
Relationships of Echium plantagineum L. bee pollen, dietary flavonoids and their colonic metabolites with cytochrome P450 enzymes and oxidative stress
C. Sousa,
P. B. Andrade and
P. Valentão*
REQUIMTE/LAQV, Laboratório de Farmacognosia, Departamento de Química, Faculdade de Farmácia, Universidade do Porto, Rua de Jorge Viterbo Ferreira, no. 228, 4050-313 Porto, Portugal. E-mail: valentao@ff.up.pt; Fax: +351 226093390; Tel: +351 220428653
First published on 6th January 2016
Abstract
Echium plantagineum L. bee pollen is a dietary source of flavonoids, which can play a protective role in the gastrointestinal tract by modulating cytochrome P450 (CYP) biotransformation enzymes and by opposing oxidative stress. Two fractions of E. plantagineum bee pollen phenolic extract, enriched in either flavonols (fraction I) or anthocyanins (fraction II), a whole extract and a prototype flavonoid of each class with a colonic metabolite were evaluated in a cell-free system for their potential to inhibit the activity of CYP enzymes expressed by human enterocytes. The superoxide scavenging potential of the individual compounds, along with their ability to protect Caco-2 cells subjected to oxidative stress induced by t-BHP, was also evaluated. CYP1A1 and CYP3A4 activity was efficiently inhibited by the three extracts. Fraction I, nearly devoid of anthocyanins, was almost as active as the whole extract, while fraction II, enriched in anthocyanins but with lower amounts of flavonols heterosides, was the less active. Among prototype phenolics, kaempferol was the best CYP1A1 inhibitor and cyanidin was the most active against CYP3A4, while their colonic metabolites p-hydroxyphenylacetic acid and protocatechuic acid were inactive. In accordance with this result, cyanidin and, to a less extent, kaempferol protected Caco-2 cells against t-BHP, which is bioactivated by CYP enzymes. A direct antioxidant effect could also contribute to the afforded protection, since both compounds were able to scavenge superoxide radicals in a cell-free system, cyanidin being the most active.
Introduction
Honey-bee collected pollen has been heralded as “nature's perfect food” and represents a potential source of energy and protein,1 and of bioactive phytochemicals for human consumption.2,3 It consists on plant pollens collected by worker bees, combined with plant nectar and bee saliva and packed into pellets by the bees.4 Bee pollen often contains mixtures of pollens from several types of plants, but it can also be almost mono-floral. In particular, Echium plantagineum L. bee pollen is of very high quality, presenting levels of protein above 30% and considerable amounts of flavonoids, namely flavonols and anthocyanins.3,5Flavonoids are one of the most widespread groups of secondary metabolites in plants. These compounds can exert an array of biological actions in consumers, either in their native form or after degradation/biotransformation. For this reason, the metabolic fate of flavonoids after ingestion is considered to be especially important. In particular, metabolites produced by intestinal microflora can significantly contribute to the bioactivities of flavonoids in the gastrointestinal tract.6 Indeed, the intestinal microorganisms can lead to the cleavage of glycosidic bonds in heterosides, ring fission of flavonoid backbone and the reduction of double bonds in the side chain of some intermediates.7 These reactions lead to the formation of phenylacetic acids derivatives as one of the metabolites obtained from ingested flavonols. Regarding kaempferol, the major flavonol aglycone present in E. plantagineum bee pollen extracts, the formation of p-hydroxyphenylacetic acid was observed both in in vivo assays and with cultures of intestinal microorganisms.3,7
In what concerns to anthocyanins, it appears that their spontaneous degradation at physiologic pH rather than the biotransformation by microorganisms contributes for the formation of benzoic acids derivatives, by ring fission of the anthocyanidin backbone.8 It has been reported that protocatechuic acid is the major metabolite found in rat and human plasma after cyanidin glucosides ingestion.9,10
In a previous work E. plantagineum bee pollen extracts enriched either in flavonol heterosides or anthocyanins, as well as a whole extract, were tested for their ability to protect human epithelial colorectal adenocarcinoma (Caco-2) cells against tert-butyl hydroperoxide (t-BHP)-induced oxidative stress.3 The dual effects exerted by the extracts were partly explained by the interaction of phenolic compounds with the cellular mechanisms involved in the detoxification/bioactivation of t-BHP.3 This short-chain analogue of lipid hydroperoxide can be detoxified by glutathione peroxidase (GPx) at expenses of reduced glutathione (GSH). Nevertheless, as commonly seen with many xenobiotics, t-BHP is also bioactivated by cytochrome P450 (CYP) phase I enzymes.11 Phase I and phase II enzymes are also modulated by phenolic compounds.12,13
CYP enzymes are a family of haem-containing monooxygenases that participate in the biosynthesis of hormones, second messengers, and other natural endobiotics. They also dominate xenobiotic detoxification and human drug metabolism, mainly by catalysing the first step of the detoxification of lipophilic substances.14,15 However, in some cases foreign compounds are bioactivated to products with much greater cytotoxicity, mutagenicity, or carcinogenicity.14 Human enterocytes, as well as the Caco-2 cell line, mainly express two CYP isoforms: CYP1A1 (highly inducible, but whose basal expression in the intestine and Caco-2 cells is very low) and CYP3A4 (responsible for most oxidations performed by enterocytes).12
This study aims to investigate the effect of E. plantagineum bee pollen extracts, as well as selected polyphenols and colonic metabolites, in human CYP1A1 and CYP3A4 activities. For this, two fractions of previously characterized E. plantagineum bee pollen phenolic extract, enriched in either flavonols (fraction I) or anthocyanins (fraction II) and a whole extract were evaluated in a cell-free system. Since most flavonoids present in E. plantagineum bee pollen must be at least partly deglycosylated in the lumen before being absorbed,16 the aglycones kaempferol and cyanidin and their colonic metabolites p-hydroxyphenylacetic acid and protocatechuic acid were assayed, in order to establish a possible correlation between the chemical composition of the extracts and the observed activity. The effects of these compounds in Caco-2 cells insulted with t-BHP, an oxidant that is bioactivated by CYP enzymes, was also evaluated. Furthermore as these compounds are claimed to have a direct antioxidant effect, the superoxide scavenging potential was included, in order to clarify the mechanisms of protection.
Experimental
Standards and reagents
Sodium pyruvate, (4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), β-nicotinamide adenine dinucleotide reduced form (NADH), dimethyl sulfoxide (DMSO), t-BHP, phenazine methosulfate (PMS), nitroblue tetrazolium chloride (NBT), superoxide dismutase (SOD) bovine (EC 1.15.1.1), Triton X-100, 2-amino-2-(hydroxymethyl)-1,3-propanediol (Trizma base), potassium phosphate monobasic, magnesium chloride, 7-ethoxyresorufin (7ER), resorufin, α-naphthoflavone, fluorescein, glucose-6-phosphate dehydrogenase from baker's yeast (S. cerevisiae) (EC 1.1.1.49), glucose-6-phosphate, β-nicotinamide adenine dinucleotide phosphate hydrate (NADP+), cyanidin chloride (Ph. Eur. Reference Standard), protocatechuic acid and p-hydroxyphenylacetic acid were from Sigma-Aldrich (St. Louis, MO, USA). Kaempferol was obtained from Extrasynthese (Genay, France). Methanol and acetonitrile were purchased from Merck (Darmstadt, Germany). Sodium hydroxide, ethyl acetate and hydrochloric acid were from BDH Prolabo (Dublin, Ireland). Recombinant human cytochromes P450 (Supersomes™) CYP1A1 and CYP3A4 (+P450 reductase + cytochrome b5), dibenzylfluorescein (DBF) and ketoconazole were from Corning® (Woburn, MA). Dulbecco's Modified Eagle Medium (DMEM) + GlutaMAX™-I, heat inactivated foetal bovine serum (FBS), nonessential amino acids (NEAA), Pen Strep solution (penicillin 5000 units per ml and streptomycin 5000 mg ml−1), fungizone (250 μg ml−1 amphotericin B), human transferrin (4 mg ml−1) and trypsin–EDTA were purchased from Gibco, Invitrogen™ (Invitrogen, Paisley, UK). Human epithelial colorectal adenocarcinoma (Caco-2) cell line was acquired from American Type Culture Collection (ATCC) (LGC Standards S.L.U., Barcelona, Spain). Water was deionized using a Milli-Q water purification system (Millipore, Bedford, MA, USA).Compounds and extracts
E. plantagineum bee pollen, provided by beekeepers in the Spanish Extremadura region, was extracted as previously described.3 Briefly, aliquots of 1 g of bee pollen was sonicated for 1 h with 10 ml of methanol containing 0.5% HCl (v/v) and centrifuged at 4000 rpm for 10 min. The supernatant was decanted, the residue was re-extracted by thoroughly mixing with 10 ml of acidified methanol for 2 min, and centrifuged. The supernatants were pooled together, the solvent was evaporated under reduced pressure at 30 °C and the residue was dissolved in 10 ml of water. A solid-phase extraction (SPE) procedure was performed using C18 columns (45 μm particle size, 60 Å pore size; 10 g sorbent mass/70 ml reservoir volume) preconditioned with 20 ml of ethyl acetate, 20 ml of methanol, and 20 ml of water. The loaded cartridge was washed with 10 ml of water and the compounds were eluted with 100 ml ethyl acetate (fraction I), followed by 40 ml of methanol (fraction II). The whole extract was obtained by gathering ethyl acetate and methanol eluates.20 mM stock solutions of kaempferol, protocatechuic acid and p-hydroxyphenylacetic were prepared in methanol. Due to the instability of anthocyanins at physiological pH, cyanidin stock solutions (400× the higher concentration tested) were prepared in methanol with pH adjusted to 2 with HCl, in order to prevent degradation. Before tests, the stock solution was diluted with phosphate buffer (CYP activity assays) or culture medium (Caco-2 cells and superoxide scavenging assays) and assayed at physiologic pH. Final concentration of methanol never exceeded 1%.
Cytochrome P450 activity assays
Fluorescence-based assays were used to assess the inhibition of human recombinant CYP isoenzymes activity by the extracts and compounds.17,18 The substrates were 7ER for CYP1A1 and DBF for CYP3A4.18 Reactions were carried out at 37 °C in black/transparent 96-well plates; final concentrations in 200 μl of 50 mM phosphate buffer pH 7.4 were as follows: 1.5 mM MgCl2, 0.2 U ml−1 of glucose-6-phosphate dehydrogenase, 1.65 mM glucose-6-phosphate, 0.65 mM NADP+, 1 μM substrate (7ER or DBF) and sample or vehicle (blank). Reactions were initiated with the addition of the enzyme (0.5 pmol per well) and were maintained for 10 min (CYP1A1) or 60 min (CYP3A4). CYP1A1 reactions were stopped by addition of 75 μl of a acetonitrile![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :0.5 M Trizma base solution (4:1). Resorufin resulting from the O-de-ethylation of 7ER was measured in a microplate spectrofluorimeter (Synergy HT, Bio-Tek Instruments Winooski, USA, operated by Gen5 2.01 software) at the excitation wavelength of 530 nm and emission wavelength of 590 nm. The O-de-benzylation of DBF to fluorescein by CYP3A4 was stopped by adding 75 μl of NaOH 2 M and the plates were further incubated for 20 min at room temperature. Fluorescein was measured at the excitation wavelength of 485 nm and emission wavelength of 528 nm. α-Naphthoflavone dissolved in methanol:ethyl acetate (2:3) was used as positive control for CYP1A1, while ketoconazole dissolved in acetonitrile was the positive control for CYP3A4. The final concentration of solvents never exceeded 0.25%. At least, four independent assays were performed in duplicate.
:0.5 M Trizma base solution (4:1). Resorufin resulting from the O-de-ethylation of 7ER was measured in a microplate spectrofluorimeter (Synergy HT, Bio-Tek Instruments Winooski, USA, operated by Gen5 2.01 software) at the excitation wavelength of 530 nm and emission wavelength of 590 nm. The O-de-benzylation of DBF to fluorescein by CYP3A4 was stopped by adding 75 μl of NaOH 2 M and the plates were further incubated for 20 min at room temperature. Fluorescein was measured at the excitation wavelength of 485 nm and emission wavelength of 528 nm. α-Naphthoflavone dissolved in methanol:ethyl acetate (2:3) was used as positive control for CYP1A1, while ketoconazole dissolved in acetonitrile was the positive control for CYP3A4. The final concentration of solvents never exceeded 0.25%. At least, four independent assays were performed in duplicate.
The turnover numbers were calculated based on a standard curve prepared using resorufin (CYP1A1) or fluorescein (CYP3A4). In order to check for interferences, the calibration curves were built under the assays conditions, except for the presence of the enzymes and substrates that where substituted by the CYP reaction product. Due to interference of cyanidin in the fluorescence signal of resorufin and fluorescein, a calibration curve was built in the presence of each cyanidin concentration for interpolation of the respective turnover number.
Caco-2 culture conditions and treatments
Caco-2 cells were cultured according to a previously published method.3 Briefly, cells were grown in DMEM + GlutaMAX™-I supplemented with 10% calf serum (FCS) heat-inactivated, 1% NEAA, 1% fungizone and 6 μg ml−1 transferrin, 1% penicillin and 1% streptomycin, at 37 °C in 5% CO2. For cells treatments the culture medium was prepared without FCS and cells were exposed to the compounds for 24 h after reaching confluence. In order to evaluate the antioxidant potential, cells were further exposed to 150 μM of t-BHP for 6 h in fresh medium (without the test compounds).Viability assays
The viability of cells was assessed according to previously published methods.3 The mitochondria-dependent reduction of MTT (0.5 mg ml−1) to formazan during 30 min was quantified at 560 nm in a microplate reader (Multiskan ASCENT) and the percentage of the cytosolic enzyme lactate dehydrogenase (LDH) released into culture medium was measured by following the rate of oxidation of NADH at 340 nm during the conversion of pyruvate to lactate. The total amount of LDH was measured after treating the cells for 30 min with Triton X-100 1%. Viability is expressed as percentage of control (without t-BHP). Five independent experiments were performed in triplicate.Superoxide scavenging
The scavenging of PMS–NADH-generated superoxide was determined in biological relevant conditions, using DMEM as a solvent (pH 7.4; physiological salt and glucose concentrations), by measuring the reduction of NBT at 560 nm.3 Bovine SOD was assayed as positive control. Five independent assays were performed in triplicate. The results are expressed as % of control.Statistical analysis
Statistical analysis was performed using GraphPad Prism 6 Software (San Diego, CA, USA). Data are expressed as the mean ± standard error of the mean (SEM) of, at least, four independent experiments performed, at least, in duplicate. One Way ANOVA and Dunnets's test as post hoc test, were used to determine the statistical significance in comparison to control. One way ANOVA and Tukey's multiple comparisons test or unpaired Mann–Whitney test (only two groups) were used to compare IC50 values. Values of p ≤ 0.05 were considered significant.Results and discussion
Cytochrome P450 inhibition
The CYP family encodes numerous enzymes that are remarkable in the variety of chemical reactions catalysed and are particularly prone to inhibition because of their broad substrate specificity.14 Two important isoforms expressed by enterocytes are CYP1A1 and CYP3A4, suggesting a strategy for controlling the entrance of xenobiotics into the body.19 Since these enzymes can be modulated by dietary flavonoids,12 in this work the inhibition of CYP1A1 and CYP3A4 by E. plantagineum bee pollen extracts was evaluated. Human recombinant enzymes were used due to species specificity in modulating biotransformation enzymes.20 All extracts contained kaempferol derivatives (9.58 mmol kg−1 in fraction I, 1.63 mmol kg−1 in fraction II and 11.06 mmol kg−1 in whole extract) and anthocyanins (0.50 μmol kg−1 in fraction I, 55.67 μmol kg−1 in fraction II and 65.01 μmol kg−1 in whole extract), as previously determined by HPLC-DAD.3 All these flavonoids have glycosylated structure, which is water-soluble and chemically stable. In the human body the bioavailability of glycosides can be either higher or lower than that of the aglycones. For instance, flavonols conjugated with glucose can be better absorbed in the small intestine than the aglycones, whereas polyphenols linked to rhamnose must be deconjugated by the colon microflora before absorption.16 In this study, only free kaempferol and cyanidin were tested, as most flavonols and anthocyanins present in E. plantagineum bee pollen have rhamnose in their glycosidic moiety and must probably be, at least partly, hydrolysed in the lumen before absorption by enterocytes.3,16 p-Hydroxyphenylacetic acid, one of the main compounds resulting from kaempferol metabolism by colonic microflora,7 and protocatechuic acid, resulting from the metabolization of cyanidin,9 were also studied.The potential to inhibit CYP1A1 was evaluated using the substrate 7-ethoxyresorufin, which is known to be selective towards CYP1A1/2 isoforms.21 The amount of resorufin formed was calculated by interpolation in the calibration curve (Fig. 1A). In the assay conditions resorufin reached 91.973 ± 1.101 nM after 10 min incubation with 2.5 nM CYP1A1 and 1 μM 7ER (turnover of 3.679 ± 0.044 nmol resorufin−1 nmol CYP1A1−1 min−1). Under the assay conditions α-naphthoflavone (CYP1A1/2 selective inhibitor) showed half-maximal inhibition (IC50) of 0.521 ± 0.059 μM (Fig. 1B). Considering E. plantagineum bee pollen, all the extracts were able to significantly inhibit CYP1A1 activity (Fig. 1C, Table 1). The whole extract was the most active with an IC50 of 0.335 ± 0.053 mg ml−1, followed by fraction I (IC50 of 0.651 ± 0.134 mg ml−1), and fraction II (IC50 of 5.853 ± 0.314 mg ml−1). Statistical differences in the IC50 were found between fraction II and the other extracts (p < 0.0001, n = 5).
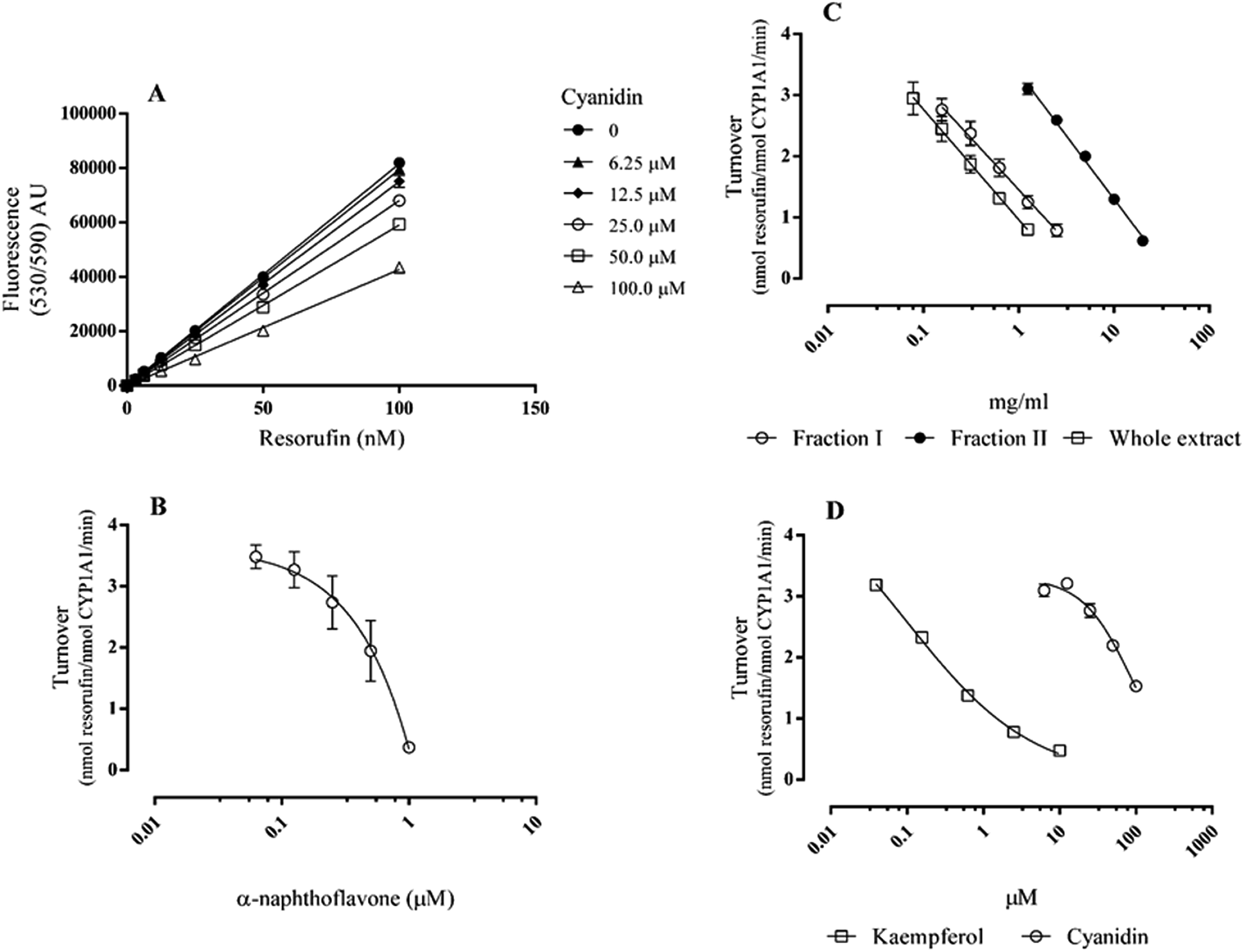 | ||
| Fig. 1 CYP 1A1 inhibition by Echium plantagineum L. bee pollen and selected phenolic compounds: resorufin calibration curve alone and with variable concentrations of cyanidin (A); dose–response curve of the positive control (B), E. plantagineum bee pollen extracts (C) and flavonoids (D). Results are presented as mean ± standard error of the mean of, at least, three independent experiments performed in duplicate. | ||
| Fraction I | Fraction II | Whole extract | |
|---|---|---|---|
| a Different superscript letters in the same row show statistically significant differences (p < 0.001 or higher).b Calculated from data on Sousa et al., 2015.3 | |||
| CYP 1A1 | |||
| IC50 (mg ml−1) | 0.651 ± 0.134a | 5.853 ± 0.314b | 0.335 ± 0.053a |
| Anthocyanins (nM)b | 0.33 | 325.86 | 21.78 |
| Flavonols (nM)b | 570 | 140 | 300 |
|
|||
| CYP3A4 | |||
| IC50 (mg ml−1) | 0.570 ± 0.089a | 2.159 ± 0.295b | 0.374 ± 0.043a |
| Anthocyanins (nM)b | 0.29 | 120.2 | 24.3 |
| Flavonols (nM)b | 500 | 50 | 340 |
The CYP inhibitory potential of the selected phenolic compounds was assayed exactly under the same conditions as the extracts for comparison purposes. Among them only kaempferol and cyanidin were able to significantly inhibit CYP1A1 activity (Fig. 1D), while their metabolites p-hydroxyphenylacetic acid and protocatechuic acid, which are not polycyclic, did not exert a significant activity until the concentration of 200 μM (data not shown). Kaempferol, with an IC50 of 0.294 ± 0.019 μM was more active than cyanidin (IC50 of 86.599 ± 8.671 μM). Although kaempferol has been reported as a good CYP1A1 inhibitor in other models, when compared with α-naphthoflavone, its inhibitory potential is generally weaker.22 In this work, the IC50 of kaempferol was significantly lower (nearly half) than the one of α-naphthoflavone (p < 0.01, n = 5). The inhibitory potential of CYP1A1 by kaempferol heterosides, the type of flavonols present in E. plantagimeum bee pollen extracts3 has been reported to be statistically significant, although much lower than that of kaempferol.21 Nevertheless all extracts were able to inhibit CYP1A1 activity (Table 1).
Cyanidin, which does not display autofluorescence at the wavelengths and concentrations used, significantly and dose-dependently quenched the fluorescence of resorufin, probably due to its high absorption at these wavelengths (Fig. 1A).23 So, in order to compensate for cyanidin interference, the amount of resorufin formed was calculated by interpolation in the calibration curve obtained with the respective cyanidin concentration (Fig. 1A). An R square value above 0.99 was obtained.
Considering CYP3A4 inhibition the substrate DBF was chosen, and the amount of fluorescein formed was quantified by interpolation in a calibration curve (Fig. 2A). The reaction performed with 2.5 nM CYP3A4 and 1 μM DBF was followed from 10 to 120 min and a reaction time of 60 min, within the linear range of metabolite formation, was selected for all the subsequent assays (data not shown). Under these conditions, the amount of fluorescein formed was 151.97 ± 5.38 nM (turnover of 1.014 ± 0.036 nmol fluorescein−1 nmol CYP3A4−1 min−1).
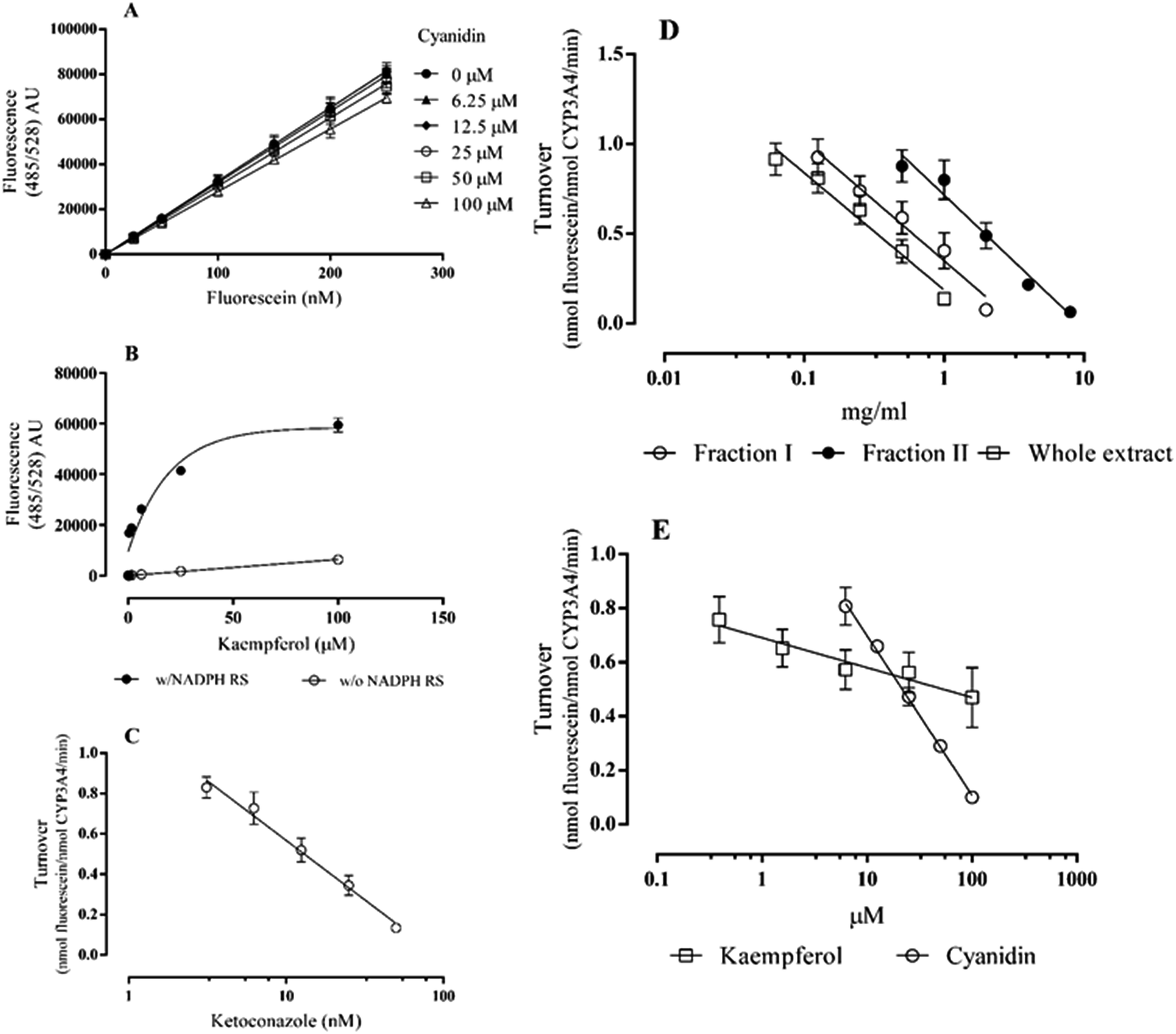 | ||
| Fig. 2 CYP 3A4 inhibition by Echium plantagineum L. bee pollen and selected phenolic compounds: fluorescein calibration curve alone and with variable concentrations of cyanidin (A); kaempferol autofluorescence (B); dose–response curve of the positive control (C), E. plantagineum bee pollen extracts (D) and flavonoids (E). Results are presented as mean ± standard error of the mean of, at least, three independent experiments performed in duplicate (w/NADPH RS: with NADPH regenerating system; w/o NADPH RS: without NADPH regenerating system). | ||
Extracts and compounds were checked for autofluorescence under the assay conditions. Kaempferol was found to be fluorescent: after 60 min incubation in the presence of NADPH regenerating system and in the absence of DBF (either with or without CYP3A4), the fluorescence increased significantly more than with kaempferol alone (Fig. 2B). This behaviour was only observed with kaempferol and the signal obtained in the reaction with DBF and CYP3A4 was corrected by subtracting the autofluorescence signal. Ketoconazole, a CYP3A4 selective inhibitor, strongly inhibited this CYP isoform with an IC50 of 12.50 ± 1.61 nM (Fig. 2C). Extracts of E. plantagineum bee pollen were able to inhibit CYP3A4 in this in vitro assay. Again, the whole extract (IC50 of 0.374 ± 0.043 mg ml−1) was more active than fraction I (IC50 of 0.570 ± 0.089 mg ml−1) (Fig. 2D, Table 1). Although fraction II (IC50 of 2.159 ± 0.295 mg ml−1) is significantly less active than both fraction I and the whole extract (p < 0.001), the differences in the inhibitory potential of this fraction towards CYP3A4 are smaller than the ones observed with the CYP1A1 isoform.
Among the tested compounds, only kaempferol and cyanidin were able to inhibit CYP3A4 activity, the last being the most active (Fig. 2E): the IC50 found for cyanidin was 26.80 ± 1.39 μM. It should be noted that the IC50 value found for ketoconazole is more than 1000× lower than that of cyanidin. Kaempferol reached 44.3 ± 3.9% inhibition at 100 μM. Higher concentrations of kaempferol were not tested due to its high autofluorescence (Fig. 2B).
The results showed that the whole extract was the most effective in inhibiting both enzymes, but fraction I, nearly devoid of anthocyanins,3 was almost as active (Fig. 1C and 2D, Table 1). Fraction II, containing almost the same amount of anthocyanins of the whole extract, but with lower amounts of flavonols heterosides than the other two extracts,3 was the less active, especially towards CYP1A1.
Concerning the selected phenols, only the parent compounds showed activity, kaempferol being more efficient in inhibiting CYP1A1 and cyanidin more active against CYP3A4 (Fig. 1D and 2E). These results are in accordance with the ones obtained with the different extracts of E. plantagineum bee pollen. Although the levels of anthocyanins in all extracts are low,3 their effect is evidenced in the CYP3A4 inhibition by fraction II: this extract inhibited more efficiently this isoform than CYP1A1. The lower ability of anthocyanidins to inhibit CYP1A1 than CYP3A4 can be inferred from previous works using human liver microsomes: the activity of CYP3A4 could be almost completely inhibited by some anthocyanidins, with a significant inhibition by cyanidin itself, although slightly lower than 50% in this system.24 Sugar-conjugated anthocyanidins were weaker CYP3A4 inhibitors than the respective aglycones, but, even so, cyanidin-3-O-rahmnoside reached 25% CYP3A4 inhibition at 100 μM.24 In comparison, the inhibition of CYP1A1 by anthocyanidins and anthocyanins was always lower than 50%, and lower than 20% for cyanidin, petunidin, malvidin and peonidin in a similar system.25,26
Previous studies have shown that CYP3A4 is more resistant to flavonol inhibition than CYP1A1 (for instance, galangin has an IC50 of 2.3 μM for CYP3A4, compared to 0.073 μM for CYP1A1)22 which is also in accordance with the results obtained in this work with kaempferol (Fig. 1D and 2E).
Besides their ability to inhibit CYP enzymes, flavonoids can be a substrate for these enzymes.27 However, as commonly seen with other compounds, biotransformation of flavonoids by CYP enzymes is not considered a major clearance pathway when phase II metabolic pathway is active.28 Flavonoids can also be CYP inducers, as many other substrates that enhance xenobiotic detoxification during prolonged periods of body's exposition.15 The induction of CYP enzymes involves receptor-like xeno-sensors.20 The aryl hydrocarbon receptor (AhR) is clearly involved in the transcriptional activation of CYP1A1 gene,19 while the expression of CYP3A4 is induced by compounds that activate the pregnenolone X receptor (PXR), a nuclear receptor that binds to a response element in CYP3A4 promotor.29 Interaction of kaempferol with AhR and PXR is described as being rather weak.30 Similarly, cyanidin was described as a weak AhR agonist25 and it did not induce the expression of CYP3A4 mRNA or protein.24 Interestingly, moderate induction of CYP3A4 by plant extracts beverages in a human intestinal cell line has already been observed, CYP1A1 induction being less frequent.31 CYP1A1 is known to be a major contributor to the bioactivation of environmental toxins and pollutants, but a combined treatment with kaempferol and benzo[a]pyrene (B[a]P) restricted the CYP1A1 activity increase induced by B[a]P in Caco-2 cells.32
Superoxide scavenging
In addition to the substrate oxidation, cytochrome P450 enzymes can also produce reactive oxygen species (ROS) as side products, including hydrogen peroxide, superoxide and hydroxyl radicals.33 So, the scavenging of oxidant species can be an important mechanism of cellular protection linked to biotransformation reaction. The superoxide scavenging effect of phenolics was evaluated (Fig. 3A), as this radical is involved in the formation of not only more powerful oxygen reactive species, but also in the formation of nitrogen reactive species, upon its collision with nitric oxide radical, which is especially important under some pathological conditions in which nitric oxide synthases are stimulated.34 With the exception of p-hydroxyphenylacetic acid, all compounds scavenged superoxide in a dose-dependent manner. The concentrations causing 50% inhibition of formazan formation induced by NADH–PMS–O2 generated superoxide (EC50) were 107.8 ± 6.9 μM for cyanidin, 694.1 ± 32.1 μM for kaempferol and 839.8 ± 88.1 μM for protocatechuic acid. No significant differences were seen between kaempferol and protocatechuic acid. Under the same conditions, recombinant bovine SOD displayed an EC50 of 7.2 ± 0.4 nM (Fig. 3B). In the human body superoxide inactivation is probably exerted mainly by SOD,35 which has been reported to reach a level of 7.6 ± 0.1 μM SOD on the erythrocytes of healthy volunteers.36 Nevertheless, it has previously been shown, using different in vitro models, that kaempferol,35 cyanidin37 and protocatechuic acid38 are able to scavenge superoxide radical. Significantly, the ratio between the EC50 of kaempferol (12000 μM) and SOD (170 nM) was found to be considerably high,35 in accordance with the results obtained in this work. Kaempferol is a poor superoxide scavenger, in comparison with other flavonoids with a catechol group in the B.35 This fact can explain why the scavenging potential of protocatechuic acid, a phenolic acid with a catechol moiety, is not significantly different from that of kaempferol, as this work demonstrates. As expected, p-hydroxyphenylacetic acid, with only one phenolic hydroxyl group in its structure, has no superoxide scavenging activity.
 | ||
| Fig. 3 Dose–response curve of superoxide scavenging by the test compounds (A) and the positive control (B). Results are presented as mean ± standard error of the mean of five independent experiments performed in triplicate. | ||
The superoxide scavenging of E. plantagineum bee pollen extracts previously reported was inconsistent.3 Fraction I displayed ability to scavenge superoxide at all concentrations tested, while the extracts containing anthocyanins were only slightly antioxidant at some of the tested concentrations. However, it must be noted that in the superoxide scavenging assay with the extracts previously reported,3 the highest amounts of flavonols and anthocyanins tested were 221.2 μM and 1.3 μM, respectively (corresponding to whole extract at 20 mg ml−1), which are much lower than the EC50 of kaempferol, cyanidin or its metabolite protocatechuic acid found herein. This can partly explain the low ability of the extracts to act as efficient antioxidants in this model.
Cellular viability
It is known that CYP enzymes are involved in the bioactivation of t-BHP.11 For this reason the effect of the selected phenolic compounds against t-BHP induced toxicity was evaluated in human intestinal Caco-2 cells using a model previously applied to E. plantagineum bee pollen extracts.3 The effects of each compound on cellular viability were assessed in the range of 0.625 to 10 μM for cyanidin, and 12.5 to 200 μM for kaempferol, protocatechuic acid and p-hydroxyphenylacetic acid, with no significant differences in LDH leakage and MTT reduction (data not shown).The exposure of Caco-2 cells to 150 μM t-BHP for 6 h decreased the viability to 82.9 ± 2.6% of control in LDH leakage assay (p < 0.0001, n = 5) and to 40.1 ± 6.4% in the MTT assay (p < 0.0001; n = 5). Pre-exposition to kaempferol protected Caco-2 cells from t-BHP-induced toxicity for concentrations higher than of 12.5 μM in the LDH assay; with kaempferol at 50 μM the viability corresponded to 97.6 ± 1.5% of unexposed cells (p < 0.0001, n = 5) (Fig. 4A). In the MTT assay a significant increase of cellular viability was only seen for concentrations higher than 25 μM (p < 0.0001, n = 5) (Fig. 4B). Cyanidin at concentrations higher than 1.25 μM provided significant protection in both assays (p < 0.001 or higher, n = 5) (Fig. 4).
 | ||
| Fig. 4 Viability of Caco-2 cells pre-exposed to test compounds for 24 h and further insulted with 150 μM t-BHP for 6 h (no extract co-incubation): LDH (A) and MTT (B) assays. Results are presented as mean ± standard error of the mean of five independent experiments performed in triplicate. *p < 0.05, ***p < 0.001, ****p < 0.0001 compared with control. | ||
In the MTT assay, both kaempferol and cyanidin at the higher concentrations tested increase the cellular viability above the control level (around 115% and 130%, respectively) (Fig. 4B). This behaviour was already observed in other studies using MTT assay (which provides information on mitochondrial function) to assess cell viability.2 It is known that flavonoids can interact with Nrf2, a key regulator of phase II detoxifying enzymes and this effect can contribute to a better cellular status.39
Considering the colonic metabolites, protocatechuic acid at 200 μM afforded some protection, as evaluated by the LDH assay (p < 0.05, n = 5) (Fig. 4A). This tendency to protect cell viability was also seen in the MTT assay (Fig. 4B). p-Hydroxyphenylacetic acid at the concentrations tested did not impart any protection to Caco-2 cells in both assays (Fig. 4).
The higher cellular protection afforded by cyanidin correlates with the antioxidant ability demonstrated by this compound in the superoxide scavenging assay (Fig. 3 and 4). However, protocatechuic acid, which was found to have an antioxidant potential similar to that of kaempferol in the superoxide scavenging assay, did not provide the same protection level of kaempferol in Caco-2 cells insulted with t-BHP. Since the insult with t-BHP was performed after removing the tested compounds from the culture medium, only those absorbed by the cells could exert a direct scavenging effect. Kaempferol, with only one hydroxyl group in the B ring, is considered a highly permeable flavonoid in Caco-2 cells, and a better absorption of kaempferol during the pre-exposition period can, at least partly, justify the differences in cellular protection between these two compounds.40 Interestingly, it has been reported that protocatechuic acid can be an excellent antioxidant in aqueous solution, but in non-polar environments it is only moderately protective.38 Despite the results of protocatechuic acid, plant extracts rich in esters of phenolic acids, like rosmarinic acid, can be highly protective in this cellular model and completely abolish the deleterious effects of t-BHP in Caco-2 cells viability.41
Besides the scavenging potential of the phenolic compounds, their effects on CYP enzymes can greatly contribute for the differences in cellular protection. As kaempferol and cyanidin can significantly inhibit CYP1A1 and CYP3A4, they can contribute in this way to decrease the generation of toxic radicals in exposed cells, decreasing cellular injury and death. Furthermore, since cyanidin revealed a higher ability to inhibit CYP3A4, which is the major CYP isoform constitutively expressed by enterocytes, the better protection afforded by cyanidin was expected.12
Comparing with the cellular effects of E. plantagineum bee pollen extracts in t-BHP insulted Caco-2 cells previously published,3 the flavonoid aglycones were more effective in protecting cells against oxidative stress. It should be noted that although the levels of flavonols in the extracts where in some cases higher than the amounts of kaempferol tested, the levels of anthocyanins in the extracts were well below the amount of cyanidin needed to afford some protection (Table 1). As it can be seen in Fig. 4, cyanidin started to be protective at 2.5 μM and the levels of anthocyanins tested never exceeded 1.3 μM (whole extract at 20 mg ml−1).3 Nevertheless, all extracts were able to significantly inhibit both CYP1A1 and CYP3A4 in the cell free assay, at concentrations lower than the ones used in the cellular assay, which was not always the case with the tested phenolic compounds. As only heterosides were quantified in E. plantagineum bee pollen extracts, and this kind of heterosides probably need to be hydrolysed by colonic microflora in order to be absorbed, the capacity to decrease the levels of oxidative stress in Caco-2 cells insulted with t-BHP could not be clearly seen with any extract. Furthermore, the concentrations in enterocytes depend on several processes, including uptake via passive diffusion or active transport, on one hand, and their affinity to various efflux pumps that can actively transport them back out, on the other, limiting the amount of compound present inside the cell after the pre-exposition period.28
Conclusions
Dietary sources of flavonoids are expected to modulate cytochrome P450 biotransformation enzymes, while affording protection against oxidative stress in the gastrointestinal tract, due to the action of the ingested compounds or their metabolites. In this study three E. plantagineum bee pollen extracts significantly inhibited CYP1A1 and CYP3A4 activity, the whole extract being more effective than flavonols or anthocyanins enriched fractions. Considering individual compounds, cyanidin inhibited more effectively CYP3A4 than kaempferol, while the opposite was seen for CYP1A1 inhibition, emphasizing the importance of the ingestion of whole complex mixtures for a wider range of effects. Colonic metabolites of cyanidin and kaempferol revealed to be inactive against these two CYP isoforms. Considering antioxidant activity, kaempferol and cyanidin were able to protect Caco-2 cells against oxidative stress induced by t-BHP. Since t-BHP is bioactivated by CYP enzymes, which were found to be effectively inhibited by cyanidin and kaempferol, it can be postulated that these compounds may decrease the levels of oxidative species originated from t-BHP. A direct antioxidant effect could also contribute for the afforded protection, since the best results in cells were obtained with cyanidin, which was also the best superoxide scavenger among the tested compounds. In what concerns to E. plantagineum bee pollen extracts, the potential to inhibit CYP enzymes observed in the cell free assays was not reflected in a clear cellular protection, maybe due to low permeability of flavonoid heterosides predominating in the extracts, which contain glycosidic chains with rhamnose. Since these compounds can be deglycosylated by intestinal microflora, a better protection by bee pollen compounds can be expected in in vivo conditions. Overall this work contributed to establish a connection between modulation of biotransformation enzymes and antioxidant activity in the beneficial effects afforded by phenolic compounds-rich products or individual compounds.Acknowledgements
This work received financial support from the European Union (FEDER funds through COMPETE) and National Funds (FCT, Fundação para a Ciência e Tecnologia) through project UID/QUI/50006/2013. To all financing sources the authors are greatly indebted.Notes and references
- R. R. Bell, E. J. Thornber, J. L. L. Seet, M. T. Groves, N. P. Ho and D. T. Bell, J. Nutr., 1983, 113, 2479 CAS.
- E. Moita, A. Gil-Izquierdo, C. Sousa, F. Ferreres, L. R. Silva, P. Valentão, R. Domínguez-Perles, N. Baenas and P. B. Andrade, PLoS One, 2013, 8, e59131 CAS.
- C. Sousa, E. Moita, P. Valentão, F. Fernandes, P. Monteiro and P. B. Andrade, J. Agric. Food Chem., 2015, 63, 2083 CrossRef CAS PubMed.
- H. F. Linskens and W. Jorde, Econ. Bot., 1997, 51, 78 CrossRef.
- A. M. G. Paramás, J. A. G. Bárez, C. C. Marcos, R. J. García-Villanova and J. S. Sánchez, Food Chem., 2006, 95, 148 CrossRef.
- C. Manach, G. Williamson, C. Morand, A. Scalbert and C. Rémésy, Am. J. Clin. Nutr., 2005, 81, 230S CAS.
- L. A. Griffiths and G. E. Smith, Biochem. J., 1972, 128, 901 CrossRef CAS.
- C. D. Kay, P. A. Kroon and A. Cassidy, Mol. Nutr. Food Res., 2009, 53, S92 Search PubMed.
- T. Tsuda, F. Horio and T. Osawa, FEBS Lett., 1999, 449, 179 CrossRef CAS PubMed.
- P. Vitaglione, G. Donnarumma, A. Napolitano, F. Galvano, A. Gallo, L. Scalfi and V. Fogliano, J. Nutr., 2007, 137, 2043 CAS.
- C. R. Myers, in Free radical toxicology, ed. K. B. Wallace, Taylor & Francis, Washington DC, 1997, ch. 3, pp. 30–31 Search PubMed.
- T. Sergent, I. Dupont, E. van der Heiden, M. L. Scippo, L. Pussemier, Y. Larondelle and Y. J. Schneider, Toxicol. Lett., 2009, 191, 216 CrossRef CAS PubMed.
- W. Jiang and M. Hu, RSC Adv., 2012, 2, 7948 RSC.
- T. D. Porter and M. J. Coon, J. Biol. Chem., 1991, 266, 13469 CAS.
- T. M. Willson and S. A. Kliewer, Nat. Rev. Drug Discovery, 2002, 1, 259 CrossRef CAS PubMed.
- C. Manach, A. Scalbert, C. Morand, C. Rémésy and L. Jiménez, Am. J. Clin. Nutr., 2004, 79, 727 CAS.
- C. L. Crespi, V. P. Miller and B. W. Penman, Anal. Biochem., 1997, 248, 188 CrossRef CAS PubMed.
- A. Ghosal, N. Hapangama, Y. Yuan, X. Lu, D. Horne, J. E. Patrick and S. Zbaida, Biopharm. Drug Dispos., 2003, 24, 375 CrossRef CAS PubMed.
- Q. Y. Zhang, D. Dunbar, A. Ostrowska, S. Zeisloft, J. Yang and L. S. Kaminsky, Drug Metab. Dispos., 1999, 27, 804 CAS.
- W. Xie, J. L. Barwick, M. Downes, B. Blumberg, C. M. Simon, M. C. Nelson, B. A. Neuschwander-Tetri, E. M. Bruntk, P. S. Guzelian and R. M. Evans, Nature, 2000, 406, 435 CrossRef CAS PubMed.
- L. Ribonnet, A. Callebaut, I. Nobels, M. L. Scippo, Y. J. Schneider, S. de Saeger, L. Pussemier and Y. Larondelle, Toxicol. Lett., 2011, 202, 193 CrossRef CAS PubMed.
- T. Shimada, K. Tanaka, S. Takenaka, N. Murayama, M. V. Martin, M. K. Foroozesh, H. Yamazaki, P. Guengerich and M. Komori, Chem. Res. Toxicol., 2010, 23, 1921 CrossRef CAS PubMed.
- M. J. Rebelo, C. Sousa, P. Valentão, R. Rego and P. B. Andrade, Food Chem., 2014, 163, 16 CrossRef CAS PubMed.
- A. Srovnalova, M. Svecarova, M. K. Zapletalova, P. Anzenbacher, P. Bachleda, E. Anzenbacherova and Z. Dvorak, J. Agric. Food Chem., 2014, 62, 789 CrossRef CAS PubMed.
- A. Kamenickova, E. Anzenbacherova, P. Pavek, A. A. Soshilov, M. S. Denison, P. Anzenbacher and Z. Dvorak, Toxicol. Lett., 2013, 218, 253 CrossRef CAS PubMed.
- A. Kamenickova, E. Anzenbacherova, P. Pavek, A. A. Soshilov, M. S. Denison, M. Zapletalova, P. Anzenbacher and Z. Dvorak, Toxicol. Lett., 2013, 221, 1 CrossRef CAS.
- V. M. Breinholt, E. A. Offord, C. Brouwer, S. E. Nielsen, K. Brøsen and T. Friedberg, Food Chem. Toxicol., 2002, 40, 609 CrossRef CAS PubMed.
- R. Mukkavilli, S. R. Gundala, C. Yang, S. Donthamsetty, G. Cantuaria, G. R. Jadhav, S. Vangala, M. D. Reid and R. Aneja, PLoS One, 2014, 9, e108386 Search PubMed.
- J. M. Lehmann, D. D. McKee, M. A. Watson, T. M. Willson, J. T. Moore and S. A. Kliewer, J. Clin. Invest., 1998, 102, 1016 CrossRef CAS PubMed.
- H. P. Ciolino, P. J. Daschner and G. C. Yeh, Biochem. J., 1999, 340, 715 CrossRef CAS PubMed.
- A. Kamenickova, R. Vrzal and Z. Dvorak, Food Chem., 2012, 131, 1201 CrossRef CAS.
- J. Niestroy, A. Barbara, K. Herbst, S. Rode, M. van Liempt and P. H. Roos, Toxicol. in Vitro, 2011, 25, 671 CrossRef CAS PubMed.
- H. Kuthan, H. Tsuji, H. Graf, V. Ullrich, J. Werringloer and R. W. Estabrook, FEBS Lett., 1978, 91, 343 CrossRef CAS PubMed.
- R. B. Mikkelsen and P. Wardman, Oncogene, 2003, 22, 5734 CrossRef CAS PubMed.
- D. Taubert, T. Breitenbach, A. Lazar, P. Censarek, S. Harlfinger, R. Berkels, W. Klaus and R. Roesen, Free Radicals Biol. Med., 2003, 35, 1599 CrossRef CAS PubMed.
- Y. Sun, L. W. Oberley and Y. Li, Clin. Chem., 1988, 34, 497 CAS.
- H. Yamasaki, H. Uefuji and Y. Sakihama, Arch. Biochem. Biophys., 1996, 332, 183 CrossRef CAS PubMed.
- A. Galano and A. Pérez-González, Theor. Chem. Acc., 2012, 131, 1 Search PubMed.
- A. Gopalakrishnan and A. N. T. Kong, Food Chem. Toxicol., 2008, 46, 1257 CrossRef CAS PubMed.
- X.-J. Tian, X.-W. Yang, X. Yang and K. Wang, Int. J. Pharm., 2009, 367, 58 CrossRef CAS PubMed.
- F. Ferreres, J. Bernardo, P. B. Andrade, C. Sousa, A. Gil-Izquierdo and P. Valentão, RSC Adv., 2015, 5, 41576 RSC.
| This journal is © The Royal Society of Chemistry 2016 |