
Role of counter-ion and helper lipid content in the design and properties of nanocarrier systems: a biophysical study in 2D and 3D lipid assemblies†
Ana C. N. Oliveiraab,
Sara S. Nogueirabc,
Odete Gonçalvesab,
M. F. Cerqueirab,
P. Alpuimbd,
Júlia Tovarb,
Carlos Rodriguez-Abreud,
Gerald Brezesinskic,
Andreia C. Gomesa,
Marlene Lúcio *b and
M. E. C. D. Real Oliveirab
*b and
M. E. C. D. Real Oliveirab
aCBMA (Centre of Molecular and Environmental Biology), Department of Biology, University of Minho, Campus of Gualtar, 4710-057 Braga, Portugal
bCFUM (Centre of Physics), Department of Physics, University of Minho, Campus of Gualtar, 4710-057 Braga, Portugal. E-mail: mlucio@fisica.uminho.pt
cMax Planck Institute of Colloids and Interfaces, Science Park Potsdam-Golm, 14476 Potsdam, Germany
dINL (International Iberian Nanotechnology Laboratory), Av. Mestre José Veiga, 4715-330 Braga, Portugal
First published on 5th May 2016
Abstract
There is a direct correlation between the physicochemical properties of nanocarrier systems and their biological performance, including stability under physiological conditions, cellular internalization and transfection efficiency. Therefore, understanding the biophysical aspects that affect self-assembled nanocarriers is determinant for a rational design of efficient formulations. In this study, a comprehensive evaluation of the effects of each component on the molecular organization of aggregates formed by the cationic lipids dioctadecyldimethylammonium bromide and chloride (DODAB and DODAC) and the neutral lipid monoolein (MO) was made. Specifically, the effects of the helper lipid content (MO) and the role of the counter-ion of the cationic lipids were evaluated in 2D and 3D assemblies by Langmuir surface pressure–molecular area (π–A) isotherms, Brewster Angle Microscopy (BAM), infrared reflection absorption spectroscopy (IRRAS), confocal Raman microscopy, and Small Angle X-ray Scattering (SAXS). The results show that MO has a different distribution on the DODAC and DODAB bilayers, and a fluidizing effect dependent on the MO content. For low MO molar ratios, the fluidizing effect was more pronounced in DODAC![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :MO mixtures, indicating a more homogeneous distribution of MO in DODAC than in DODAB bilayers. For high MO molar ratios, packing of membranes was similar for both cationic lipids, and the effect of the counter-ion is attenuated. The distribution of MO in the two cationic systems is closely related with the efficiency of the counter-ions in the screening of the charged group.
:MO mixtures, indicating a more homogeneous distribution of MO in DODAC than in DODAB bilayers. For high MO molar ratios, packing of membranes was similar for both cationic lipids, and the effect of the counter-ion is attenuated. The distribution of MO in the two cationic systems is closely related with the efficiency of the counter-ions in the screening of the charged group.
Introduction
Liposomes have been extensively explored as drug and genetic material delivery systems.3,4 Cationic liposomes, for instance, are largely used for gene delivery purposes because they promote compaction of nucleic acids by electrostatic interactions between the positive charges of cationic lipids and the negative charges of phosphate backbones.5 Neutral (helper) lipids are usually included in cationic liposomal formulations to: help nucleic acids escaping from the endosomes;6–8 aid nucleic acid complexation;9 decrease toxicity associated with cationic lipids;4 or confer more favourable properties in terms of fluidity of the lipid aggregates.10,11 Cholesterol and 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine (DOPE) are the most commonly used helper lipids, but monoolein (MO) also shows promising results in formulations developed for gene therapy.12–20 MO is a neutral lipid that has a low solubility in water but easily swells in contact with an aqueous phase. MO exhibits a lyotropic behaviour forming various lipid phases depending on the temperature and water content, including lamellar (Lα), inverted hexagonal (HII), and bicontinuous cubic (QDII and QGII) phases.21,22 The rich polymorphism of MO, in particular its capacity to form inverted bicontinuous cubic phases in excess water, consisting of two intertwined but not interpenetrating water channels separated by a lipid bilayer surface, offers the possibility to entrap hydrophilic substances in the water channels, lipophilic compounds in the lipid bilayer, and amphiphilic substances.22–24 Besides increasing the entrapment capacity of chemically different payloads, MO inverted non-lamellar phases allow controlling the release of those payloads by a slow diffusion process.22,24,25 Hence, MO can be a very interesting helper lipid in nanostructured lipid carriers for nucleic acid and/or drug delivery purposes.Liposomes composed of MO and lipids from the dioctadecyldimethylammonium family (DODAX, where X stands for Br or Cl) have shown to be efficient nucleic acid delivery systems.14–20,26 Dioctadecyldimethylammonium bromide (DODAB) and chloride (DODAC) assemble into bilayer structures when dispersed in aqueous media above the phase transition temperature.27 Although only differing in the counter-ion, they form vesicles that after conjugation with nucleic acids originate lipoplexes with different characteristics. Indeed, DODAB:MO and DODAC:MO lipoplexes were tested for gene delivery purposes and compared in terms of gene knockdown, toxicity, and stability in human serum.28 Important differences were found between the biological activities of both systems,28 but the explanation for these differences remains elusive, although it seems evident that differences in physical organization of lipid transfection agents may affect their biological outcome.
The main goal of this study is to compare the effects of MO on DODAB and DODAC structural organization, by evaluating the influence of the cationic lipid:neutral lipid molar ratio as well as the effect of the counter-ions on the overall aggregate structure. To our knowledge, this is the first report on the effects of helper lipid content and counter-ion effect on DODAX lipid transfection agents. The structural aspects of DODAX:MO prepared in systems of different dimensionality (two-dimensional (2D) and three-dimensional (3D) membrane models) will also be compared since the molecular organization in different dimensions can provide complementary information on distinct parameters. In 3D spherical membrane mimetic systems (e.g. liposomes), membrane parameters such as packing density, mobility of the molecular components, and topology of the surfaces are simultaneously affected by interactions between the lipid components, and it is very difficult to extract individual contributions of any of these parameters. In this context, 2D membrane models, such as Langmuir monolayers at the soft air/liquid interface, complement the information gathered with 3D lipid models. At the same time, 2D systems provide many possibilities to control, in an effective way, a wide range of experimental variables, such as the phase state of the film-forming molecules, packing density and compressibility of the molecules. Nevertheless, although Langmuir monolayers are excellent model systems for the study of order and packing of lipids in two dimensions29 some properties like miscibility, phase equilibrium, hydration or dynamics might be different for 2D monolayer and 3D multilayer systems.30 This reinforces the importance of using different techniques to confirm and complement results obtained in lipid organizations of different dimensionality. Therefore, we have evaluated DODAX:MO monolayers in 2D assemblies by Langmuir isotherms, Brewster Angle Microscopy (BAM) and infrared reflection absorption spectroscopy (IRRAS in ESI†) and the results were compared with those obtained in 3D assemblies studied by confocal Raman microscopy and Small Angle X-ray Scattering (SAXS).
Experimental
Materials
Dioctadecyldimethylammonium bromide (DODAB) (purity > 98.0%), dioctadecyldimethylammonium chloride (DODAC) (purity > 97.0%) and 1-monooleoyl-rac-glycerol (MO) (purity > 99.0%) were purchased from Sigma-Aldrich (Bornem, Belgium), and used without further purification. Fig. 1 shows the chemical structure of the lipids. | ||
| Fig. 1 Chemical structures of DODAB/DODAC and monoolein (MO) lipids. | ||
Methods
:MO at molar ratios (1:2) and (2:1) were studied as 2D membrane model systems. Surface pressure–molecular area isotherms (π–A) were measured in a PTFE Langmuir trough (Riegler & Kirstein, Potsdam, Germany), equipped with barriers for changing the surface area, and a Wilhelmy microbalance with filter paper plate (accuracy superior to 0.1 mN m−1) for measuring the surface pressure of the monolayer.Monolayers were obtained after spreading lipid solutions (1 mM in chloroform) on the aqueous subphase. After an equilibration time of 10 min, the monolayers were compressed at a rate of 5 Å2 per molecule per min. Before each measurement, the trough was cleaned thoroughly with chloroform and double-deionized water. Cleanliness was confirmed by compressing a water subphase and achieving a zero surface pressure reading. All measurements were performed at 20 °C.
To study the morphology of lipid monolayers, a Brewster Angle Microscope (BAM2plus from NanoFilm Technologie, Göttingen, Germany) was combined with a Langmuir Film Microbalance (NIMA Technology Ltd, United Kingdom) placed on an anti-vibration table. The beam coming from frequency-doubled Nd:YAG laser (532 mm, 50 mW) passed through a polarizer and hits the surface under an incidence angle of 53.1°. The reflection was captured by an analyzer combined with a CCD camera (image size 355 × 470 μm2, resolution approximately 2 μm).
:MO (2:1) or DODAX:MO (1:2) (mol:mol) were formed at 3 mM.Raman scattering experiments of DODAX:MO (2:1) and DODAX:MO (1:2) were performed at room temperature (20 °C) in a back scattering geometry on alpha300 R confocal Raman microscope (WITec), using a 532 nm Nd:YAG laser for excitation. The laser beam was focused on the sample by a ×50 lens (Zeiss); the spectra were collected with a 600 groove per mm grating using 10 acquisitions with a 2 s acquisition time. Lorentzians were fitted to the vibrational bands in the Raman spectra, and the peak positions and corresponding full-width at half-maximum (FWHM) were determined.
Results and discussion
Understanding the key parameters influencing processes such as formation and disassembly of lipoplexes, as well as colloidal stability in physiological conditions are determinant to the rational design of highly efficient nanosystems for gene therapy.Lipoplexes composed by DODAX:MO have been previously reported to induce dissimilar biological responses (e.g. gene silencing efficiency) and to have different colloidal stability in human serum.28 Moreover, DODAX:MO liposomes have shown different siRNA complexation profiles (Fig. S1 – ESI†).
The different biological responses observed motivated the need to perform a detailed study of the influence of the counter-ion and MO content on the biophysical properties of DODAX:MO based formulations. In this regard, the current biophysical study is based on a multi-technique approach to clarify the differences observed between the various DODAX:MO aggregates, by the analysis of lipid structures with different dimensionality (2D by Langmuir isotherms, BAM, IRRAS and 3D by confocal Raman microscopy and SAXS).
Analysis of DODAX![[thin space (1/6-em)]](https://www.rsc.org/images/entities/h3_char_2009.gif) :MO mixtures in 2D assemblies by Langmuir isotherms and BAM
:MO mixtures in 2D assemblies by Langmuir isotherms and BAM
The structures of neat DODAB and DODAC monolayers at the air–water interface have been reported before,31–35 however, to our knowledge, there is no comparative study on the influence of the helper lipid MO on these cationic lipids. Although the effect of MO on the conformation of DODAB molecules has been already investigated,26 that study was not sufficient to clearly explain the different transfection and serum stability behaviours observed for DODAX:MO lipoplexes28 (Fig. S1 – ESI†). Therefore, our previous study26 was extended also to DODAC and to different MO ratios. Accordingly, the surface pressure–molecular area (π–A) isotherms of DODAX:MO monolayers at the air–water interface were obtained for several DODAX:MO compositions, at (20.0 ± 0.2) °C, in order to monitor differences in the interfacial molecular organization of the lipids (Fig. 2A).
 | ||
| Fig. 2 (A) Surface pressure–molecular area (π–A) isotherms (20 °C, aqueous subphase) of DODAX (DODAB or DODAC) and MO monolayers as well as of mixed monolayers at different DODAX:MO molar ratios (0:1, 1:0, 2:1, and 1:2). (B) Compression modulus (Cs−1) calculated from the isotherms presented in (A). Pink stand for DODAC based monolayers, purple colors stand for DODAB based monolayers, and blue color stands for the MO monolayer. The regions of the liquid-condensed phase (LC), of the coexistence between liquid-condensed and liquid-expanded phases (LE + LC), and of the liquid-expanded phase (LE) are marked. (C) BAM images of DODAB monolayer on an aqueous subphase in the different regions LE, LE + LC and LC. | ||
The π–A isotherm obtained for DODAB is in agreement with previously reported isotherms prepared under the same conditions, i.e. aqueous subphase and 20 °C (ref. 32 and 33) and exhibits three distinct regions: a liquid-expanded (LE) phase, at low surface pressures and large areas per molecule; a liquid-expanded/liquid-condensed (LE–LC) phase transition region; and a liquid-condensed (LC) phase at areas per molecule of less than 60 Å2 and high surface pressures, before the collapse near 47 mN m−1 (Fig. 2A – dashed purple line).
The π–A isotherm obtained for DODAC presents a LE phase similar to DODAB, but the transition region cannot be seen (Fig. 2A – pink line), and the areas per molecule at higher pressures are clearly larger compared to DODAB. Hence, in comparison to the LC phase of DODAB, DODAC forms rather a liquid-ordered phase. These results are supported by the IRRAS experiments (Fig. S2 – ESI†). The phase transition from LE to LC in DODAB monolayers is connected with a jump in the wavenumbers of the CH2– stretching vibrations to lower values. E.g., the asymmetric stretching vibration band is observed at 2925 cm−1 in the LE phase and 2920 cm−1 in the LC phase (Fig. S2 – ESI†). The situation is different with DODAC. As in the isotherms, the transition to the LC phase cannot be observed as a jump in wavenumbers (fist-order transition), instead of that, the wavenumbers decrease continuously with increasing pressure. At the highest lateral pressure measured, the wavenumbers of DODAC are at least 1 cm−1 larger than those of DODAB indicating a less dense packing (Fig. S2 – ESI†). Therefore, the change of the counter-ion (from bromide to chloride ions) has a big influence on the type of the transition (second-order instead of first-order) and on the packing density in the monolayer at high-pressure. Chloride ions have a larger hydration shell than bromide ions, and consequently they are not as tightly connected to the charged lipid head group and screen less effectively the cationic charge in the head group region.36–38 This assumption is strongly supported by our previous work using a long-chain sulfate on subphases containing different cations.39 In this work we have shown that the size of counter-ions affects the electrical double layer (EDL). Therefore, the electrostatic term of the free energy of the total system (monolayer + subphase) appears to be dependent on the counter-ion size for highly charged monolayers. In the presence of two different counter-ions, preferential participation of the smaller one in the EDL formation is favourable in terms of free energy. In the present case, since chloride ions are effectively larger, the DODAC monolayers are dominated by stronger electrostatic repulsions between the charged head groups leading to a less dense packing (supported by the higher CH2 wavenumbers at high pressures observed in Fig. S2 – ESI†) compared to DODAB. To illustrate this effect more clearly, DODAB and DODAC have been measured on 10 mM NaBr or NaCl solutions, respectively. In the case of DODAB, the phase transition region (coexistence of LE and LC) is seen as a perfect horizontal plateau at ≈2 mN m−1, whereas DODAC on the 10 mM NaCl solution does not exhibit any plateau but the area decreases continuously with increasing pressure (Fig. S3 – ESI†). In the LC phase, the DODAB chains are strongly tilted (≈34° to the surface normal on a 2 mM NaBr containing subphase) due to the electrostatic repulsions of the positively charged head groups.40 Above 20 mN m−1, the molecular area of DODAC is even smaller than that of DODAB. Therefore, it is assumed a liquid-ordered phase with almost perpendicular orientation of the DODAC chains, which must have a larger cross-sectional area than the ones of DODAB (as confirmed by the higher wavenumbers of the CH2 stretching vibration seen in Fig. S2 – ESI†) indicating a less dense packing.
MO monolayer (Fig. 2A – blue line) agrees well with previously reported data41 and has the typical shape of a LE phase of single-chain amphiphiles, but the molecular areas are smaller than expected. MO is highly hygroscopic. Therefore, the molecular areas determined in the isotherms are apparent because of an effectively lower concentration of MO molecules in the spreading solution.
The addition of small amounts of MO to the DODAB monolayer (molar ratio 2:1) caused a shift in the isotherm towards a smaller average molecular area and the disappearance of the phase transition region. However a change of slope close to 30 mN m−1 indicates the transition into the LC phase (Fig. 2A purple dashed-dotted line).
The isotherm of the DODAC:MO (2:1) monolayer is located at larger areas per molecule (Fig. 2A pink dashed-dotted line) compared to the isotherm of DODAB:MO (2:1). The increase in the area per molecule of DODAC:MO (2:1) when compared to DODAB:MO (2:1) can be explained by the progressive screening of the monolayer surface charge and is consistent with the assumption that DODAC molecules are effectively more positively charged (due to the reduced screening capacity of the larger chloride ion in the hydrated state, which is less tightly bound to the charged head group in the inner Helmholtz plane) than DODAB molecules. Thus, DODAC has a less ordered polar region due to stronger repulsive forces between the positively charged lipid head groups. Oppositely, DODAB forms layers more densely packed at the head group region than DODAC. These different packing characteristics influence how MO is incorporated as a helper lipid into the monolayers of these cationic lipids. Plotting the molecular areas versus the mole fraction of MO indicates a non-ideal mixing between DODAB and MO at 2:1 molar ratio. On the other hand, MO eased insertion within DODAC monolayers is confirmed as the molecular areas of these two lipids as a function of the mole fraction points to an ideal mixing (data not shown).
The isotherms of the monolayers DODAC:MO (1:2) and DODAB:MO (1:2) are almost superimposed (Fig. 2A pink dotted line and purple dotted line), thus changing the counter-ion from chloride to bromide does not seem to have a significant influence on the monolayer structure at high MO contents.
The phase behaviour of the monolayer is better seen when looking at the dependence of the compression modulus (Cs−1) on the area per molecule (Fig. 2B). According to the literature,42 the variation of the Cs−1 reflects the monolayer phase transitions and three distinct regions can be observed. The regions with low Cs−1 at large area per molecule correspond to LE phases, whereas the regions with high Cs−1 at small areas per molecule correspond to LC phases. These regions are separated by a pronounced drop of the compression modulus correspondent to the LE-LC coexistence region. Accordingly, DODAB forms the least compressible monolayer (highest Cs−1 max), confirming that this monolayer also presents the highest head group ordering. The most compressible monolayer is DODAC:MO (2:1) (smallest Cs−1 max). DODAB-monolayers show higher Cs−1 max than DODAC-monolayers for the same cationic lipid:neutral lipid molar ratio. However, when MO is in excess (molar ratio 1:2), the value is the same for both systems. Monolayers containing higher contents of MO (DODAX:MO molar ratio 1:2) are not very different in terms of compressibility when compared to the neat MO monolayer.
Since DODAB was the only monolayer forming the LE, LE + LC and LC phases, BAM images are shown for this composition in Fig. 2C. At maximum expansion, a 2D foam often appears for the DODAB monolayer, and a homogeneous BAM image is recovered after the lift-off point at a low surface pressures (Fig. 2C, LE). BAM images of the DODAB monolayer revealed the presence of small domains showing ‘bubble-like’ features (Fig. 2C, LE + LC), that progressively became more densely packed (Fig. 2C, LC) until the image gets homogeneous upon further compression (data not shown). This behaviour indicates a large nucleation rate and a slow growth of the condensed domains which makes difficult to compare the shape of the domains in the different mixed monolayers. However it is possible to observe that at the smaller MO contents (DODAX:MO (2:1)) the same type of domains appeared for DODAB:MO while the mixed DODAC:MO monolayer appeared homogeneous when visualized by BAM (Fig. S4 – ESI†). This confirms that MO is more homogeneously distributed in the case of DODAC, but forms phase-separated DODAB- and MO-enriched domains in DODAB aggregates.
Analysis of DODAX:MO mixtures in 3D assemblies by confocal Raman microscopy
Raman spectra of DODAC:MO (2:1), DODAB:MO (2:1), DODAC:MO (1:2) and DODAB:MO (1:2) liposome dispersions are shown in Fig. 3 for the range between 1000 and 3100 cm−1.
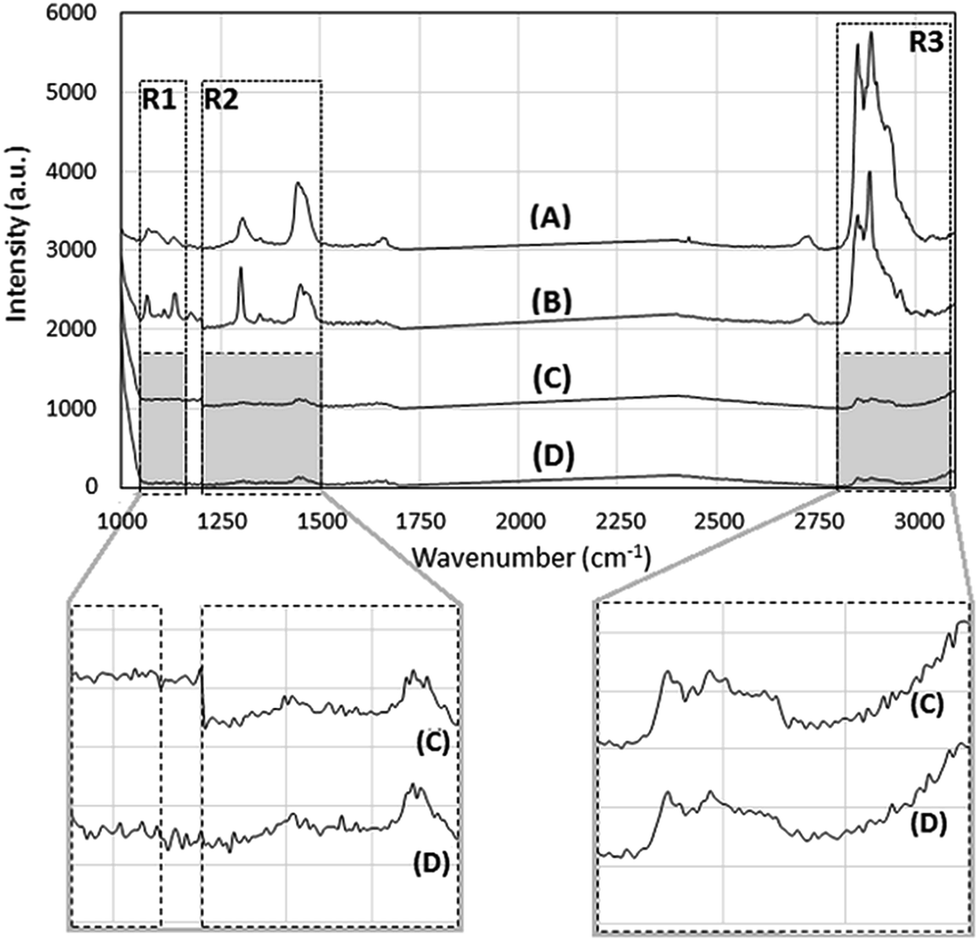 | ||
| Fig. 3 Raman spectra of DODAC:MO (2:1) (A), DODAB:MO (2:1) (B), DODAC:MO (1:2) (C) and DODAB:MO (1:2) (D) liposomes. Insets show the spectra of DODAC:MO (1:2) (C) and DODAC:MO (1:2) (D) with regions R1, R2 and R3 in more detail. | ||
The Raman spectra obtained are in good agreement with the reported for other lipid composition showing the ν(C–H), ν(C–C) and δ(C–H) vibration modes.43
The most relevant vibrational mode frequencies, obtained after fitting each Raman spectrum, are shown in Table 1. For better clarity, 3 regions of the spectra (R1, R2 and R3) were defined and analysed separately: R1 (1050–1150 cm−1) comprising the ν(C–C) stretching vibrations, R2 (1200–1500 cm−1) comprising the δ(CH2) deformation vibrations, and R3 (2800–3100 cm−1) corresponding to the ν(C–H) stretching vibrations, which dominate the spectra.
| DODAC:MO (2:1) |
DODAB:MO (2:1) |
DODAC:MO (1:2) |
DODAB:MO (1:2) |
Assignmenta | |
|---|---|---|---|---|---|
| a ν – stretch; δ – bend and or scissor/deformations. T – trans, G – gauche, ip – in-phase, tw – twist, sc – scissor, s – symmetric, a – antisymmetric. | |||||
| R1 C–C stretching region | 1068 | 1066 | 1068 | 1064 | ν(C–C)T |
| 1083 | 1084 | 1085 | 1088 | ν(C–C)G | |
| 1107/1135 | 1109/1135 | 1107/1134 | 1107/1134 | ν(C–C)ip | |
| R2 C–H deformation region | 1305 | 1301 | — | — | δ(CH2)tw |
| 1440/1448/1462 | 1450/1464/1474 | 1438/1446/1464/1484 | 1438/1445/1458/1476 | δ(CH2)sc | |
| 1657/1665 | 1646/1667 | — | 1644/1667 | ν(C![[double bond, length as m-dash]](https://www.rsc.org/images/entities/b_char_e001.gif) C) C) |
|
| R3 C–H stretching region | 2855 | 2855 | 2855 | 2855 | νs(CH2) |
| 2888 | 2885 | 2889 | 2884 | νa(CH2) | |
| 2935 | 2935 | 2935 | — | νs(CH3) | |
| 2965 | 2963 | — | 2962 | νa(CH3) | |
Fig. 3 shows that all the vibration modes were more intense for DODAX:MO liposomes prepared at the (2:1) molar ratio (Fig. 3A and B) when compared to the (1:2) molar ratio (Fig. 3C and D).
The ν(C–C) stretching mode (in region R1) reflects intramolecular trans/gauche conformational changes within hydrocarbon chains. According to the literature1,43–45 the out-of-phase aliphatic C–C stretch all-trans vibrations ν(C–C)T are observed between 1060 and 1065 cm−1; the C–C stretch gauche vibrations ν(C–C)G correspond to the Raman mode at ≈1086 cm−1; and the in-phase aliphatic C–C stretch all-trans vibrations ν(C–C)ip appear between 1100 and 1135 cm−1.
Fig. 3 shows that the lipid systems with 2:1 molar ratios DODAB:MO (B spectra) and DODAC:MO (A spectra) present a well-defined Raman band assigned for ν(C–C)T at 1066 cm−1, however being shifted to higher wavenumbers and broadened for DODAC:MO (2:1) liposomes. This ν(C–C)T band presents very low intensity in both lipid mixtures containing higher MO content (molar ratio 1:2). The ν(C–C)G vibration peak appeared poorly defined for DODAB:MO (2:1) (1084 cm−1) and DODAC:MO (2:1) (1083 cm−1), and again presented very low intensity for lipid mixtures containing higher MO content.
The ν(C–C)ip modes in the range 1100 and 1135 cm−1, corresponding to the in-phase aliphatic C–C stretch all-trans, were well-defined for the lipid systems with 2:1 molar ratios DODAB:MO (B spectra) and DODAC:MO (A spectra), however, the low frequency mode is shifted (to lower wavenumbers) and broadened for DODAC:MO (2:1). The contribution of ν(C–C)ip vibrations was almost irrelevant for DODAX:MO (1:2) liposomes.
The Raman spectrum in the R2 region (1200 to 1500 cm−1) corresponds to the C–H deformation vibrations δ(CH2). These modes are very sensitive to lipid chain architecture and lateral packing. The methylene twisting deformation vibrations δ(CH2)tw are observed between 1295 and 1305 cm−1, and the methylene scissor deformation vibrations δ(CH2)sc between 1400 and 1500 cm−1.1
As seen in Fig. 3, the δ(CH2)tw mode is well defined for DODAB:MO (2:1) and DODAC:MO (2:1), but the peak position was shifted and the mode is broader for the latter (peaks at 1301 cm−1 and 1305 cm−1, respectively). Moreover, this peak is very weak for lipid mixtures containing higher MO content (molar ratio 1:2) indicating that the lateral packing is very much reduced at high MO contents.
The δ(CH2)sc (1400–1500 cm−1) modes were also well-defined for DODAX:MO (2:1). It is interesting to note the intensity inversion between the 1301 cm−1 (δ(CH2)tw) and 1400–1500 cm−1 (δ(CH2)sc) modes, when comparing DODAB:MO (2:1) and DODAC:MO (2:1), as well as the red and blue shift of the peak position for DODAC:MO (2:1).
The C–H stretching vibration region R3 (2800–3100 cm−1) is the most sensitive region and the ν(CH2) vibrations are usually used to monitor the changes in the lipid chain lateral packing, or order/disorder properties.46 The Raman spectra intensity in the region R3 was higher for DODAC:MO (2:1), followed by DODAB:MO (2:1), while it was very low and poorly defined for the mixtures with higher MO content (molar ratio 1:2), as observed in the others vibrational ranges. The methylene symmetric stretching vibration (νs(CH2)) appeared at 2855 cm−1 and maintained its position for all the samples, although the intensity varied in the order DODAC:MO (2:1) > DODAB:MO (2:1) > DODAX:MO (1:2). The νs(CH3) vibration mode at 2935 cm−1 was better defined for DODAC:MO (2:1) while the νa(CH3) vibration mode at ≈2963 cm−1 was better defined for DODAB:MO (2:1). These two contributions seemed irrelevant for formulations with higher MO content.
Ratios between defined peaks of Raman spectra provide information regarding different bilayer characteristics. For example, the peak intensity ratio [IνC–CG/IνC–CT] indicates the relative number of gauche and trans conformers, and thus it can be used to monitor changes in membrane order for the C–C stretching mode region.43 The [IνaCH2/IνsCH2] peak height intensity ratio is a measure of lateral packing density of alkyl chains and an indicator of conformational order, reflecting deviations from a perfect all-trans conformation.46,47 The ratio lies between 0.6 and 0.9 for alkanes in the liquid state, and between 1.6 and 2.0 for the crystalline state.48,49 Also, the higher the ratio, the more ordered the hydrocarbon chains.50 The [IνsCH3/IνsCH2] peak height intensity ratio is correlated to the chain coupling (intermolecular interactions) of alkyl chains.51 This ratio is sensitive to intermolecular chain interactions and also reflects the mobility of the terminal methyl group of the hydrocarbon chains.
Fig. 4 shows the intensity ratio [IνC–CG/IνC–CT] as a function of [IνaCH2/IνsCH2], and [IνsCH3/IνsCH2] as a function of [IνaCH2/IνsCH2].
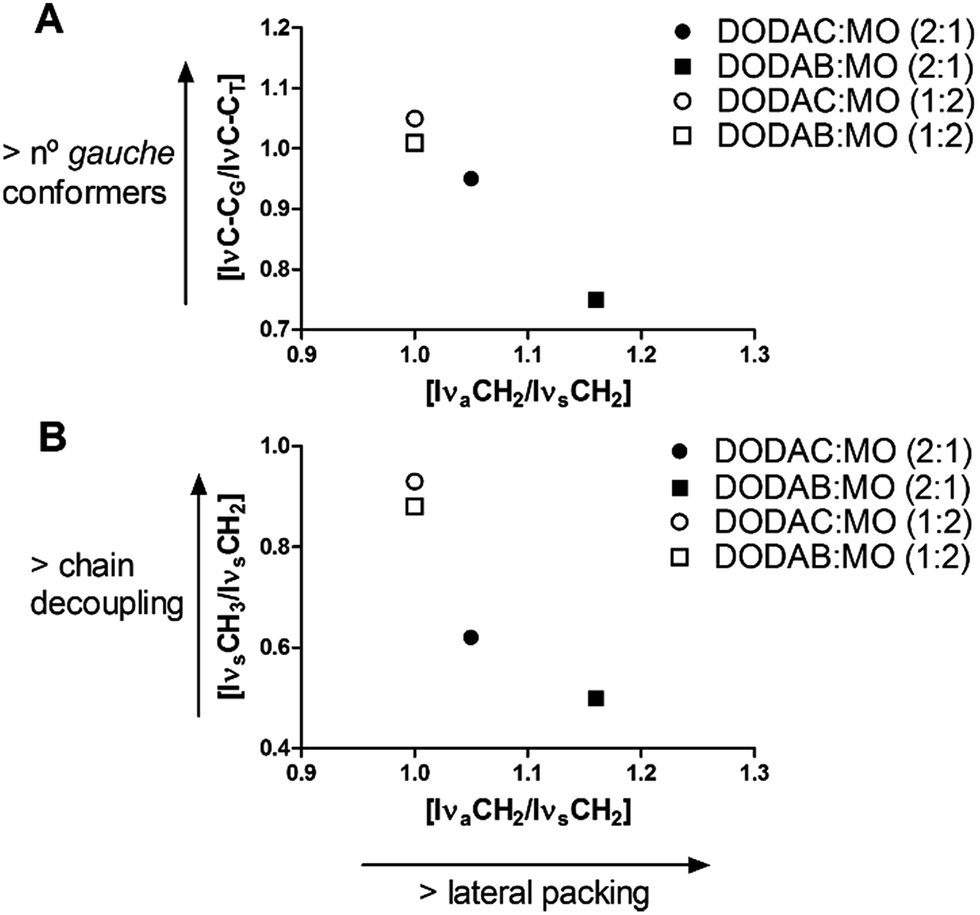 | ||
| Fig. 4 Intensity ratio [IνC–CG/IνC–CT] as function of [IνaCH2/IνsCH2] (A) and [IνsCH3/IνsCH2] as function of [IνaCH2/IνsCH2] (B). | ||
Fig. 4A indicates how the number of gauche conformers is related to the lateral packing density of the acyl chains. A higher [IνC–CG/IνC–CT] ratio reflects a decrease in the order of the acyl chains, since it indicates an increase in gauche conformers. Contrarily, a higher [IνaCH2/IνsCH2] ratio indicates an increase in conformational order. DODAB:MO (2:1) presented the lowest value of [IνC–CG/IνC–CT] (0.75) and the highest value of [IνaCH2/IνsCH2] (1.16), followed by DODAC:MO (2:1) ([IνC–CG/IνC–CT] = 0.95 and [IνaCH2/IνsCH2] = 1.05) which means that DODAB:MO (2:1) has a more densely packed bilayer, closer to a perfect all-trans state.
Similar values of [IνaCH2/IνsCH2] ratio (1.00) were found for DODAB:MO (1:2) and DODAC:MO (1:2) liposomes. Thus, when MO was in excess in the formulations (molar ratio 1:2), the presence of different counter-ions does not have an influence on the lateral packing as observed for (2:1) molar ratios. Therefore, both DODAC:MO and DODAB:MO (1:2) systems possess great content of gauche conformers and lower lateral packing density. Although DODAX:MO (1:2) had the same [IνaCH2/IνsCH2] ratio, DODAC:MO (1:2) presented a slightly higher value of [IνC–CG/IνC–CT] than DODAB:MO (1:2) (1.05 > 1.00). Hence, generally speaking and for the same molar ratio, DODAC-based liposomes have more gauche conformers than DODAB-based liposomes, which means that the perturbation of DODAC membranes by MO is mainly driven by a change in the gauche/trans ratio of the acyl chains.
Fig. 4B shows how chain decoupling (that reflects the mobility of the methyl terminal group of hydrocarbon chains) and lateral packing density of the acyl chains are related. Higher [IνsCH3/IνsCH2] ratios indicate higher decoupling and more mobile CH3, which increase the molar volume of the molecule.
As expected, DODAB:MO and DODAC:MO (1:2) had higher chain decoupling ratios, since that higher MO content promotes a fluidizing effect at the membrane level. We nevertheless observed that, for the same molar ratio, DODAC-based liposomes had higher [IνsCH3/IνsCH2] values indicating a more mobile terminal methylene end group [IνsCH3/IνsCH2] = 0.62 for DODAC:MO (2.1) versus [IνsCH3/IνsCH2] = 0.50 for DODAB:MO (2:1) and [IνsCH3/IνsCH2] = 0.93 for DODAC:MO (1:2) versus [IνsCH3/IνsCH2] = 0.88 for DODAB:MO (1:2). The higher chain decoupling obtained for DODAC based liposomes indicates that DODAC hydrocarbon chains interact less strongly than DODAB chains, especially for (1:2) molar ratio. This is in agreement with the gauche conformer increase observed for DODAC:MO liposomes (higher values of [IνC–CG/IνC–CT]). Langmuir π–A isotherms are also in agreement with the Raman spectroscopy data, indicating a higher packing density for DODAB:MO (2:1) than DODAC:MO (2:1) monolayers as shown by the higher compressibility modulus (Fig. 2B). This is again related to the capability of bromide counter-ions to screen the cationic head group charge better than chloride counter-ions, thus increasing the head group packing density. As a consequence, insertion of MO into DODAB bilayers is more difficult leading to demixing of the compounds into distinctive DODAB- and MO-rich domains in the corresponding mixtures. Also, in both 3D and 2D systems, the increase in MO content (DODAX:MO (1:2)), reduced the influence that the counter-ions had on the aggregate structures. Furthermore, the effect of MO content in chain decoupling is also consistent with previous results where an increase in MO content was correlated with an increase in membrane fluidity.17
Apart from the three regions of the Raman spectra previously defined, there is another region correspondent to the ν(C![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) C) vibrations of unsaturated chain segments existent in MO (Fig. 1), that appears around 1660 cm−1 (1654 cm−1 for cis and 1669 cm−1 for trans).44 The ν(CC) vibration bands were clearly visible for DODAC:MO (2:1) at 1657 cm−1 and 1665 cm−1, but were very weak or absent for the other formulations (Fig. 3 and Table 1).
C) vibrations of unsaturated chain segments existent in MO (Fig. 1), that appears around 1660 cm−1 (1654 cm−1 for cis and 1669 cm−1 for trans).44 The ν(CC) vibration bands were clearly visible for DODAC:MO (2:1) at 1657 cm−1 and 1665 cm−1, but were very weak or absent for the other formulations (Fig. 3 and Table 1).
This can be due to the different distribution of MO in DODAX:MO (2:1) membranes. In DODAC:MO (2:1) membranes, MO is more homogeneously distributed, thus increasing the fluidity, as observed by an increased number of gauche conformers and a smaller lateral packing density (Fig. 4). The smaller lateral packing density explains in turn the fact of unsaturated CC stretching vibrations being more evident in DODAC:MO (2:1) due to higher molecular freedom. In the case of DODAB:MO (2:1) membranes, MO is more heterogeneously distributed (phase-separated), thus increasing the fluidity in the regions where it is located but imposing constraints to the neighboring DODAB lipid molecules. This can be translated into a reduced number of gauche conformers and in an average higher lateral packing density (Fig. 4).
Analysis of DODAX:MO mixtures in 3D assemblies by small angle X-ray scattering (SAXS)
3D assemblies of DODAX:MO mixtures were also investigated by SAXS at 20 °C. SAXS curves for 40 mM DODAB (Fig. 5A) show two well-resolved Bragg peaks that can be assigned to the first and third order reflections of a lamellar structure (q3:q1 = 3:1).
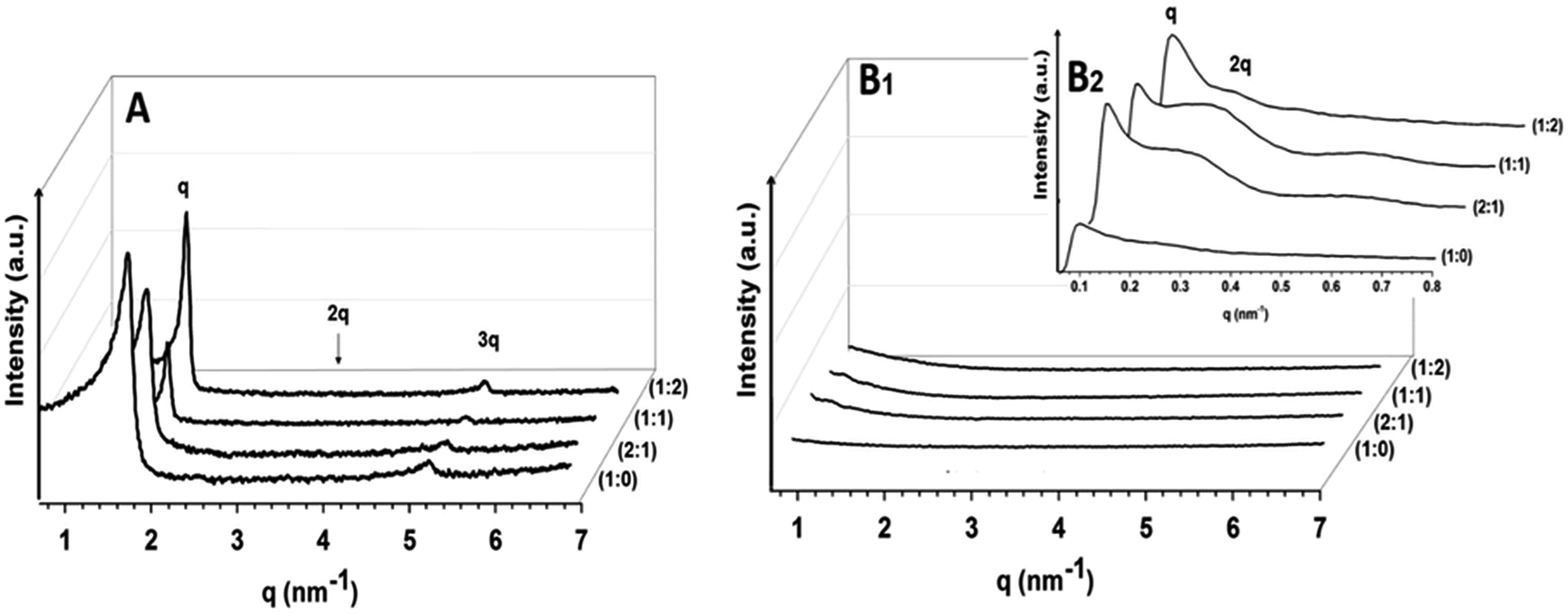 | ||
| Fig. 5 (A) SAXS curves for DODAB:MO (40 mM) in water at 20 °C, obtained at (1:0); (2:1); (1:1) and (1:2) ratios. (B) SAXS curves for DODAC:MO (40 mM) in water at 20 °C, obtained at (1:0); (2:1); (1:1) and (1:2) ratios at higher q values (B1) and at lower q values (B2). q, 2q and 3q indicate the first-, second- and third-order of reflection. | ||
The first and most intense reflection at q1 of 1.73 nm−1 corresponds to a Bragg distance of d = 36.3 Å comprising the lipid bilayer thickness plus the enclosed water layer. The third reflection order at q3 of 5.20 nm−1 is much less intense and fits well with the position expected for a lamellar phase, for which the second order reflection expected at q2 = 3.46 nm−1 is absent.
The increase of MO content does not change the observed DODAB structure (first-order reflection at q1 = 1.74 nm−1, and a higher-order reflection at 3q1 (q3 = 5.2 nm−1)). The second-order reflection expected at 2q1 (q2 at 3.5 nm−1) is also absent (Fig. 5A). Missing reflection orders are reported for well-defined bilayer structures and are the result of the interplay between structure and form factors.52 If the bilayers have slightly different thicknesses, then the even-order peaks will be present.53 This is in agreement with the fact that the DODAB assemblies are well-ordered with no bilayer thickness fluctuations.
While DODAB assemblies presented a well-ordered lamellar structure (Fig. 5A), in the same q range, SAXS scattering data of DODAC and the corresponding mixtures with MO do not show any Bragg peaks (Fig. 5B1) of a bilayer arrangement at least up to the concentrations tested in the current work (40 mM). The absence of diffraction peaks can, in this case, be associated with the formation of unilamellar vesicles (ULVs), as described previously for DODAAC (another DODAX cationic lipid where the counter-ion is the acetate) at concentrations up to 50 mM.53 Indeed, as previously stated, the bigger hydration shell of the chloride counter-ion leads to a less effective screening of the positive charge of DODAC head groups. The charged head groups are repelled by electrostatic interactions and preferentially form ULVs that are weak scatterers compared to aligned bilayers and MLVs.54–56
The formation of ULVs of DODAC:MO can be confirmed by the observation of diffraction peaks at much smaller q values (Fig. 5B2) with the first-order of reflection at q1 of 0.112 nm−1 and a higher-order reflection at q2 of 0.231 nm−1 that fits well with a peak position expected for the second-order reflection of a lamellar structure (q2:q1 = 2:1). These diffraction peaks are consistent with other SAXS diffraction studies using ULVs of 50 nm size that presented the first-order reflection at low q values (0.1 nm−1).55,56 The Bragg peaks obtained for DODAC:MO mixtures are however less intense and more diffuse than the ones obtained with DODAB:MO mixtures for higher q values. These observations are consistent with transient stages of the structural transformation from swollen lamellae to cubosomal intermediates that have been previously noted for MO and several other amphiphiles including monoelaidin (ME) and mixtures of Lauric Acid (LA) and di-laurylphosphatidylcholine (DLPC) (LA:DLPC 2:1).57–60
From the Bragg peaks positions the values of interlayer spacing d obtained for 3D lamellar structures of DODAX:MO at different cationic lipid:neutral lipid molar ratios were calculated. In the case of DODAB:MO the introduction of MO does not significantly change the interlayer spacing of the system. Indeed, the values of the bilayer thickness plus the water layer obtained for every DODAB:MO molar ratio (d = 36.0 Å) are in good agreement with previously data reported for neat DODAB without MO.61,62 The correlation length (ξ) of the bilayers is not affected by the inclusion of lower contents of MO (ξ = 555 Å for DODAB:MO (1:0) and DODAB:MO (2:1)), being however strongly increased for higher MO contents (ξ > 900 Å for DODAB:MO (1:1) and DODAB:MO (1:2)).
In the case of DODAC:MO much larger d-spacings and lower correlations lengths (ξ) compared to DODAB:MO are observed. The much higher d-spacings are consistent with the previously reported study of neat DODAC, and also explained by the authors as being due to a pronounced electrostatic repulsion between the highly charged DODAC bilayers in the absence of additional electrolyte.63 The insertion of MO into the DODAC bilayers is connected with a significant increase in the repeating distance consistent with the fluid Lα phase of closely packed onion-like vesicles confining a highly swollen cubic phase that have been reported as membrane fusion intermediates.57–60
Correlation between biophysical characteristics of the liposomes DODAX:MO, evaluated in this work and the resultant biological performance of their lipoplexes
The information gathered with all the biophysical techniques used within this work (Langmuir isotherms, BAM, IRRAS, Raman scattering and SAXS experiments) and with previously reported cryo-TEM24 and cryo-SEM14 validate a proposed model for DODAX:MO assemblies that depends of the counter-ion and MO content (Fig. 6).
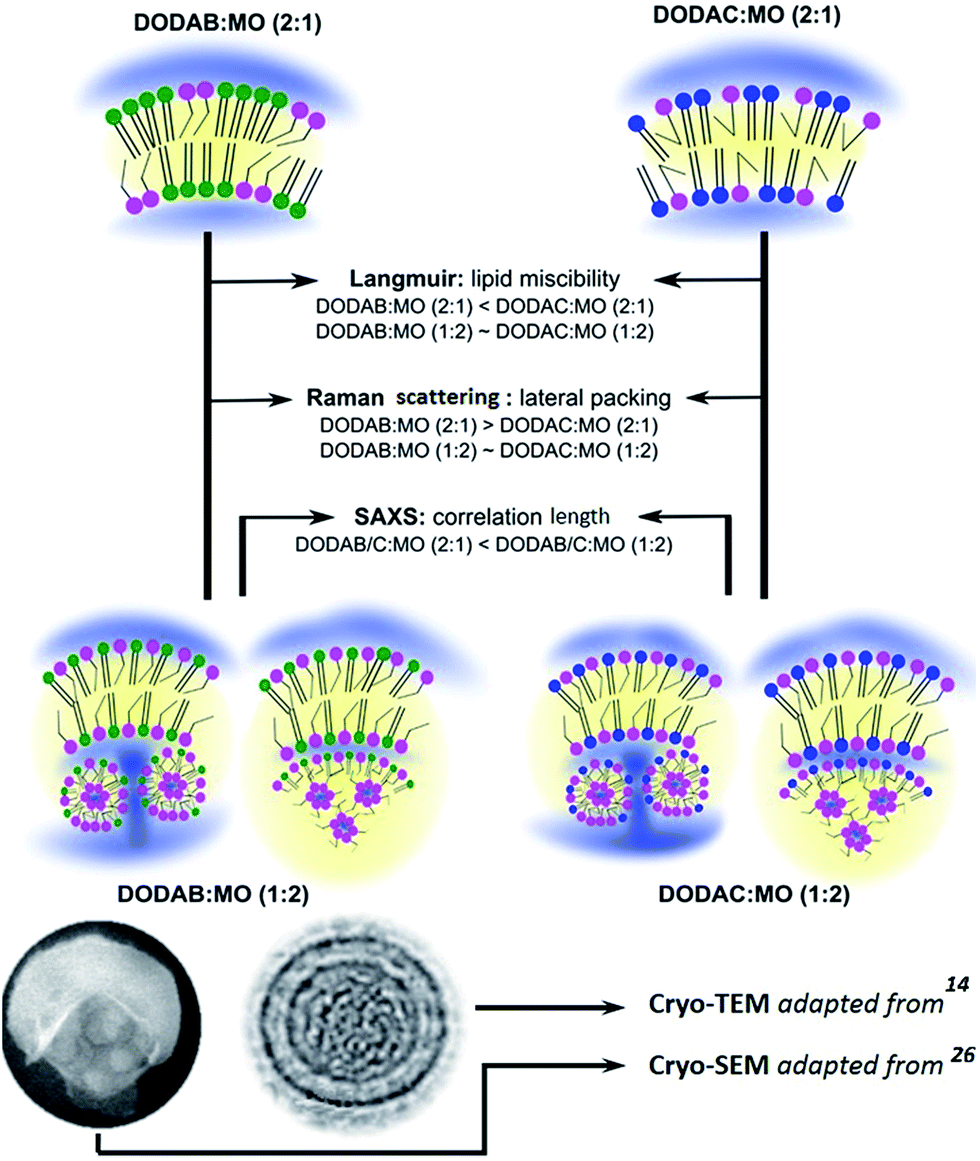 | ||
| Fig. 6 Schematic representation of the main results obtained for DODAB:MO (2:1), DODAC:MO (2:1), DODAB:MO (1:2) and DODAC:MO (1:2) liposomes by Langmuir isotherms, Raman scattering and SAXS experiments. Previously reported cryo-TEM14 and cryo-SEM26 micrographs of DODAB:MO with high MO ratios validate the proposed model. | ||
In the systems with smaller MO content, DODAX:MO (2:1), there is a higher influence of the counter-ion. Chloride ions are less able to effectively screen the positive charges of the cationic lipid head group in DODAC containing systems. This leads to a less ordered polar region due to stronger repulsive forces between the positively charged lipid head groups. As a result DODAC:MO (2:1) systems are less densely packed facilitating a homogeneous distribution of MO into DODAC bilayers (Fig. 6). DODAB:MO (2:1) systems are more densely packed at the head group region, and thus MO distribution within the bilayers is not as homogeneous, forming phase-separated DODAB- and MO-enriched domains (Fig. 6).
The lower screening of the positive charges of the cationic lipid head group in DODAC explains why DODAC:MO (2:1) liposomes reached total siRNA complexation at lower charge ratios (C.R. (+/−) = 2.5) when compared to DODAB:MO (2:1) liposomes (C.R. (+/−) = 5) (Fig. S1A – ESI†). The fact that DODAB bilayers are more densely packed and do not incorporate MO as homogeneously as DODAC bilayers also explains the higher resistance to destabilization by serum proteins achieved for DODAB:MO based lipoplexes (Fig. S1B – ESI†). Furthermore DODAC:MO (2:1) systems originate lipoplexes with less transfection efficiency than DODAB:MO (2:1) systems (Fig. S1C – ESI†). Given that both systems present the same MO content, the differences presented must be correlated with the MO distribution. MO is more homogeneously distributed in the case of DODAC bilayers, but forms phase-separated DODAB- and MO-enriched domains in DODAB based systems. These MO-enriched domains among DODAB domains might facilitate the adherence and fusion of the lipoplexes to the surfaces of the cells improving transfection.64
When MO was in excess in the formulations (molar ratio 1:2), the presence of different counter-ions did not have such a strong influence in terms of lateral packing as observed for (2:1) molar ratios. This indicates that saturation of bilayers with MO leads to disruption of DODAB-rich domains, making DODAB:MO and DODAC:MO bilayers more alike (Fig. 6). As previously observed by cryo-TEM24 and cryo-SEM,14 exceeding MO seems to be excluded from the liposomal bilayer to the liposomal core where it might form MO-rich inverted non-lamellar structures (also observed by other authors in lipid mixtures containing MO12,65) or MO containing vesicles (Fig. 6).
As the systems with higher MO content DODAX:MO (1:2) are similar in terms of MO distribution (Fig. 6) their performance as lipoplexes has shown to be similar as well. Hence, DODAB:MO (1:2) and DODAC:MO (1:2) based lipoplexes present comparable transfection efficiencies (Fig. S1C – ESI†).
Conclusions
This work describes the detailed characterization of the influence of the counter-ion and MO content on DODAX:MO structural organization. The results found for DODAX:MO (2:1) systems can be summarized as follows: (i) DODAB:MO (2:1) forms bilayers more densely packed at the head group region than DODAC:MO (2:1) bilayers; (ii) MO is more homogeneously distributed in DODAC-based bilayers, but forms phase-separated DODAB- and MO-enriched domains in DODAB aggregates; (iii) chloride and bromide counter-ions induce different repulsive interactions at the head groups' level due to differences in the surfactant hydration shell. E.g. the chloride ion has a larger hydration shell, resulting in lower screening of the positive charges of the cationic lipid head group in DODAC. This leads to a less ordered polar region due to stronger repulsive forces between the positively charged lipid head groups, facilitating a homogeneous distribution of MO into DODAC bilayers. DODAX:MO (1:2) systems are more similar because the excess of MO reduces the effect of the counter-ion.
The differences observed between the formulations studied are correlated with key processes such as lipoplex assembly/disassembly or colloidal stability, ultimately influencing lipid-mediated transfection efficiency. Therefore, this work highlights the importance of a detailed biophysical characterization of lipoplexes' lipid components to better understand their biological outcomes and to aid scientists in the rational design of new and efficient gene therapy nanocarriers.
Acknowledgements
We acknowledge DAAD/FCT that provided the financial support required to gather the Portuguese and the German co-workers. This work was further supported by FEDER through POFC-COMPETE and by national funds from FCT, through the projects PEst-OE/BIA/UI4050/2014 (CBMA) and PEst-C/FIS/UI0607/2013 (CFUM). Marlene Lúcio acknowledges FCT for the financial support provided by the exploratory project IF/00498/2012. C. R.-A. is grateful to the European Union through the Operational Programme for Cross-border Cooperation: Spain-Portugal under Grant POCTEP 2007–2013 and to European Regional Development Fund for research funding (Innovation in Nanomedicine Project). The authors would also like to acknowledge Irina Berndt and Claudia Botelho.References
- J. R. Beattie, S. E. Bell and B. W. Moss, Lipids, 2004, 39, 407–419 CrossRef CAS PubMed.
- C. J. Orendorff, M. W. Ducey and J. E. Pemberton, J. Phys. Chem. A, 2002, 106, 6991–6998 CrossRef CAS.
- T. M. Allen and P. R. Cullis, Adv. Drug Delivery Rev., 2013, 65, 36–48 CrossRef CAS PubMed.
- D. A. Balazs and W. T. Godbey, J. Drug Delivery, 2011, 2011, 326497 Search PubMed.
- S. Zhang, Y. Xu, B. Wang, W. Qiao, D. Liu and Z. Li, J. Controlled Release, 2004, 100, 165–180 CrossRef CAS PubMed.
- H. Farhood, N. Serbina and L. Huang, Biochim. Biophys. Acta, 1995, 1235, 289–295 CrossRef.
- I. Koltover, T. Salditt, J. O. Radler and C. R. Safinya, Science, 1998, 281, 78–81 CrossRef CAS PubMed.
- Y. Xu and F. C. Szoka, Biochemistry, 1996, 35, 5616–5623 CrossRef CAS PubMed.
- N. J. Zuidam and Y. Barenholz, Biochim. Biophys. Acta, 1998, 1368, 115–128 CrossRef CAS.
- M. J. Blandamer, B. Briggs, P. M. Cullis, B. J. Rawlings and J. B. F. N. Engberts, Phys. Chem. Chem. Phys., 2003, 5, 5309–5312 RSC.
- F. Koster, D. Finas, C. Schulz, C. Hauser, K. Diedrich and R. Felberbaum, Int. J. Mol. Med., 2004, 14, 769–772 Search PubMed.
- B. Angelov, A. Angelova, M. Drechsler, V. M. Garamus, R. Mutafchieva and S. Lesieur, Soft Matter, 2015, 11, 3686–3692 RSC.
- B. Angelov, A. Angelova, S. K. Filippov, G. Karlsson, N. Terrill, S. Lesieur and P. Stepanek, Soft Matter, 2011, 7, 9714–9720 RSC.
- C. Carneiro, A. Correia, T. Collins, M. Vilanova, C. Pais, A. C. Gomes, M. E. Real Oliveira and P. Sampaio, Eur. J. Pharm. Biopharm., 2015, 89, 190–200 CrossRef CAS PubMed.
- J. P. Neves Silva, P. J. Coutinho and M. E. Real Oliveira, J. Fluoresc., 2008, 18, 555–562 CrossRef CAS PubMed.
- J. P. Silva, A. C. Oliveira, M. P. Casal, A. C. Gomes, P. J. Coutinho, O. P. Coutinho and M. E. Oliveira, Biochim. Biophys. Acta, 2011, 1808, 2440–2449 CrossRef CAS PubMed.
- J. P. Silva, A. C. Oliveira, M. Lucio, A. C. Gomes, P. J. Coutinho and M. E. Oliveira, Colloids Surf., B, 2014, 121, 371–379 CrossRef CAS PubMed.
- J. P. Silva, I. M. Oliveira, A. C. Oliveira, M. Lucio, A. C. Gomes, P. J. Coutinho and M. E. Oliveira, Biochim. Biophys. Acta, 2014, 1838, 2555–2567 CrossRef CAS PubMed.
- J. P. N. Silva, A. C. N. Oliveira, M. Lúcio, A. F. C. Gomes and M. E. C. D. Real Oliveira, J. Appl. Solution Chem. Model., 2014, 3, 94–105 CAS.
- J. P. Neves Silva, A. C. N. Oliveira, A. C. Gomes and M. E. C. D. Real Oliveira, Development of Dioctadecyldimethylammonium Bromide/Monoolein Liposomes for Gene Delivery, InTech, Rijeka (Croatia), 2012 Search PubMed.
- J. Briggs and M. Caffrey, Biophys. J., 1994, 67, 1594–1602 CrossRef CAS PubMed.
- H. Qiu and M. Caffrey, Biomaterials, 2000, 21, 223–234 CrossRef CAS PubMed.
- A. Deshpande, P. Sicinski and P. W. Hinds, Oncogene, 2005, 24, 2909–2915 CrossRef CAS PubMed.
- C. V. Kulkarni, W. Wachter, G. Iglesias-Salto, S. Engelskirchen and S. Ahualli, Phys. Chem. Chem. Phys., 2011, 13, 3004–3021 RSC.
- A. Ganem-Quintanar, D. Quintanar-Guerrero and P. Buri, Drug Dev. Ind. Pharm., 2000, 26, 809–820 CrossRef CAS PubMed.
- I. M. Oliveira, J. P. Silva, E. Feitosa, E. F. Marques, E. M. Castanheira and M. E. Real Oliveira, J. Colloid Interface Sci., 2012, 374, 206–217 CrossRef CAS PubMed.
- E. Feitosa and F. R. Alves, Chem. Phys. Lipids, 2008, 156, 13–16 CrossRef CAS PubMed.
- A. C. Oliveira, T. F. Martens, K. Raemdonck, R. D. Adati, E. Feitosa, C. Botelho, A. C. Gomes, K. Braeckmans and M. E. Real Oliveira, ACS Appl. Mater. Interfaces, 2014, 6, 6977–6989 CAS.
- P. Dynarowicz-Latka, A. Dhanabalan and O. N. Oliveira Jr, Adv. Colloid Interface Sci., 2001, 91, 221–293 CrossRef CAS PubMed.
- M.-H. Ropers and G. Brezesinski, Soft Matter, 2013, 9, 9440–9448 RSC.
- F. Bonosi and G. Gabrielli, Colloids Surf., 1991, 52, 277–285 CrossRef CAS.
- A. M. Gonçalves da Silva and R. I. S. Romão, Chem. Phys. Lipids, 2005, 137, 62–76 CrossRef PubMed.
- K. Hąc-Wydro, P. Wydro and P. Dynarowicz-Łątka, J. Colloid Interface Sci., 2005, 286, 504–510 CrossRef PubMed.
- D. M. Taylor, Y. Dong and C. C. Jones, Thin Solid Films, 1996, 284–285, 130–133 CrossRef CAS.
- I. Petrov, D. Moebius and A. Angelova, Langmuir, 1992, 8, 201–205 CrossRef CAS.
- E. Feitosa, F. R. Alves, E. M. S. Castanheira and M. E. C. D. Real Oliveira, Colloid Polym. Sci., 2009, 287, 591–599 CAS.
- C. K. Liu and G. G. Warr, Soft Matter, 2014, 10, 83–87 RSC.
- K. M. McGrath, Langmuir, 1995, 11, 1835–1839 CrossRef CAS.
- V. L. Shapovalov and G. Brezesinski, J. Phys. Chem. B, 2006, 110, 10032–10040 CrossRef CAS PubMed.
- M. N. Antipina, B. Dobner, O. V. Konovalov, V. L. Shapovalov and G. Brezesinski, J. Phys. Chem. B, 2007, 111, 13845–13850 CrossRef CAS PubMed.
- V. Kolev, A. Ivanova, G. Madjarova, A. Aserin and N. Garti, J. Chem. Phys., 2012, 136, 074509 CrossRef PubMed.
- A. P. Dabkowska, D. J. Barlow, A. V. Hughes, R. A. Campbell, P. J. Quinn and M. J. Lawrence, J. R. Soc., Interface, 2012, 9, 548–561 CrossRef CAS PubMed.
- C. B. Fox, R. A. Horton and J. M. Harris, Anal. Chem., 2006, 78, 4918–4924 CrossRef CAS PubMed.
- F. N. R. Petersen and C. H. Nielsen, Spectroscopy, 2009, 24, 1–7 Search PubMed.
- J. M. Sanderson and A. D. Ward, Chem. Commun., 2004, 1120–1121, 10.1039/b316757g.
- K. Larsson and R. P. Rand, Biochim. Biophys. Acta, 1973, 326, 245–255 CrossRef CAS.
- R. G. Snyder, S. L. Hsu and S. Krimm, Spectrochim. Acta, Part A, 1978, 34, 395–406 CrossRef.
- D. Lin-Vien, N. B. Colthup, W. G. Fateley and J. G. Grasselli, in The Handbook of Infrared and Raman Characteristic Frequencies of Organic Molecules, Academic Press, San Diego, 1991, pp. 9–28, DOI: 10.1016/B978-0-08-057116-4.50008-0 Search PubMed.
- D. F. H. Wallach, S. P. Verma and J. Fookson, Biochim. Biophys. Acta, Rev. Biomembr., 1979, 559, 153–208 CrossRef CAS.
- M. Prochazka, J. Stepanek and P. Y. Turpin, Chem. Phys. Lipids, 2004, 132, 145–156 CrossRef CAS PubMed.
- R. G. Snyder, H. L. Strauss and C. A. Elliger, J. Phys. Chem., 1982, 86, 5145–5150 CrossRef CAS.
- C. Suryanarayana and M. G. Norton, X-Ray Diffraction – A practical approach, Springer Sciences+Business Media, 1998 Search PubMed.
- F. R. Alves and W. Loh, J. Colloid Interface Sci., 2012, 368, 292–300 CrossRef CAS PubMed.
- M. R. Brzustowicz and A. T. Brunger, J. Appl. Crystallogr., 2005, 38, 126–131 CrossRef CAS.
- N. Kucerka, Y. Liu, N. Chu, H. I. Petrache, S. Tristram-Nagle and J. F. Nagle, Biophys. J., 2005, 88, 2626–2637 CrossRef CAS PubMed.
- N. Kucerka, J. Pencer, J. N. Sachs, J. F. Nagle and J. Katsaras, Langmuir, 2007, 23, 1292–1299 CrossRef CAS PubMed.
- C. E. Conn, O. Ces, X. Mulet, S. Finet, R. Winter, J. M. Seddon and R. H. Templer, Phys. Rev. Lett., 2006, 96, 108102 CrossRef PubMed.
- J. Kraineva, R. A. Narayanan, E. Kondrashkina, P. Thiyagarajan and R. Winter, Langmuir, 2005, 21, 3559–3571 CrossRef CAS PubMed.
- A. M. Squires, R. H. Templer, J. M. Seddon, J. Woenckhaus, R. Winter, S. Finet and N. Theyencheri, Langmuir, 2002, 18, 7384–7392 CrossRef CAS.
- A. Yaghmur, P. Laggner, M. Almgren and M. Rappolt, PLoS One, 2008, 3, e3747 Search PubMed.
- E. Feitosa, R. D. Adati, P. Hansson and M. Malmsten, PLoS One, 2012, 7, e44702 CAS.
- F. G. Wu, Z. W. Yu and G. Ji, Langmuir, 2011, 27, 2349–2356 CrossRef CAS PubMed.
- P. Saveyn, J. Cocquyt, M. Gradzielski and P. Van der Meeren, Colloids Surf., A, 2008, 319, 62–70 CrossRef CAS.
- B. Ma, S. Zhang, H. Jiang, B. Zhao and H. Lv, J. Controlled Release, 2007, 123, 184–194 CrossRef CAS PubMed.
- B. Angelov, A. Angelova, V. M. Garamus, M. Drechsler, R. Willumeit, R. Mutafchieva, P. Stepanek and S. Lesieur, Langmuir, 2012, 28, 16647–16655 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c6ra08125h |
| This journal is © The Royal Society of Chemistry 2016 |