
Multi-target detection of oxidative stress biomarkers in quercetin and myricetin treated human red blood cells
Abstract
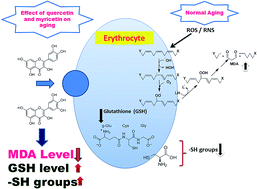
Quercetin and myricetin are important dietary flavonoids with potential health benefits and interfere with reactive oxygen species metabolism. The objective of this study was multi-target spectroscopic analysis of oxidative stress biomarkers (malondialdehyde (MDA), glutathione (GSH) and sulfhydryl (–SH) groups) in quercetin and myricetin treated red blood cells (RBCs) during human aging. The study was carried out on clinically relevant blood samples obtained from 105 healthy subjects between the ages of 18–82 years. The subjects were divided into three age groups, young (18–35 years), middle (36–60 years) and old (>60 years). Oxidative stress was induced in vitro by incubating RBCs with 10−5 M tert-butyl hydroperoxide (t-BHP). The effects of flavonoids were evaluated by detecting MDA, GSH and –SH groups by co-incubating the RBCs in the presence of flavonoid (10−8 M to 10−5 M final concentration) and t-BHP. The GSH/GSSG ratios were estimated to demonstrate the antioxidant power of the RBCs. The results showed elevated MDA levels (p < 0.001) after incubation with t-BHP as compared to a control. The GSH and –SH groups significantly (p < 0.001) decreased when incubated with t-BHP. In vitro administration of both flavonoids significantly attenuated the deleterious effect of oxidative stress in erythrocytes from all age groups. We showed a significant (p < 0.0001) negative correlation (r = −0.8334) between GSH/GSSG during human aging. We believe that these findings are novel and will help in the fast screening of new chemical molecules which may help against oxidative stress in RBCs, and thereby have tremendous scope in medical diagnostics and therapeutics.
 Please wait while we load your content...
Please wait while we load your content...

