
Assessment of the interacting mechanism between Candida rugosa lipases and hydroxyapatite and identification of the hydroxyapatite-binding sequence through proteomics and molecular modelling†
Abstract
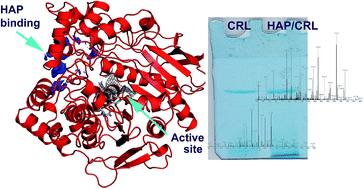
Hydroxyapatite (HAP), a calcium-phosphate bioactive ceramic, is actively employed in medical and separation sciences. Although different classes of biomacromolecules interact with this material, interactions with proteins are the most important, since they directly affect the biocompatibility of the carrier and it's industrial application. In the presented work, we thoroughly investigate and elucidate the interaction mechanism between Candida rugosa lipase (CRL) upon it's immobilization on HAP, since this immobilized enzyme showed advanced catalytic properties in previous studies. Applying elution and protein modification strategies we concluded that Ca-chelation of HAP's C-site and CRL's –COOH groups is the most probable interacting mechanism. A proteomics approach revealed that this chelation is conserved throughout all CRL isoforms, while results of molecular modelling led us to propose the involvement of a specific region of the protein surface and side chains in interactions with HAP.
 Please wait while we load your content...
Please wait while we load your content...

