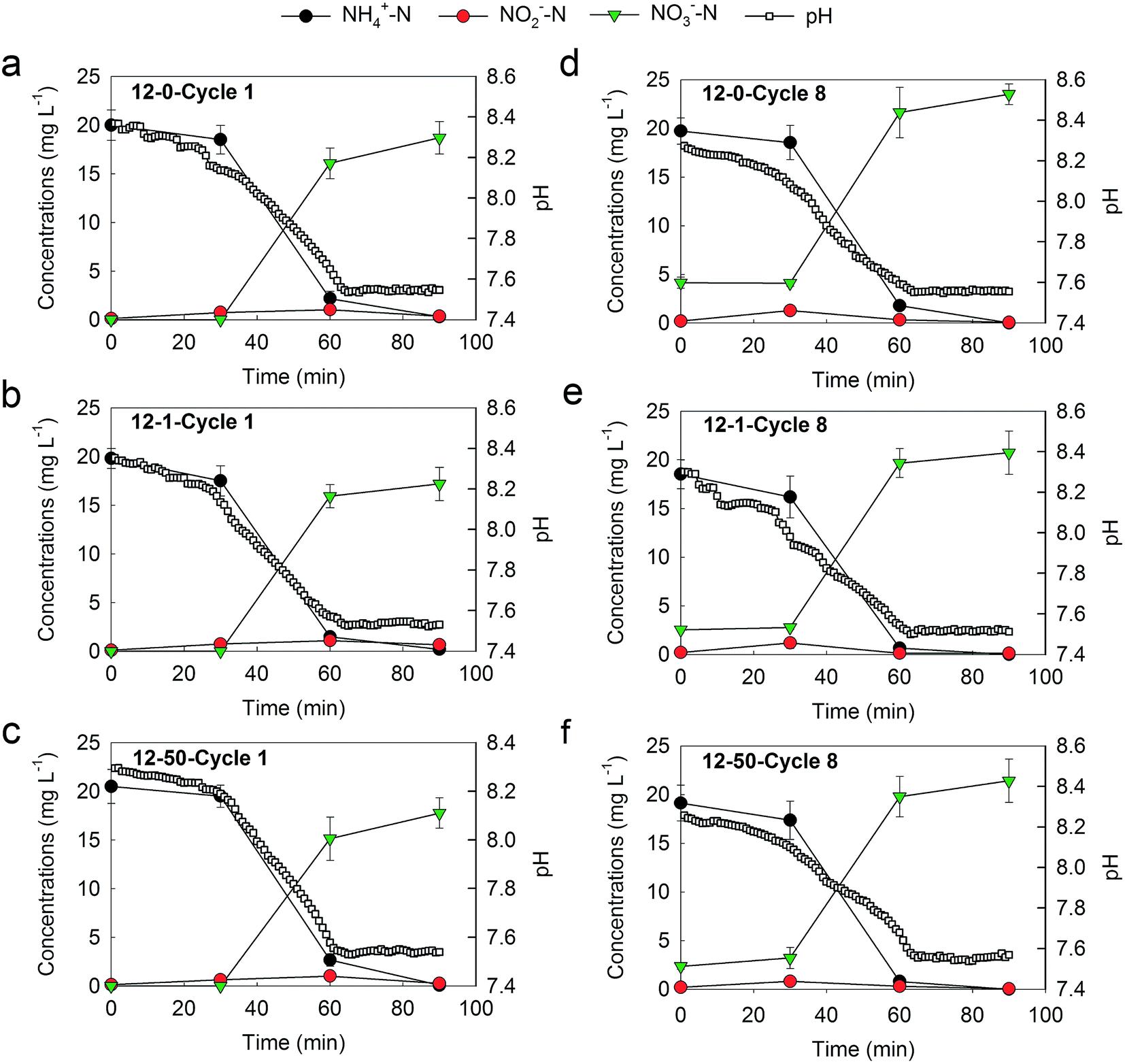 | ||
| Fig. 3 Dynamics of ammonia, nitrite and nitrate concentrations and pH for microcosm 12-0, 12-1 and 12-50 during the cycle 1 (a, b and c) and cycle 8 (d, e, f), respectively. Data plotted are means and standard deviations of influent ammonia and effluent nitrite and nitrate concentrations of the duplicate microcosms. Some error bars are smaller than the symbol size. | ||
3.2. In situ activity of AOA and AOB in original sludge
DNA-SIP was applied to determine the in situ activity of AOMs in the full-scale WWTP. Ultracentrifugation of the total DNA extracted from each microcosm incubation was performed to separate the 12C-control DNA and 13C-labeled DNA, which resulted in 15 equal fractions covering buoyant densities of 1.69 to 1.75 g mL−1. The buoyant densities decreased from fraction 1 to 15. All the fractions were subjected to amplifying AOA and AOB amoA genes. For the control microcosm (12-0), both AOA and AOB amoA genes could be amplified from the “light” DNA fractions. For the labeled microcosm (13-0), only AOB amoA gene could be amplified from the “heavy” and “light” DNA fractions. The results provided evidence for the incorporation of 13C into genomic DNA of AOB during the active nitrification in microcosm 13-0, and suggested that AOB were the active AOMs in original sludge.The abundances of AOB and AOA amoA genes in each fraction of control (12-0) and labeled microcosms (13-0) were further quantified by qPCR. However, the quantification of AOA amoA gene failed. Fig. 4a shows the distribution of the relative abundance of AOB amoA gene in CsCl gradient for 12-0 and 13-0 microcosms. The plot values are the proportion of AOB amoA gene copy numbers in each fraction to the maximum abundance across the gradient. For the control microcosm (12-0), the highest abundance of AOB amoA gene was observed in the “light” DNA fractions (∼1.72 g mL−1) (Fig. 4a). For the labeled microcosm (13-0), AOB amoA gene peaked in both “light” and “heavy” fractions (Fig. 4a). These results were in agreement with the PCR results, and further provided the compelling evidence for successful labeling of AOB in original sludge. However, the peak of AOB amoA gene in “light” fraction was higher than that in “heavy” fraction, suggesting the insufficient labeling of AOB in the labeled microcosm. In this study, we aimed to investigate the active AOMs directly involved in the in situ nitrification of the full-scale WWTP, therefore short-term microcosms (only eight cycles) were designed to avoid or minimize carbon cross-feeding. A previous study has suggested that the cross-feeding of 13C occurred after long-term incubation.52
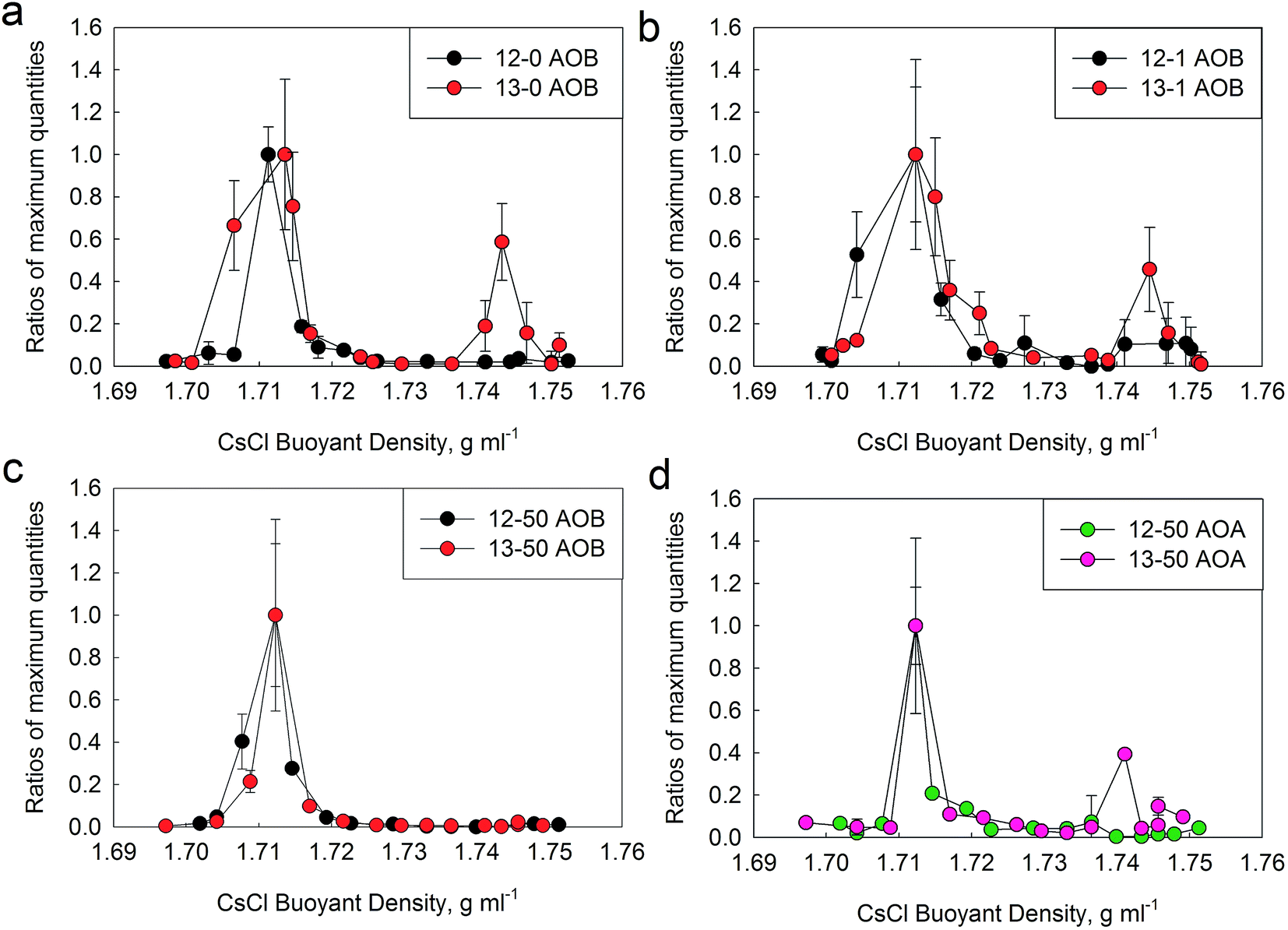 | ||
| Fig. 4 Distribution of the relative abundance of AOB (a, b and c) and AOA (d) amoA genes retrieved from CsCl gradients for the 13C-Na2CO3 and 12C-Na2CO3 treatments during DNA-SIP microcosms under different TiO2 NPs concentrations. Fifteen fractions of genomic DNA extracted from 5.1 mL centrifuge tube covered a buoyant densities of 1.69 to 1.75 g mL−1. The abundances of AOB and AOA amoA genes were determined by qPCR in triplicate. The plot values are the proportion of AOB or AOA amoA gene copy numbers in each fraction to the average maximum abundance across the gradient. The stand deviation of the triplicate qPCR results of each fraction is shown, with some error bars smaller than the symbol size. | ||
In this study, AOB rather than AOA dominated the active ammonia oxidation in original sludge. This result was in agreement with the previous studies regarding active AOMs in neutral, alkaline and N-rich agriculture soils,22,23,53–55 estuarine sediments56 and the laboratory cultures.57 However, the reverse result, active ammonia oxidation was carried out by AOA rather than AOB, was obtained in a grassland soil.58 The most significant difference between the grassland soil and the two neutral agricultural soils is the ammonia concentration in the microcosms.58 The ammonia in the grassland soil was derived from mineralization of soil organic matter, resulting in the low ammonia concentrations (<3.5 μg NH4+-N per g soil), whereas 100 μg NH4+-N per g soil was added to the agricultural soil microcosms once a week. The results suggested that ammonia concentration is a key factor for the different niches of active AOMs. Moreover, AOA were also the active AOMs in acid soils.24–26,54,59 Low pH can result in the exponential ionization of ammonia to ammonium,60 and therefore decreasing the concentration of ammonia substrate. Compared with AOB, the half-saturation constant of AOA is extremely low and the affinity to ammonia to AOA is high.61 Therefore, in lower pH environment, the low ammonia was the critical factor for active AOA. In this study, the average ammonia concentration and pH of synthetic wastewater were (20.39 ± 2.82) mg L−1 and 7.8 ± 0.5, respectively. Both ammonia concentration and pH were in favor of AOB, which were one possible reason for that active AOB dominated the active ammonia oxidation in original sludge.
Moreover, AOB (3.96 × 108 copies per g sludge) outnumber AOA (1.73 × 105 copies per g sludge) in original sludge,11 which was another possible reason for that AOB were responsible for the active ammonia oxidation in the full-scale WWTP. However, in an agriculture soil, where AOA outnumbered AOB in different depth soil samples, the 13C incorporation into DNA of AOB is observed,23 suggesting AOB are the active AOMs. Therefore, the abundance of AOMs could not be directly linked with the active AOMs.
3.3. Communities of active AOB in original sludge
The PCR results suggested that only AOB amoA gene was detected in the “heavy” DNA fractions of 13-0 microcosm. Therefore, the 13C-labeled DNA of microcosm 13-0 was applied for subsequent analysis to investigate the communities of active AOB in original sludge collected from the full-scale WWTP. In total, 34 AOB amoA gene sequences were obtained, which were grouped into five OTUs. As shown in Fig. 5, all the OTUs belonged to Nitrosomonas genera, including two clusters: N. oligotropha cluster (64.71%) and N. europaea cluster (35.29%). The results suggested that N. oligotropha cluster and N. europaea cluster were the dominant active AOB in the full-scale WWTP investigated. The majority of active AOB in original sludge belonged to the N. oligotropha cluster, which was in agreement with the active AOB in biofilm enrichments62 and coastal sediment.21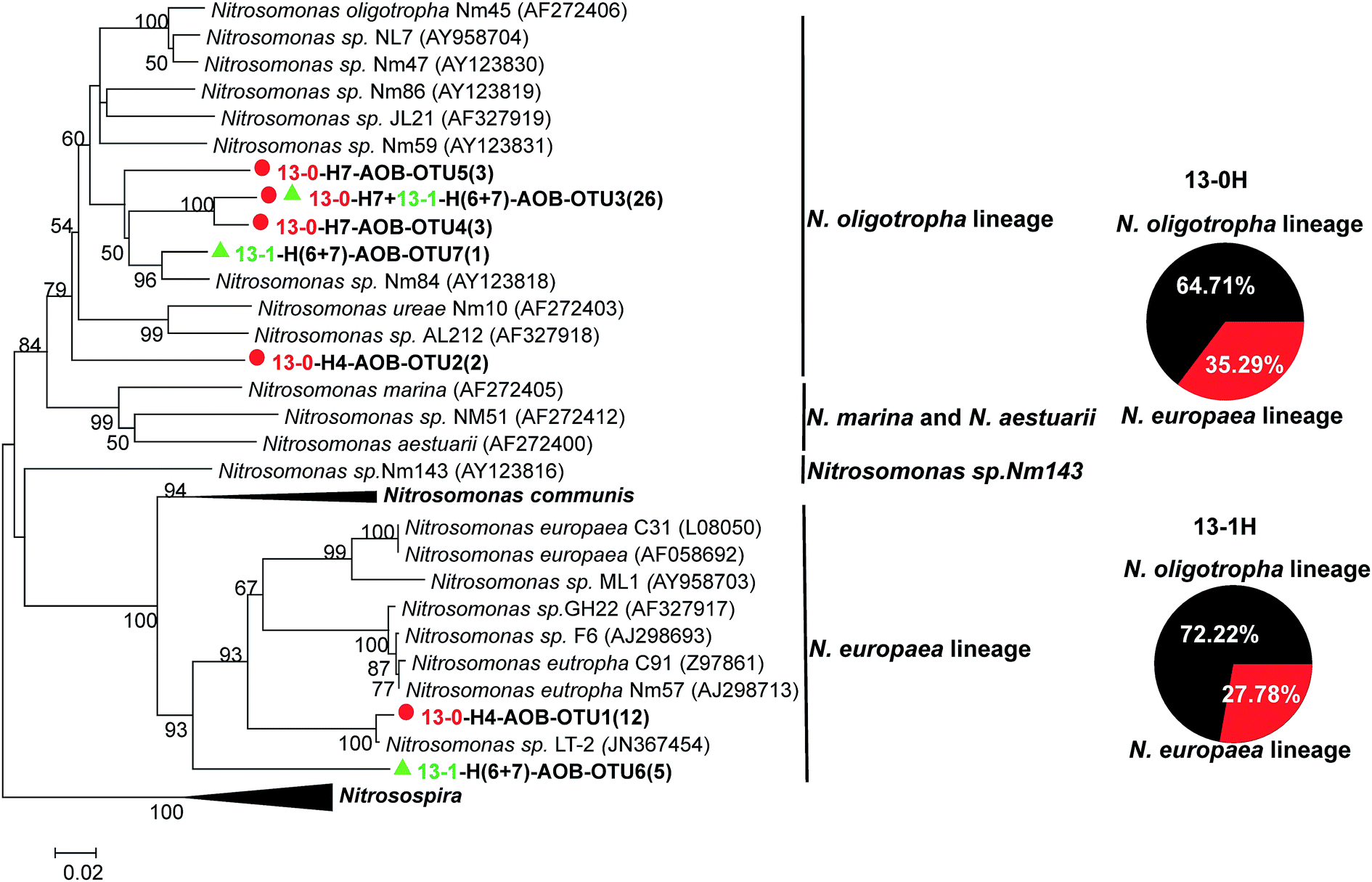 | ||
| Fig. 5 NJ phylogenetic tree based on AOB amoA gene sequences in 13C-labeled DNA from the two labeled microcosms (13-0 and 13-1) after an incubation period of eight cycles. The designation “H” indicates the “heavy” fractions of the labeled microcosm. The designation “13-0-H7-AOB-OTU5(3)” indicates that OTU5 contains three AOB amoA sequences in the fraction seven of the 13C-labeled DNA from 13-0 microcosm incubation. Representative sequences obtained from 13-0 and 13-1 microcosms are indicated by red circle and green triangle, respectively. | ||
In original sludge, N. oligotropha cluster (70.01%) and N. europaea cluster (10.06%) were the dominant AOB,11 which were also the dominant active AOMs. The results indicated that the short-term laboratory microcosms of original sludge well reflected the in situ conditions of the sludge in the full-scale WWTP. Also, the results suggested that 13C-DNA-SIP is an effective method to investigate the in situ activity of AOMs. Moreover, in high ammonia concentration environment, other active AOB belonging to Nitrosomonas genera, e.g., Nitrosomonas sp. Nm143 cluster and Nitrosomonas communis, were observed in estuarine sediments and aquatic environment, respectively.56,63 The results further suggested that ammonia concentration plays an important role in different niches of active AOB genera. However, the active AOB in agriculture soils were Nitrosospira,23,53 which was not detected in the 13C-DNA from original sludge in this study.
3.4. Short-term effects of TiO2 NPs on the abundance and community of active AOMs
DNA-SIP was also applied to assess the short-term effects of different TiO2 NPs concentrations (1 and 50 mg L−1) on the abundance and composition of active AOMs. For the control microcosms (12-1 and 12-50), both AOA and AOB amoA genes could be amplified from the “light” DNA fractions. For the labeled microcosm (13-1), only AOB amoA gene could be amplified from the “light” and “heavy” DNA fractions, providing evidence for incorporation of 13C into the genomic DNA of AOB under 1 mg L−1 of TiO2 NPs. For another labeled microcosm (13-50), only AOA amoA gene could be amplified from the “light” and “heavy” DNA fractions, suggesting the successful labeling of AOA under 50 mg L−1 of TiO2 NPs. However, the bands of AOA in the “heavy” DNA fractions from 13-50 were weaker than those of AOB in the “heavy” DNA fractions from 13-0 and 13-1. These results suggested that AOB and AOA were the active AOMs under 1 and 50 mg L−1 TiO2 NPs, respectively.For these four microcosms, the proportions of AOB and AOA amoA gene copy numbers in each fraction to the maximum abundance across the entire gradient are depicted in Fig. 4. As shown in Fig. 4b and c, similar as the original sludge, the highest abundance of AOB amoA gene was observed in the “light” DNA fractions (∼1.72 g mL−1) under 1 and 50 mg L−1 TiO2 NPs. However, compared with the original sludge, the peak value of AOB in the “heavy” fractions decreased slightly under 1 mg L−1 TiO2 NPs, and which was almost disappeared under 50 mg L−1 TiO2 NPs. The results indicated that the presence of environmentally relevant concentration of TiO2 NPs had a slight inhibition on the abundance of active AOB. However, the higher concentration of TiO2 NPs could completely inhibit the activity of AOB. The abundance of AOA was only successfully quantified in the microcosm 12-50 and 13-50, which was in accordance with the results of PCR mentioned above. As shown in Fig. 4d, for the control microcosm 12-50, AOA amoA gene only peaked in the “light” fractions (∼1.72 g mL−1), and for the labeled microcosm (13-50), AOA amoA gene peaked in both “light” and “heavy” fractions. The results indicated the successful labeling of AOA in activated sludge under 50 mg L−1 TiO2 NPs, suggesting that the active AOA could endure higher TiO2 NPs concentration than AOB.
For AOB amoA gene, no product was obtained from PCR amplification owing to the extremely low abundance in the 13C-DNA from microcosm 13-50. The clone library of 13C-labeled DNA in microcosm 13-1 was successfully constructed. In total, 18 AOB amoA gene sequences were obtained, which were grouped into three OTUs. The rarefaction curve arrived to the plateau phase. All the OTUs belonged to N. oligotropha cluster and N. europaea cluster. Except the presence of 1 mg L−1 TiO2 NPs, 13-0 and 13-1 microcosms were incubated under the same conditions. However, compared with original sludge, distributions of the two clusters in the 13C-labeled DNA from microcosm 13-1 changed (Fig. 5). The relative abundance of N. oligotropha cluster increased from 64.71% to 72.22% in the presence of 1 mg L−1 TiO2 NPs, and which of N. europaea cluster decreased from 35.29% to 27.78%. Moreover, in this study, short-term incubation was designed to avoid 13C cross-feeding. Therefore, shifts of the distributions of active AOB under 0 and 1 mg L−1 TiO2 NPs suggested that environmentally relevant concentration of TiO2 NPs had a slight impact on the compositions of active AOMs.
For 13C-labeled DNA from microcosm 13-50, 16 AOA amoA gene sequences were obtained, which were classified into 11 OTUs. The representative AOA amoA gene sequences were selected for phylogenetic analysis. Similar as the original sludge, all the OTUs obtained were affiliated to Nitrososphaera cluster, including general Nitrososphaera cluster (68.75%), Nitrososphaera subcluster 9 (18.75%), Nitrososphaera subcluster 8.1 (6.25%) and Nitrososphaera subcluster 4.1 (6.25%) (Fig. 6), suggesting AOA within these Nitrososphaera clusters were the active AOMs under higher concentration of TiO2 NPs. The results also suggested that the microcosm under 50 mg L−1 TiO2 NPs could well reflect the short-term impact of TiO2 NPs on AOA in the full-scale WWTP. Furthermore, the result suggested that some or all of the Nitrososphaera cluster had a strong tolerance to higher concentration of TiO2 NPs. AOB within N. oligotropha cluster and N. europaea cluster were the active AOMs under 0 and 1 mg L−1 TiO2 NPs, however their relative abundances were different. These results provided direct evidence that the presence of TiO2 NPs caused shifts in the compositions of active AOMs. In summary, higher concentration of TiO2 NPs showed a more obvious impact on the abundance and communities of active AOMs than the environmentally relevant concentration.
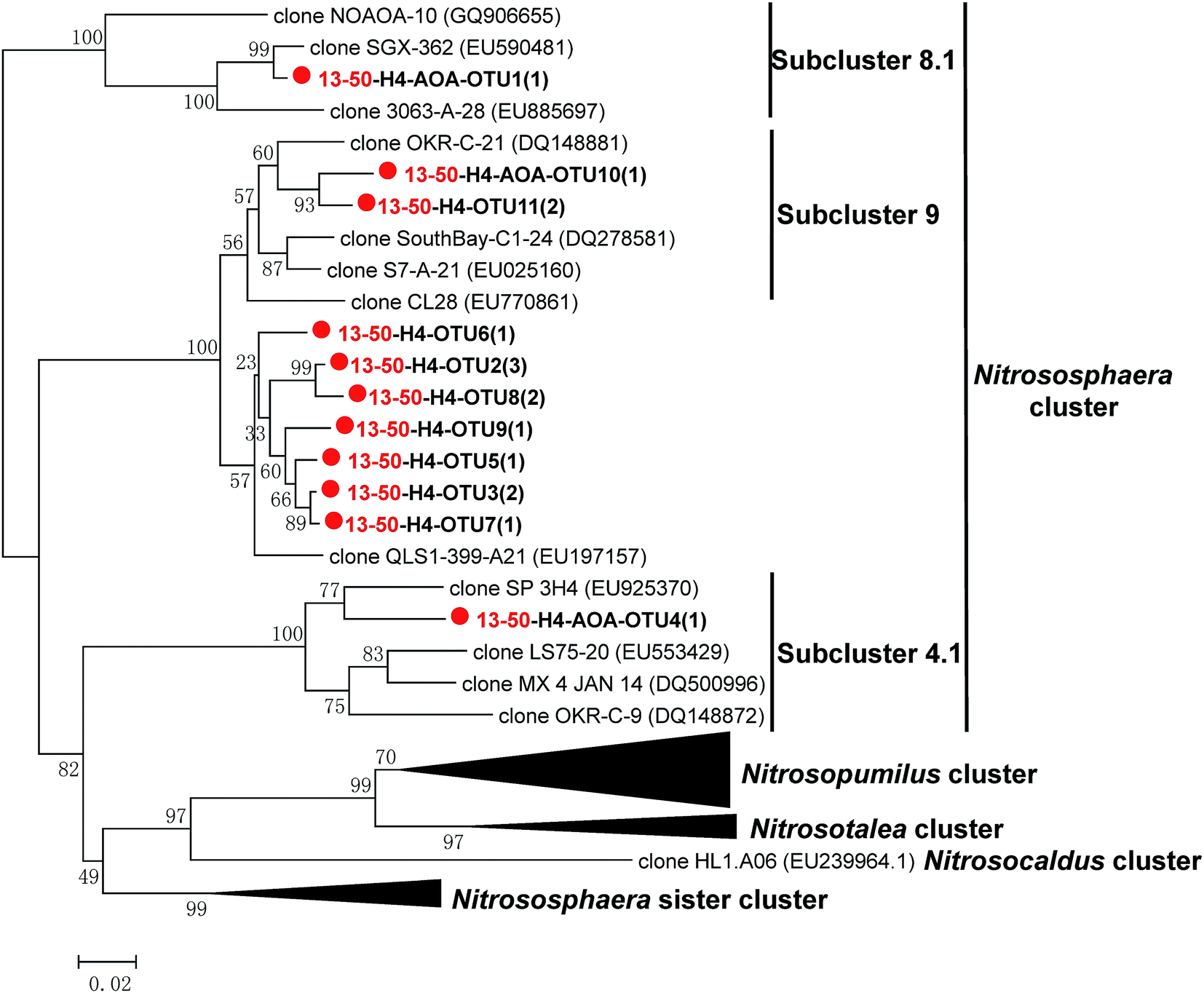 | ||
| Fig. 6 NJ phylogenetic tree based on AOA amoA gene sequences in 13C-labeled DNA from the labeled microcosm (13-50). The designation “H” indicates the “heavy” fractions of the labeled microcosm. The designation “13-50-H4-AOA-OTU1(1)” indicates that OTU1 contains one AOA amoA sequence in the fraction four of the 13C-labeled DNA from 13-50 microcosm. Representative sequences obtained from the microcosm are indicated by color symbol (red circle). | ||
Previous studies suggested that the abundance of AOB was inhibited by TiO2 NPs at predicted environmentally relevant concentration and higher concentration,38,64 which were further confirmed in this study using DNA-SIP. The abundance of active AOB decreased at environmentally relevant concentration of TiO2 NPs (1 mg L−1) and even completely inhibited at higher concentration of TiO2 NPs (50 mg L−1). The 50 mg L−1 TiO2 NPs are confirmed to inhibit the gene expressions and catalytic activities of ammonia monooxygenase,38,39 which might be the reason for the significant decrease of AOB abundance. Moreover, the distributions of active AOB genera changed at the environmentally relevant concentration of TiO2 NPs. At the presence of 1 mg L−1 TiO2 NPs, the relative abundance of N. oligotropha cluster increased, suggesting that that the environmentally relevant concentration of TiO2 NPs might be in favor of the N. oligotropha cluster. Whereas, the relative abundance of N. europaea cluster decreased, indicating the environmentally relevant concentration of TiO2 NPs showed a negative impact on N. europaea cluster. In previous studies, TiO2 NPs are reported to be toxic to the pure culture of N. europaea cluster, a model AOB, which might explain the decrease of N. europaea cluster.65,66
In this study, some interesting and important issues were found: (1) the nitrification rate was similar at 0, 1 and 50 mg L−1 TiO2 NPs; (2) AOB were the active AOMs in original sludge and the sludge incubated under 1 mg L−1 TiO2 NPs, and AOA were the active AOMs under 50 mg L−1 TiO2 NPs. However, the previous study suggested that Km of AOA is much lower than AOB,61 and how to explain the similar nitrification rate of AOA under 50 mg L−1 TiO2 NPs as AOB under 0 and 1 mg L−1 TiO2 NPs? The impact of higher concentration of TiO2 NPs on AOA is unknown. One possible reason was that high concentration of TiO2 NPs acted as a catalyst, which enhanced the ammonia degradation speed of active AOA. A previous study suggested that the addition of TiO2 NPs accelerates the decomposition speed of poly(L-lactide) by microorganism.67 The previous studies mainly focused on the impact of TiO2 NPs on bacterial communities (including AOB),40,68,69 little is known about the impact of TiO2 NPs on AOA. Therefore, more studies are needed to investigate the impact of higher concentration of TiO2 NPs on AOA.
3.5. Autotrophic growth of active AOMs
In this study, Na213CO3 was used as the labeled substrate in DNA-SIP microcosms. AOB within N. oligotropha cluster and N. europaea cluster, and AOA within general Nitrososphaera cluster were the active AOMs in different microcosms. Therefore, our study provided direct evidence for the autotrophic growth of these active AOMs. Previous studies using 13CO2 as the labeled substrate in agriculture soils22,53 also demonstrated the autotrophic growth of AOB and AOA, which was in agreement with this study. The autotrophic growth of AOB was further confirmed in other agriculture soil23 and biofilm enrichments.62 There are also evidences of autotrophic growth of AOA in some acidic soils24,25 and an agriculture soil in Europe.58 Despite the autotrophic growth of AOB and AOA by using inorganic carbon, their pathways for carbon fixation were completely different. AOB use the Calvin–Bassham–Benson cycle for carbon fixation, whereas AOA seems to use 3-hydroxypropionate/4-hydroxybutyrate (3HP/4HB) and oxidative tricarboxylic acid (TCA) cycle.704. Conclusions
The in situ activity of AOA and AOB in a full-scale WWTP and their short-term responses to different concentrations of TiO2 NPs were investigated using 13C-DNA-SIP. Nitrification activity in all the microcosms was observed, which was slightly or not inhibited by TiO2 NPs. This study provided the compelling evidence for Na213CO3 assimilation of AOB rather than AOA in original sludge, suggesting a strong contribution of AOB to nitrification in the full-scale WWTP studied. The active AOB belonged to the N. oligotropha cluster and N. europaea cluster, which was in accordance with the compositions of AOB in original sludge. Interestingly, AOA within Nitrososphaera cluster were the active AOMs under higher TiO2 NPs concentration (50 mg L−1). TiO2 NPs showed a potential short-term impact on the abundance and composition of active AOMs. Furthermore, this study provided direct evidence for the autotrophic growth of N. oligotropha cluster, N. europaea cluster and Nitrososphaera cluster in the full-scale WWTP. Taken together, this study revealed the critical role of AOA to nitrification under higher TiO2 NPs concentration.Acknowledgements
We would like to thank the NSFC (51078007, 51378027 and 51578015), BJNSF (8162010) and Beijing Talent Foundation of BJUT (2013-JH-L06) for the financial supports of this study.References
- M. Konneke, A. E. Bernhard, J. R. de la Torre, C. B. Walker, J. B. Waterbury and D. A. Stahl, Nature, 2005, 437, 543–546 CrossRef PubMed.
- C. A. Francis, K. J. Roberts, J. M. Beman, A. E. Santoro and B. B. Oakley, Proc. Natl. Acad. Sci. U. S. A., 2005, 102, 14683–14688 CrossRef CAS PubMed.
- S. Leininger, T. Urich, M. Schloter, L. Schwark, J. Qi, G. W. Nicol, J. I. Prosser, S. C. Schuster and C. Schleper, Nature, 2006, 442, 806–809 CrossRef CAS PubMed.
- J. M. Beman and C. A. Francis, Appl. Environ. Microbiol., 2006, 72, 7767–7777 CrossRef CAS PubMed.
- D. R. Rogers and K. L. Casciotti, Appl. Environ. Microbiol., 2010, 76, 7938–7948 CrossRef CAS PubMed.
- I. Kasuga, H. Nakagaki, F. Kurisu and H. Furumai, Water Sci. Technol., 2010, 61, 3070–3077 CrossRef CAS PubMed.
- H. D. Park, G. F. Wells, H. Bae, C. S. Criddle and C. A. Francis, Appl. Environ. Microbiol., 2006, 72, 5643–5647 CrossRef CAS PubMed.
- T. Zhang, T. Jin, Q. Yan, M. Shao, G. Wells, C. Criddle and H. H. P. Fang, J. Appl. Microbiol., 2009, 107, 970–977 CrossRef CAS PubMed.
- J. F. Gao, X. Luo, G. X. Wu, T. Li and Y. Z. Peng, Appl. Microbiol. Biotechnol., 2014, 98, 3339–3354 CrossRef CAS PubMed.
- J. You, A. Das, E. M. Dolan and Z. Hu, Water Res., 2009, 43, 1801–1809 CrossRef CAS PubMed.
- J. F. Gao, X. Luo, G. X. Wu, T. Li and Y. Z. Peng, Bioresour. Technol., 2013, 138, 285–296 CrossRef CAS PubMed.
- G. F. Wells, H. Park, C. Yeung, B. Eggleston, C. A. Francis and C. S. Criddle, Environ. Microbiol., 2009, 11, 2310–2328 CrossRef CAS PubMed.
- T. Limpiyakorn, M. Fuerhacker, R. Haberl, T. Chodanon, P. Srithep and P. Sonthiphand, Appl. Microbiol. Biotechnol., 2013, 97, 1425–1439 CrossRef CAS PubMed.
- L. A. Sauder, F. Peterse, S. Schouten and J. D. Neufeld, Environ. Microbiol., 2012, 14, 2589–2600 CrossRef CAS PubMed.
- T. Zhang, L. Ye, A. H. Y. Tong, M. Shao and S. Lok, Appl. Microbiol. Biotechnol., 2011, 91, 1215–1225 CrossRef CAS PubMed.
- T. Jin, T. Zhang and Q. Yan, Appl. Microbiol. Biotechnol., 2010, 87, 1167–1176 CrossRef CAS PubMed.
- P. Kayee, P. Sonthiphand, C. Rongsayamanont and T. Limpiyakorn, Microb. Ecol., 2016, 72, 262 CrossRef PubMed.
- S. Radajewski, P. Ineson, N. R. Parekh and J. C. Murrell, Nature, 2000, 403, 646–649 CrossRef CAS PubMed.
- S. Coyotzi, J. Pratscher, J. C. Murrell and J. D. Neufeld, Curr. Opin. Biotechnol., 2016, 41, 1–8 CrossRef CAS PubMed.
- J. D. Neufeld, J. Vohra, M. G. Dumont, T. Lueders, M. Manefield, M. W. Friedrich and J. C. Murrell, Nat. Protoc., 2007, 2, 860–866 CrossRef CAS PubMed.
- Y. Zhang, L. Chen, T. Dai, R. Sun and D. Wen, Appl. Microbiol. Biotechnol., 2015, 99, 6481–6491 CrossRef CAS PubMed.
- J. Pratscher, M. G. Dumont and R. Conrad, Proc. Natl. Acad. Sci. U. S. A., 2011, 108, 4170–4175 CrossRef CAS PubMed.
- Z. J. Jia and R. Conrad, Environ. Microbiol., 2009, 11, 1658–1671 CrossRef CAS PubMed.
- L. M. Zhang, H. W. Hu, J. P. Shen and J. Z. He, ISME J., 2012, 6, 1032–1045 CrossRef CAS PubMed.
- L. Lu and Z. J. Jia, Environ. Microbiol., 2013, 15, 1795–1809 CrossRef CAS PubMed.
- J. Z. He, H. W. Hu and L. M. Zhang, Soil Biol. Biochem., 2012, 55, 146–154 CrossRef CAS.
- J. Zhao, B. Wang and Z. Jia, Appl. Environ. Microbiol., 2015, 81, 3218–3227 CrossRef CAS PubMed.
- X. J. Jiang, X. Y. Hou, X. Zhou, X. P. Xin, A. Wright and Z. J. Jia, Soil Biol. Biochem., 2015, 81, 9–16 CrossRef CAS.
- H. W. Hu, C. A. Macdonald, P. Trivedi, B. Holmes, L. Bodrossy, J. Z. He and B. K. Singh, Environ. Microbiol., 2015, 17, 444–461 CrossRef CAS PubMed.
- J. Niu, I. Kasuga, F. Kurisu, H. Furumai and T. Shigeeda, Water Res., 2013, 47, 7053–7065 CrossRef CAS PubMed.
- F. Gottschalk, T. Sonderer, R. W. Scholz and B. Nowack, Environ. Sci. Technol., 2009, 43, 9216–9222 CrossRef CAS PubMed.
- A. B. Djurisic, Y. H. Leung, A. M. C. Ng, X. Y. Xu, P. K. H. Lee, N. Degger and R. S. S. Wu, Small, 2015, 11, 26–44 CrossRef CAS PubMed.
- K. R. Raghupathi, R. T. Koodali and A. C. Manna, Langmuir, 2011, 27, 4020–4028 CrossRef CAS PubMed.
- Q. Li, S. Mahendra, D. Y. Lyon, L. Brunet, M. V. Liga, D. Li and P. J. Alvarez, Water Res., 2008, 42, 4591–4602 CrossRef CAS PubMed.
- P. Westerhoff, G. Song, K. Hristovski and M. A. Kiser, J. Environ. Monit., 2011, 13, 1195–1203 RSC.
- M. A. Kiser, H. Ryu, H. Jang, K. Hristovski and P. Westerhoff, Water Res., 2010, 44, 4105–4114 CrossRef CAS PubMed.
- B. Thio, D. Zhou and A. A. Keller, J. Hazard. Mater., 2011, 189, 556–563 CrossRef CAS PubMed.
- X. Zheng, Y. Chen and R. Wu, Environ. Sci. Technol., 2011, 45, 7284–7290 CrossRef CAS PubMed.
- X. Zheng, H. Huang, Y. Su, Y. Wei and Y. Chen, Water Sci. Technol., 2015, 72, 99–105 CrossRef CAS PubMed.
- D. Li, F. Cui, Z. Zhao, D. Liu, Y. Xu, H. Li and X. Yang, Biodegradation, 2014, 25, 167–177 CrossRef CAS PubMed.
- B. Li, W. L. Huang, C. Zhang, S. S. Feng, Z. Zhang, Z. F. Lei and N. Sugiura, Bioresour. Technol., 2015, 187, 214–220 CrossRef CAS PubMed.
- J. Zhang, Q. Dong, Y. Liu, X. Zhou and H. Shi, Chemosphere, 2016, 144, 1837–1844 CrossRef CAS PubMed.
- A. A. Keller, H. Wang, D. Zhou, H. S. Lenihan, G. Cherr, B. J. Cardinale, R. Miller and Z. X. Ji, Environ. Sci. Technol., 2010, 44, 1962–1967 CrossRef CAS PubMed.
- APHA, Standard methods for the examination of water and wastewater, Washington DC, USA, 20th edn, 1998 Search PubMed.
- J. F. Gao, X. Y. Fan, G. X. Wu, T. Li and K. L. Pan, Desalin. Water Treat., 2015, 1–13 Search PubMed.
- F. Verhagen and H. J. Laanbroek, Appl. Environ. Microbiol., 1991, 57, 3255–3263 CAS.
- U. Purkhold, A. Pommerening-Roser, S. Juretschko, M. C. Schmid, H. P. Koops and M. Wagner, Appl. Environ. Microbiol., 2000, 66, 5368–5382 CrossRef CAS PubMed.
- P. D. Schloss, S. L. Westcott, T. Ryabin, J. R. Hall, M. Hartmann, E. B. Hollister, R. A. Lesniewski, B. B. Oakley, D. H. Parks, C. J. Robinson, J. W. Sahl, B. Stres, G. G. Thallinger, D. J. Van Horn and C. F. Weber, Appl. Environ. Microbiol., 2009, 75, 7537–7541 CrossRef CAS PubMed.
- N. Goyal, D. Rastogi, M. Jassal and A. K. Agrawal, J. Dispersion Sci. Technol., 2013, 34, 611–622 CrossRef CAS.
- W. Jiang, H. Mashayekhi and B. S. Xing, Environ. Pollut., 2009, 157, 1619–1625 CrossRef CAS PubMed.
- Y. Ma, J. W. Metch, E. P. Vejerano, I. J. Miller, E. C. Leon, L. C. Marr, P. J. Vikesland and A. Pruden, Water Res., 2015, 68, 87–97 CrossRef CAS PubMed.
- E. Gallagher, L. McGuinness, C. Phelps, L. Y. Young and L. J. Kerkhof, Appl. Environ. Microbiol., 2005, 71, 5192–5196 CrossRef CAS PubMed.
- W. Xia, Z. Cai, Z. Jia, C. Zhang, X. Zeng, Y. Feng, W. Jiahua, X. Lin, J. Zhu, Z. Xiong and J. Xu, ISME J., 2011, 5, 1226–1236 CrossRef CAS PubMed.
- J. P. Shen, L. M. Zhang, H. J. Di and J. Z. He, Front. Microbiol., 2012, 3, 296 CAS.
- S. Ruyters, G. W. Nicol, J. I. Prosser, B. Lievens and E. Smolders, Soil Biol. Biochem., 2013, 58, 244–247 CrossRef CAS.
- T. E. Freitag, L. Chang and J. I. Prosser, Environ. Microbiol., 2006, 8, 684–696 CrossRef CAS PubMed.
- M. Tourna, T. E. Freitag, G. W. Nicol and J. I. Prosser, Environ. Microbiol., 2008, 10, 1357–1364 CrossRef CAS PubMed.
- L. M. Zhang, P. R. Offre, J. Z. He, D. T. Verhamme, G. W. Nicol and J. I. Prosser, Proc. Natl. Acad. Sci. U. S. A., 2010, 107, 17240–17245 CrossRef CAS PubMed.
- B. Z. Wang, Y. Zheng, R. Huang, X. Zhou, D. M. Wang, Y. Q. He and Z. J. Jia, Appl. Environ. Microbiol., 2014, 80, 1684–1691 CrossRef PubMed.
- W. De Boer and G. A. Kowalchuk, Soil Biol. Biochem., 2001, 33, 853–866 CrossRef CAS.
- W. Martens-Habbena, P. M. Berube, H. Urakawa, J. R. de la Torre and D. A. Stahl, Nature, 2009, 461, 976–979 CrossRef CAS PubMed.
- S. Avrahami, Z. Jia, J. D. Neufeld, J. C. Murrell, R. Conrad and K. Kusel, Appl. Environ. Microbiol., 2011, 77, 7329–7338 CrossRef CAS PubMed.
- Y. Chen, L. Wu, R. Boden, A. Hillebrand, D. Kumaresan, H. Moussard, M. Baciu, Y. Lu and M. J. Colin, ISME J., 2009, 3, 1093–1104 CrossRef CAS PubMed.
- Z. Luo, Z. Qiu, Z. Chen, G. Du Laing, A. Liu and C. Yan, Environ. Sci. Pollut. Res., 2015, 22, 2891–2899 CrossRef CAS PubMed.
- A. Ostermeyer, C. K. Mumuper, L. Semprini and T. Radniecki, Environ. Sci. Technol., 2013, 47, 14403–14410 CrossRef CAS PubMed.
- X. Fang, R. Yu, B. Li, P. Somasundaran and K. Chandran, J. Colloid Interface Sci., 2010, 348, 329–334 CrossRef CAS PubMed.
- J. Lee, J. Lee, B. H. Park, A. Busnaina and H. Y. Lee, J. Nanosci. Nanotechnol., 2013, 13, 6983–6987 CrossRef PubMed.
- Z. Y. Liu, S. L. Yu, H. Park, Q. B. Yuan, G. C. Liu and Q. Li, Environ. Sci. Pollut. Res. Int., 2016, 1–10 CrossRef PubMed.
- Y. Ge, J. P. Schimel and P. A. Holden, Environ. Sci. Technol., 2011, 45, 1659–1664 CrossRef CAS PubMed.
- C. B. Walker, J. R. de la Torre, M. G. Klotz, H. Urakawa, N. Pinel, D. J. Arp, C. Brochier-Armanet, P. S. Chain, P. P. Chan, A. Gollabgir, J. Hemp, M. Hugler, E. A. Karr, M. Konneke, M. Shin, T. J. Lawton, T. Lowe, W. Martens-Habbena, L. A. Sayavedra-Soto, D. Lang, S. M. Sievert, A. C. Rosenzweig, G. Manning and D. A. Stahl, Proc. Natl. Acad. Sci. U. S. A., 2010, 107, 8818–8823 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2016 |