
Magnetite nanoparticles enable a rapid conversion of volatile fatty acids to methane†
Abstract
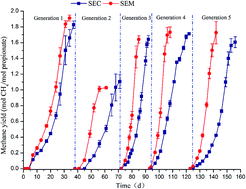
The objective of this work was to evaluate whether paddy soil enrichments obtained in the presence of magnetite and propionate (named SEM) can accelerate methane production from mixed VFAs. To do this, the SEM were firstly obtained via a semi-continuous subculture. The results showed that the addition of magnetite not only promoted the maximum production rate, but also reduced the methane production time in a dose-independent manner. The observation that Fe(III)-reducing Thauera was the dominant bacteria, combined with the observation that the methane production and Fe(II) formation proceeded in parallel, suggested that magnetite acted as an electron acceptor, may accelerate rapid oxidation of propionate and facilitated the establishment of a syntrophic interaction of Thauera with Methanobacterium. The SEM gave a methane yield and production rate that were on average 2.7-fold and 3.4-fold higher, respectively than those in paddy soil enrichments obtained in the absence of magnetite in conversion of the mixed VFAs to methane. The findings indicated the remarkable application potential of the SEM for improving methane production performance of the mixed VFAs.
 Please wait while we load your content...
Please wait while we load your content...

