
Potential hepatoprotective effects of fullerenol nanoparticles on alcohol-induced oxidative stress by ROS
Y. Lia,
H. B. Luob,
H. Y. Zhangc,
Q. Guoc,
H. C. Yaoc,
J. Q. Lic,
Q. Changd,
J. G. Yangb,
F. Wangb,
C. D. Wange,
X. Yangc,
Z. G. Liu*a and
X. Ye*b
aSchool of Biology and Pharmaceutical Engineering, Wuhan Polytechnic University, Wuhan 430023, P. R. China. E-mail: lzg_wpu@163.com; Fax: +86-27-83956793-208; Tel: +86-15327270600
bLiquor Marking Biological Technology and Application of Key Laboratory of Sichuan Province, College of Bioengineering, Sichuan University of Science & Engineering, Zigong 643000, P. R. China. E-mail: tornadoxin@163.com; Fax: +86-813-5505872; Tel: +86-18398707596
cSection of Environmental Biomedicine, Hubei Key Laboratory of Genetic Regulation and Integrative Biology, College of Life Sciences, Central China Normal University, Wuhan 430079, P. R. China
dDepartment of Chemical and Biomolecular Engineering, Hong Kong University of Science and Technology, Hong Kong Special Administrative Region, P. R. China
eSichuan University of Arts and Science, Dazhou 635000, P. R. China
First published on 22nd March 2016
Abstract
The free radical scavenging ability of fullerenols is their most exploited property in biomedical studies. The antioxidant properties and associated mechanisms of fullerenol have been previously investigated, but less research has been done to examine the hepatoprotective effects of fullerenol nanoparticles on alcohol-induced oxidative stress. This study examined the antioxidant effects in rat liver primary hepatocyte exposure to fullerenols dissolved in ethanol. In this study, liver primary hepatocytes of Wistar rats were divided into nine experimental groups that were exposed to ethanol (0.1, 1.0 and 10%), fullerenol dissolved in ethanol (0.1, 1.0 and 10%) and controls (PBS; PBS + fullerenol; Vitamin C (Vc) in 10% ethanol). The results demonstrated that fullerenol nanoparticles in 400 μmol L−1 exhibited excellent ROS scavenging abilities. This contributed to the anti-inflammatory effects involving reduced alcoholic oxidative damage and the regulated promotion of the tumor necrosis factor. The intracorporeal metabolism of rat-intake fullerenol was also evaluated by examining precision-cut rat slices of liver and kidney. All results demonstrated the potential hepatoprotective effects of fullerenol nanoparticles in the preventive treatment of alcoholic hepatopathy.
Introduction
Excessive alcohol consumption (EAC) is a worldwide problem that is associated with a significant increase in the risk of cancer in men.1,2 It is considered to be a major cause of morbidity and mortality. The liver is the most important metabolic organ due to its relationship to the gastrointestinal tract. The liver is therefore the primary target of alcohol metabolite associated hepatotoxicity. According to a WHO report, liver cancer is estimated to be responsible for 746![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 deaths globally each year.3 It is the second most common cause of cancer deaths in the world. Alcoholic liver injury is one of the major causes of liver cancer. Alcoholic fatty liver (AFL) is an initial symptom of alcoholic hepatitis caused by excessive, chronic alcohol consumption.4 AFL can further develop into alcoholic hepatitis,5 hepatic fibrosis6 and hepatocirrhosis.7 If left untreated these conditions may develop into liver cancer.
000 deaths globally each year.3 It is the second most common cause of cancer deaths in the world. Alcoholic liver injury is one of the major causes of liver cancer. Alcoholic fatty liver (AFL) is an initial symptom of alcoholic hepatitis caused by excessive, chronic alcohol consumption.4 AFL can further develop into alcoholic hepatitis,5 hepatic fibrosis6 and hepatocirrhosis.7 If left untreated these conditions may develop into liver cancer.
Many experimental and clinical studies have already reported that alcohol induced oxidative damage contributes to liver injury.8,9 Reactive oxygen species (ROS) play a key role in this hepatotoxicity mechanism.10 Acute and chronic ethanol treatments have been shown to increase the production of ROS, lower cellular antioxidant levels, and enhance peroxidation in many tissues, especially in the liver.11 The inflammation is associated with peroxidation of lipids, protein and DNA.12 ROS mediates the classic pathway from oxidative damage to pathogenesis. Over-expressed ROS destroy the innate antioxidant systems which comprise antioxidants such as glutathione (GSH), Vitamins C and E as well as small molecules.13 The generated product of lipid oxidation, malondialdehyde (MDA), could result in further damage to the DNA14 and trigger the expression of tumor necrosis factor (TNF).15 TNF is an important cytokine that regulates the internal inflammatory process. Chronic inflammation is associated with a high cancer risk,16 therefore, endosomatic ROS clearance is regarded as one of the most effective ways of protecting the liver from pathological changes and cancer.
Fullerenes have attracted considerable attention in the biomedical field since their discovery in 1985.17 The fullerene family, and especially C60, have potential as biological antioxidants. The antioxidant property is based on the fact that fullerenes possess a large number of conjugated double bonds and the low lying lowest unoccupied molecular orbital (LUMO) can easily take up electrons, making an attack of radical species highly possible.18 However, fullerene-C60 in polar solutions exhibit low solubility, which limits their use in the human body.19 In addition, the potential cytotoxicity of fullerene-C60 is also of great concern.20 Chemical modification of C60 double bonds is currently an internationally popular method to provide better properties for fullerene derivatives. The 30 double bonds of C60 are potential modification sites for generating new derivatives with particular desired functions. Hydroxylated fullerenes, also known as fullerenols, have attracted a lot of attention as an important class of fullerene derivatives. Previous studies of fullerenols have demonstrated their good water solubility as well as being non-cytotoxic.21–24 Researchers have also reported that fullerenols exhibit powerful antioxidant capabilities making them useful as potential antioxidative agents for biological systems.25,26 For example, recent in vitro and in vivo studies have demonstrated that fullerenol C60(OH)24 has a higher antioxidant activity than that of natural antioxidants such as Vitamins A, C and E.27,28 Animal experiments have shown that the liver is the primary organ that absorbs fullerenol. This absorbed fullerenol is then excreted via the digestive tract and the kidneys.29 Therefore, fullerenol is quite promising as a liver tissue-protective agent due to its biocompatibility and radical scavenging activity.30,31 Few studies, however, investigate the hepatoprotective effects of fullerenol in alcoholic liver injury.
The rat primary cultured hepatocyte is the most accepted model for hepatotoxicity and hepatopathy studies.32,33 In this study, we examined the hepatoprotective effects of fullerenol after rat hepatocytes were subjected to ethanol at different concentrations. The US CDC and US NIAAA (National Institute of Alcohol Abuse and Alcoholism) have established levels of human alcohol intake. Using these established levels as a reference, we used ethanol concentrations of 0.1%, 1% and 10% in our study.34 Biomarkers of oxidative stress (ROS, MDA, GSH), DNA damage (8-hydroxy-2′-deoxyguanosine, 8-OHdG) and tumor necrosis factor (TNF-α) were examined. The objective was to identify the hepatoprotective effects of fullerenol in preventive treatment of alcohol induced oxidative stress. The intracorporeal metabolism of rat-intake fullerenol was also evaluated by examining precision-cut slices of liver and kidney.
Results and discussion
Physicochemical characteristics of fullerenol
Size distributions of the tested material were measured using atomic force microscopy (AFM) and a scanning electron microscope (SEM). The AFM image shows the size distribution of dispersed fullerenols (Fig. 1B). Grain size in the nanoscale range was also observed (Fig. 1B). The SEM image in the top right corner demonstrates the morphology of a single fullerenol particle (Fig. 1B). The small size of nanoparticles facilitates the absorption of engineering materials across biological barriers or their uptake by cells.35,36 This capability allows the experimental fullerenol material to migrate across biological barriers and into target organs. The chemical structure of fullerenol was determined using an automatic elemental analyzer, thermogravimetric analysis and ATR-FTIR. | ||
| Fig. 1 Characterization of fullerenol. (A) Chemical structural formula; (B) images of AFM and SEM; (C) thermal analysis of fullerenol; (D) ATR-FTIR spectrum. | ||
The percentages of carbon and of hydrogen in the tested C60 derivative were determined by automatic elemental analyzer, and were found to be 34.855% and 2.955% respectively. 17 molecules of crystallization water loss were also detected by thermal analysis (Fig. 1C). From this analysis we found that the formula of the experimental C60 derivative was C60(OH)28·17H2O. The FTIR spectrum of the C60 derivative is shown in Fig. 1D. The FTIR spectrum of the C60 derivative contains four sharp absorption peaks at 1077, 1365, 1574, and 3233 cm−1. These peaks are attributed to γC–O, δC–O–H, γC![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) C, and γO–H, respectively.37 The result of the FTIR analysis is further evidence of a typical fullerenol chemical structure of experimental C60. The, chemical modification of the fullerene carbon cage by the attachment of functional groups (e.g. –OH) enhances its water solubility via hydrophilic functional adducts. The good solubility of fullerenol greatly improves its applicability as a biomedical agent in the human body.
C, and γO–H, respectively.37 The result of the FTIR analysis is further evidence of a typical fullerenol chemical structure of experimental C60. The, chemical modification of the fullerene carbon cage by the attachment of functional groups (e.g. –OH) enhances its water solubility via hydrophilic functional adducts. The good solubility of fullerenol greatly improves its applicability as a biomedical agent in the human body.
Fullerenol scavenges intracellular ROS induced by ethanol
Ethanol, oxidative stress and associated oxidative damage are mediators of cellular injury in many pathological conditions.38,39 Both clinical findings and results of experiments with animal models of alcoholic hepatopathy have shown these interactions in the onset of ethanol-induced liver damage.39 ROS generation is associated with ethanol stimulation. In the classic ethanol metabolizing pathway, the alcohol dehydrogenase enzyme reacts with the ethanol to form acetaldehyde which is highly unstable and results in the formation of free radicals. The observed intracellular ROS levels seen in this study support this theory.Acute exposures of ethanol with fullerenol to rat liver hepatocytes for 2 h were applied in intracellular ROS measurement. ROS generation occurs immediately after ethanol contact with cells. The method of combined exposure is to simulate instantaneous hepatoprotective effects of fullerenol in the process of alcohol consumption. Prepared rat liver hepatocytes were initially exposed to different concentrations of ethanol (0.1, 1.0 and 10%) for a 2 h incubation period at 37 °C in a CO2 incubator. Intracellular ROS increased in a dose-dependent manner (Fig. 2A). The ROS increases in all the ethanol groups were extremely significant (p < 0.001) as compared to the PBS group. Water-soluble fullerenols have been approved for their antioxidant ability to scavenge oxygen radicals, and for their ability to protect cells and/or tissues against ROS damage.40 In this test, the average ROS found in the samples containing fullerenol nanoparticles in PBS was almost equal to that found in the PBS control. Our results showed that the fullerenol nanoparticles had good biocompatibility (Fig. 2A and B). In addition, the fullerenol nanoparticles dissolved in ethanol (400 μmol L−1) demonstrated strong scavenging of the ethanol induced ROS (Fig. 2B). The groups with fullerenol nanoparticles dispersed in ethanol (0.1% and 1.0%) showed extremely significant differences (p < 0.001) when compared to the ethanol only groups (Fig. 2D). ROS averages for the fullerenol nanoparticles in ethanol (0.1 and 1.0%) were even closer to the control PBS group. Results demonstrated that fullerenol nanoparticles effectively quenched the ROS induced by ethanol (10%). However, it was not as effective with the lower ethanol concentrations of 0.1 and 1% (Fig. 2B). One possible reason is that 10% ethanol induced strong oxidative stress in the rat liver hepatocytes. Fullerenol at a higher concentration than 400 μmol L−1 may be better for quenching excessive ROS caused by 10% ethanol. However, as an engineering biomaterial, issues of biocompatibility and metabolism are greatest concern. Further studies of fullerenol are needed to ascertain the relationship between “effective dose” and “biocompatible concentration”. Fullerenol has been approved for its excellent antioxidant properties. Studies have demonstrated its antioxidant activity in in vitro and in vivo, is equal to or higher than that of natural antioxidants such as ascorbic acid (Vitamin C).41 Similar results have also been seen in prepared rat liver hepatocytes. Average ROS was obviously reduced (p < 0.001) in the group where fullerenol nanoparticles were dissolved in 10% ethanol. The ROS scavenging efficiency was being equal to that of Vitamin C (Fig. 2C). The fluorescence images of intracellular ROS are shown in Fig. 3A–I.
 | ||
| Fig. 2 Fluorescence intensity of intracellular ROS. (A) Ethanol with concentration of 0.1, 1.0 and 10%; (B) F + ethanol with concentration of 0.1, 1.0 and 10%; (C) the same amount of F and Vc in 10% ethanol; (D) comparison between groups of ethanol and F/ethanol (F-fullerenol nanoparticles; Vc-Vitamin C; **, p < 0.01; ***, p < 0.001). | ||
 | ||
| Fig. 3 Fluorescence images of intracellular ROS in liver primary hepatocytes (Nikon, Eclipse TS 100; 40×). (A) PBS; (B) fullerenol nanoparticles in PBS; (C) Vc in 10% ethanol; (D–F), F + ethanol with concentration of 0.1, 1.0 and 10%; (G–I), ethanol with concentration of 0.1, 1.0 and 10% (F-fullerenol nanoparticles; Vc-Vitamin C). | ||
Covalently attached groups of fullerenol might play an important role in ROS scavenging. This specific behavior of fullerenols is due to their structural flexibility, the rotation of the OH-groups around the C–O bond axis, and the distribution of these groups across different carbon sites of the fullerene surface.42 Based on this flexible structure, several mechanisms behind the antioxidant activity of fullerenol nanoparticles have recently been proposed. One such mechanism involves the radical reaction of hydroxyl radicals with the remaining olefinic double bonds of the fullerenol core. Fullerenol may also function as an antioxidant by donating a hydrogen atom from the hydroxyl group, to the particular radical compound. Another possible mechanism is for the oxygen radical to extract a hydrogen atom or an electron from the fullerenol to yield a relatively stable fullerenol. The free radicals based on the large numbers of conjugated double bonds with low energy, unoccupied molecular orbitals, can easily take up an electron and facilitate reactions with radical species.43,44
Hepatoprotective properties of fullerene nanoparticles: resistance to ethanol-induced oxidative damage and inflammation
Ethanol induced oxidative damage in the liver has been demonstrated by experimental data from rats in both acute and chronic tests.45 In general, cellular antioxidant defences are sufficient to keep the levels of ROS below a toxic threshold. However, toxic agent induced-ROS overproduction could destroy this natural anti-oxidant system.46 ROS-induced oxidative stress is always determined by measuring levels of biomacromolecules such as malondialdehyde (MDA) and glutathione (GSH). Glutathione (GSH) is an important antioxidant in this system. It is the classical biomarker used to reflect the status of oxidative stress that is indicated by the bioreaction of ROS-oxidized GSH into the oxidized form of glutathione (GSSH).In this study we found that GSH levels were greatly decreased after liver hepatocytes had been exposed to different concentrations of ethanol. The decrease followed a dose-dependent trend (Fig. 4A). The GSH decrease was significant in the 0.1% ethanol group (p < 0.01) and extremely significant in the 1.0 and 10% groups (p < 0.001) when compared to the PBS control group. The decrease in GSH levels slowed after fullerenol nanoparticles were added in the ethanol groups (Fig. 4B and D). This protective ability of GSH was equal to that of Vitamin C (Fig. 4C). This is due to the excellent ROS scavenging ability of fullerenol. Studies have shown that glutathione depletion can induce cell death.47 Our results for GSH demonstrate that the tested fullerenol nanoparticles can effectively reduce ethanol-associated oxidative stress.
 | ||
| Fig. 4 Intracellular GSH content. (A) Ethanol with concentration of 0.1, 1.0 and 10%; (B) F + ethanol with concentration of 0.1, 1.0 and 10%; (C) the same amount of F and Vc in 10% ethanol; (D) comparison between groups of ethanol and F/ethanol (F-fullerenol nanoparticles; Vc-Vitamin C; **, p < 0.01; ***, p < 0.001). | ||
Peroxidation is another consequence of oxidative stress. There is a relationship between lipid peroxidation and ROS production. Both isolated polyunsaturated fatty acids and those incorporated into lipids are easily attacked by free radicals and are oxidized to lipid peroxides.48 Malondialdehyde (MDA) is a metabolite of the lipid peroxidation of cell membranes. Increased lipid peroxidation is associated with alcohol induced pathological liver injury.49 Fig. 5A shows that MDA increases were dose dependent according to ethanol concentrations. Fullerenol nanoparticles effectively reduced the generation of MDA (Fig. 5B and D). The ability of fullerenol nanoparticles to reduce MDA was similar to that of Vitamin C (Fig. 5C). One possible mechanism to explain this is that tested fullerenol nanoparticles exhibited excellent ROS scavenging ability. In addition, fullerenol nanoparticles have numerous free oxygen electron pairs distributed around the fullerenol surface providing many opportunities to form coordinate bonds with pro-oxidant metal ions.50 This contributes to the protective abilities of the tested fullerenol nanoparticles against lipid peroxidation.
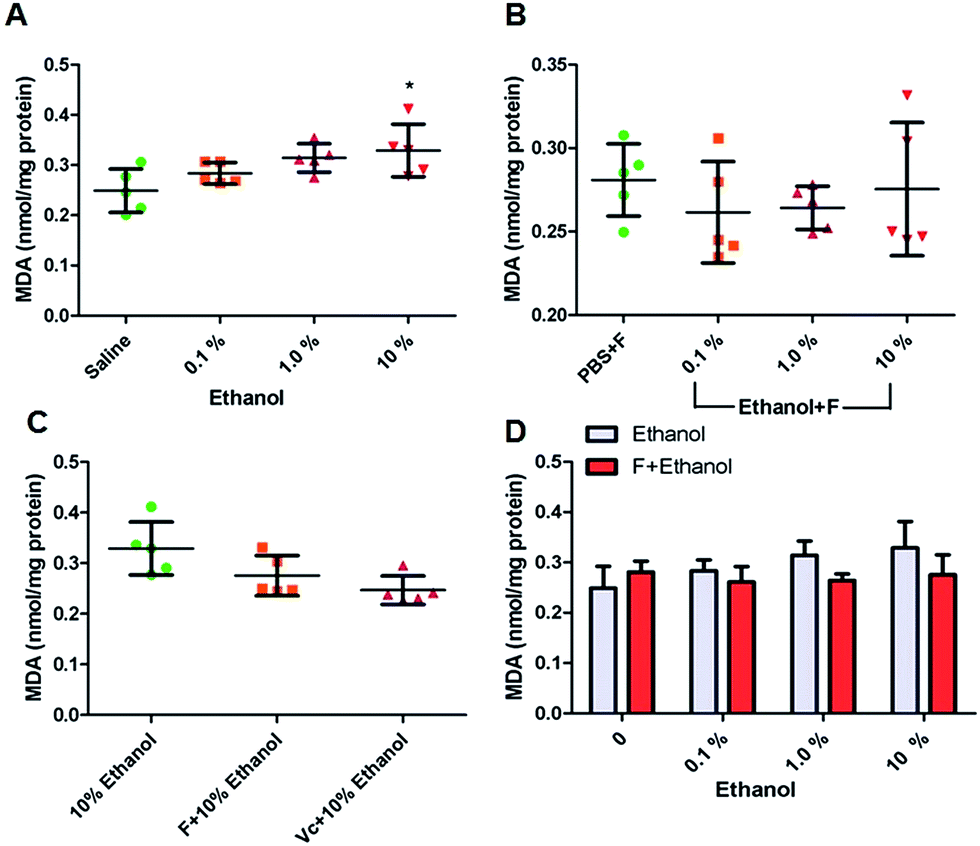 | ||
| Fig. 5 Intracellular MDA content. (A) Ethanol with concentration of 0.1, 1.0 and 10%; (B) F + ethanol with concentration of 0.1, 1.0 and 10%; (C) the same amount of F and Vc in 10% ethanol; (D) comparison between groups of ethanol and F/ethanol (F-fullerenol nanoparticles; Vc-Vitamin C; *, p < 0.05). | ||
Excessive activation of oxidative stress can lead to lipid peroxidation which is the leading cause for the expression and genetic mutation of proinflammatory cytokines that may result in cancer.51 For example, oxidative DNA damage is a major source of the mutation load in living organisms. There are more than one hundred oxidative DNA adducts identified (e.g. purine, pyrimidine, and the deoxyribose backbone). 8-OHdG is a pro-mutagenic DNA lesion resulting from oxidation damage. In this study, ethanol induced 8-OHdG was measured. No differences were detected in the control, ethanol or fullerenol/ethanol groups (Fig. 6A–D). Results indicated that a 2 h ethanol incubation may not induce any DNA damage. However, TNF-α in the liver hepatocytes showed an upward trend after exposure to ethanol (Fig. 7A). TNF-α is a pleiotropic cytokine that induces cellular responses such as proliferation, production of inflammatory mediators, and cell death. TNF-α is involved in pathological processes such as chronic inflammation. Studies have reported that TNF-α is involved in the pathophysiology of the liver. This includes viral hepatitis, alcoholic liver disease and non-alcoholic fatty liver disease.52–54 Other researchers have also demonstrated that TNF-α upexpression is associated with promotion and progression of cancer.55,56 As compared to ethanol groups, TNF-α expression was better controlled in the presence of fullerenol nanoparticles (Fig. 7B–D). Results clearly demonstrated that tested fullerenol nanoparticles exhibit great facility in anti-inflammation. The capabilities of fullerenol (ROS scavenging and anti-lipid peroxidation) demonstrated in this study contributed to these results.
 | ||
| Fig. 6 Intracellular 8-OHdG content. (A) Ethanol with concentration of 0.1, 1.0 and 10%; (B) F + ethanol with concentration of 0.1, 1.0 and 10%; (C) the same amount of F and Vc in 10% ethanol; (D) comparison between groups of ethanol and F/ethanol (F-fullerenol nanoparticles; Vc-Vitamin C). | ||
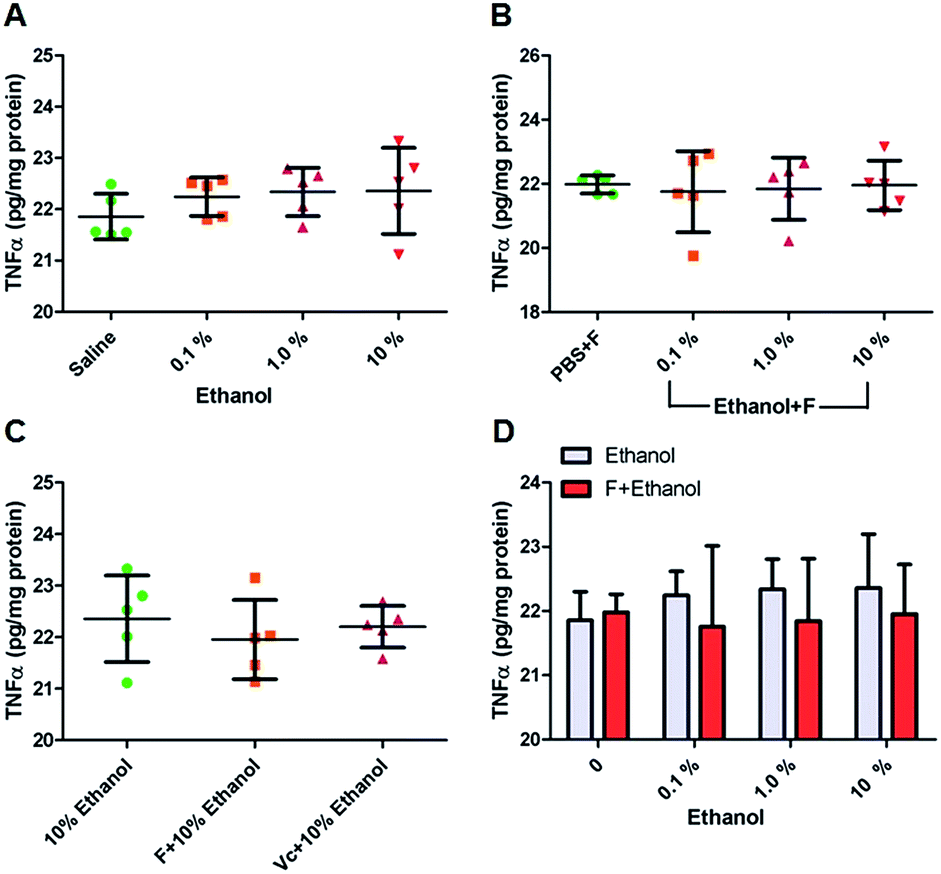 | ||
| Fig. 7 Intracellular TNF-α content. (A) Ethanol with concentration of 0.1, 1.0 and 10%; (B) F + ethanol with concentration of 0.1, 1.0 and 10%; (C) the same amount of F and Vc in 10% ethanol; (D) comparison between groups of ethanol and F/ethanol (F-fullerenol nanoparticles; Vc-Vitamin C). | ||
Biodistribution and excretion
Nanotechnology and nanomaterials have potential applications in various medical fields, such as diagnostics, imaging, gene and drug delivery and other types of therapy.57,58 Recently, much attention has been paid to the bioactive properties of water-soluble fullerenols.59 In this study, fullerenol nanoparticles exhibited a strong hepatoprotective effect. However, the issues of biodistribution and excretion are always of great concern.Many studies have reported that fullerenols can effectively distribute and aggregate in the rat liver. The liver is the second most absorptive organ next to the kidney. Ji et al. showed via a radiolabeled experiment, the kidney and liver are the main organs that absorb fullerenol.60 Additionally, results further demonstrated that most fullerenols were excreted from each organ 72 hours after dosing.61,62 In this study, rats were injected with fullerenol nanoparticles via tail intravenous injection. The two main metabolic organs, the liver and kidney, were studied using H&E-stained tissue slices (Fig. 8). Fine particle clusters were observed in the liver tissue slice (Fig. 8C). The observed results showed that injected fullerenol nanoparticles were successfully transferred to the target organ (the liver) after 2 hours from the time of dose delivery. Issues of biocompatibility and metabolism are of the great concern in preliminary studies of biomedical agents. A correlation study was also carried out using histopathological observations. No pathological inflammatory reactions were observed in the target organs (liver and kidney) after 7 days from the time of dosing (Fig. 8C, D, G and H), when compared to controls (Fig. 8A, B, E and F). Results indicate that fullerenol might be excreted from liver and kidney rather than deposited in these organs. One possible mechanism is that the fullerenol is excreted as metabolized products. Kubota et al.63 and Xu et al.64 independently demonstrated that water-soluble fullerenol could metabolize and be excreted via urine or feces. However, the detailed biodistribution of injected fullerenol nanoparticles in vivo is still unknown. An in vivo systematic study of metabolism and biological excretion is needed in further studies. In view of all these results, we found that fullerenol nanoparticles play an important role in hepatoprotection from alcohol-induced oxidative stress. In addition, since fullerenol nanoparticles are a type of biocompatible engineering material, these particles, intravenously injected, could be effectively delivered to the liver (Fig. 9). Liver cell injury is one of the effective models to study alcohol induced oxidative damage and hepatotoxicity. However, liver dysfunction is not limited to liver cell damage, but can also be a function of the internal structure of the organ. For example, a hepatic duct obstruction is a type of non-cytotoxic induced liver disease. A comprehensive study of the hepatoprotective effects of fullerenol still need to be verified by a series of functional hepatic tissue models.
 | ||
| Fig. 8 Biodistribution and excretion of fullerenol nanoparticles in metabolic organs of rat. (A) PBS-2 h (liver); (B) PBS-7 day (liver); (C) fullerenol-2 h (liver); (D) fullerenol-7 day (liver); (E) PBS-2 h (kidney); (F), PBS-7 day (kidney); (G) fullerenol-2 h (kidney); (H) fullerenol-7 day (kidney). | ||
 | ||
| Fig. 9 Mechanism describing the hepatoprotective effect of nano-fullerenol on alcohol-induced hepatotoxicity. | ||
Fullerenol was exhibited potential hepatoprotective effects depend on the existing essays. But the primary limitation of this study is that fullerenol and ethanol were delivered together, whereas the best exposure route would be to first dose the cells with ethanol, followed later by the fullerenol. Sequential dosing should be adopted in future work. It can better simulate the practical conditions of clinical application.
Experimental
Main reagents and kits
Fullerenol ultrafine-particles were purchased from Hengqiu Technologies (Suzhou, Jiangsu, People's Republic of China). Vitamin C was purchased from Sigma-Aldrich (St Louis, MO, USA). The ROS kit was purchased from Merck KGaA (Darmstadt, Germany). Rat ELISA kit for tumor necrosis factor (TNF)-α was purchased from eBioscience (San Diego, CA, USA). Rat enzyme-linked immunosorbent assay (ELISA) kits for 8-OHdG were purchased from Kamiya Biomedical Company (Seattle, WA, USA). Assay kits for MDA, GSH and protein were purchased from Beyotime (Nanjing, Jiangsu, People's Republic of China). The assay kit for determining protein content was purchased from Biotech (USA).Fullerenol and physicochemical characterization
:5:1 v/v/v]) for 24 hours at room temperature. Hematoxylin and eosin (H&E)-stained slices were then prepared as Liu described.66 Stained pieces were embedded in paraffin, sectioned into 10 μm slices, and observed using a DM 4000B microscope (Leica, Berlin, Germany). The average optical density (OD) of each slice was determined using Image-Pro Plus 6.0 (Media Cybernetics, Bethesda, MD, USA) software. All tissue sections were examined qualitatively by two experienced pathologists in a blinded fashion.Statistical analysis
GraphPad Prism software was used for statistical analysis of the experimental data, and for graphing the results. Results were evaluated statistically using an analysis of variance (ANOVA) followed by the Tukey test to determine the significance of the differences between groups. p < 0.05 is considered significant, p < 0.01 is considered a very significant difference and p < 0.001 is an extremely significant difference.Conclusions
Hepatoprotective effects of fullerenol nanoparticles in alcohol-induced oxidative stress were demonstrated in a rat liver primary hepatocyte. The fullerenol nanoparticles in 400 μmol L−1 exhibited excellent ROS scavenging ability. It effectively eliminated alcoholic oxidative damage and thus inhibited the alcohol-induced over expression of TNF-α. Moreover, fullerenol nanoparticles are easily absorbed by the liver and are excreted via the metabolic organs within a short period of time. Fullerenol nanomaterials are potential biocompatible biomedical agents that could be used in preventive treatment of alcoholic hepatopathy.Acknowledgements
This work was supported by the Open Research Fund of the Key Laboratory of Sichuan Province of China (NJ2011-01), Talent Foundation of Sichuan University of Science & Engineering (2015RC13) and The Innovation and Entrepreneurship Students Training Project of Sichuan Province of China (201510622015).References
- F. Donato, A. Tagger, R. Chiesa, M. L. Ribero, V. Tomasoni, M. Fasola, U. Gelatti, G. Portera, P. Boffetta and G. Nardi, Hepatology, 1997, 26, 579–584 CrossRef CAS PubMed.
- R. Baan, K. Straif, Y. Grosse, B. Secretan, F. EI Ghissassi, V. Bouvard, A. Altieri and V. Cogliano, WHO International Agency for Research on Cancer Monograph Working Group, Lancet Oncol., 2007, 8, 292–293 CrossRef PubMed.
- The International Agency for Research on Cancer. GLOBOCAN, Estimated cancer incidence, mortality and prevalence worldwide in 2012, World Health Organization, 2012 Search PubMed.
- C. S. Lieber, Alcohol, 2004, 34, 9–19 CrossRef CAS PubMed.
- A. G. Beckett, A. V. Livingstone and K. R. Hill, BMJ, 1961, 2, 1113 CrossRef CAS PubMed.
- R. Bataller and D. A. Brenner, J. Clin. Invest., 2005, 115, 209 CAS.
- T. A. Sørensen, K. Bentsen, K. Eghøje, M. Orholm, G. Høybye and P. Offersen, Lancet, 1984, 324, 241–244 CrossRef.
- B. Sid, J. Verrax and P. B. Calderon, Free Radical Res., 2013, 47, 894–904 CrossRef CAS PubMed.
- Y. Yang, Z. Han, Y. Wang, L. Wang, S. Pan, S. Liang and S. Wang, RSC Adv., 2015, 5, 36732–36741 RSC.
- D. F. Wu and A. I. Cederbaum, Semin. Liver Dis., 2009, 29, 141–154 CrossRef CAS PubMed.
- A. Dey and A. I. Cederbaum, Hepatology, 2006, 43, S63–S74 CrossRef CAS PubMed.
- A. I. Cederbaum, Y. Lu and D. Wu, Arch. Toxicol., 2009, 83, 519–548 CrossRef CAS PubMed.
- M. E. Medina, C. Iuga and J. R. Alvarez-Idaboy, RSC Adv., 2014, 4, 52920–52932 RSC.
- L. J. Marnett, Mutat. Res., 1998, 424, 83–95 CrossRef.
- J. C. Fernández-Checa, N. Kaplowitz, C. García-Ruiz, A. Colell, M. Miranda, M. Marí, E. Ardite and A. Morales, Am. J. Physiol., 1997, 273, 7–17 Search PubMed.
- L. J. Hofseth and M. J. Wargovich, J. Nutr., 2007, 137, 183S–185S CAS.
- H. W. Kroto, J. R. Heath, S. C. O'Brien, R. F. Curl and R. E. Smalley, Nature, 1985, 318, 162–163 CrossRef CAS.
- R. Bakry, R. M. Vallant, M. Najam-ul-Haq, M. Rainer, Z. Szabo, C. W. Huck and G. K. Bonn, Int. J. Nanomed., 2007, 2, 639–649 CAS.
- R. S. Ruoff, D. S. Tse, R. Malhotra and D. C. Lorents, J. Phys. Chem., 1993, 97, 3379–3383 CrossRef CAS.
- G. Jia, H. Wang, L. Yan, X. Wang, R. Pei, T. Yan and X. Guo, Environ. Sci. Technol., 2005, 39, 1378–1383 CrossRef CAS PubMed.
- K. Kokubo, S. Shirakawa, N. Kobayashi, H. Aoshima and T. Oshima, Nano Res., 2011, 4, 204–215 CrossRef CAS.
- G. V. Andrievsky, V. I. Bruskov, A. A. Tykhomyrov and S. V. Gudkov, Free Radical Biol. Med., 2009, 47, 786–793 CrossRef CAS PubMed.
- J. Mrdanovic, S. Solajic, C. Bogdanovic, K. Stankov, G. Bogdanovic and A. Djordevic, Mutat. Res., 2009, 680, 25–30 CAS.
- G. Bogdanovic, V. Kovic, A. Djordevic, J. Kanadanovic-Brunet, M. Vojinovic-Milardov and V. V. Baltic, Toxicol. In Vitro, 2004, 18, 629–637 CrossRef CAS PubMed.
- B. Srdjenovic, V. Milic-Torres, N. Grujic, K. Stankov, A. Djordjevic and V. Vasovic, Toxicol. Mech. Methods, 2010, 20, 298–305 CrossRef CAS PubMed.
- X. Yang, C. J. Li, Y. Wan, P. Smith, G. Shang and Q. Cui, Int. J. Nanomed., 2014, 9, 4023–4031 CrossRef PubMed.
- S. M. Mirkov, A. N. Djordjevic, N. L. Andric, S. A. Andric, T. S. Kostic, G. M. Bogdanovic and R. Z. Kovacevic, Nitric Oxide, 2004, 11, 201–207 CrossRef CAS PubMed.
- R. Injac, M. Perse, N. Obermajer, V. Djordjevic-Milic, M. Prijatelj, A. Djordjevic and B. Strukelj, Biomaterials, 2008, 29, 3451–3460 CrossRef CAS PubMed.
- W. H. Yang, D. B. Chen and X. M. Wang, Photochem. Photobiol. Sci., 2004, 28, 243–246 Search PubMed.
- B. Srdjenovic, V. Milic-Torres, N. Grujic, K. Stankov, A. Djordjevic and V. Vasovic, Toxicol. Mech. Methods, 2010, 20, 298–305 CrossRef CAS PubMed.
- J. Y. Xu, Y. Y. Su, J. S. Cheng, S. X. Li, R. Liu, W. X. Li and Q. N. Li, Carbon, 2010, 48, 1388–1396 CrossRef CAS.
- K. Tanaka, M. Sato, Y. Tomita and A. Ichihara, J. Biochem., 1978, 84, 937–946 CAS.
- W. X. Ding, H. M. Shen, H. G. Zhu and C. N. Ong, Environ. Res., 1998, 78, 12–18 CrossRef CAS PubMed.
- T. S. Naimi, D. E. Nelson and R. D. Brewer, Am. J. Prev. Med., 2010, 38, 201–207 CrossRef PubMed.
- N. Wang, L. Sun, X. Zhang, X. Bao, W. Zheng and R. Yang, RSC Adv., 2014, 4, 25886–25891 RSC.
- V. C. Sanchez, A. Jachak, R. H. Hurt and A. B. Kane, Chem. Res. Toxicol., 2011, 25, 15–34 CrossRef PubMed.
- K. Kobayashi, H. Ueno, K. Kokubo, M. Yudasaka and H. Yasuda, Carbon, 2014, 68, 346–351 CrossRef CAS.
- Z. Marković and V. Trajković, Biomaterials, 2008, 29, 3561–3573 CrossRef PubMed.
- J. B. Hoek and J. G. Pastorino, Alcohol, 2002, 27, 63–68 CrossRef CAS PubMed.
- J. Safaei-Ghomi and R. Masoomi, RSC Adv., 2014, 4, 2954–2960 RSC.
- R. Injac, M. Perse, N. Obermajer, V. Djordjevic-Milic, M. Prijatelj, A. Djordjevic and B. Strukelj, Biomaterials, 2008, 29, 3451–3460 CrossRef CAS PubMed.
- I. Rade, G. Biljana, D. Aleksandar and S. Borut, Afr. J. Biotechnol., 2008, 7, 4940 CAS.
- A. Djordjevic, B. Srdjenovic, M. Seke, D. Petrovic, R. Injac and J. Mrdjanovic, J. Nanomater., 2015, 2015, 1–15 CrossRef.
- A. Djordjevic, J. M. Canadanovic-Brunet, M. Vojinovic-Miloradov and G. Bogdanovic, Oxid. Commun., 2004, 27, 806–812 CAS.
- R. Nordmann, C. Ribire and H. Rouach, Alcohol Alcohol., 1990, 25, 231–237 CAS.
- J. Li, L. Li, X. Liu, R. Li and X. Yang, RSC Adv., 2013, 3, 25388–25395 RSC.
- J. B. Schulz, J. Lindenau, J. Seyfried and J. Dichgans, Eur. J. Biochem., 2000, 267, 4904–4911 CrossRef CAS PubMed.
- B. Halliwell and S. Chirico, Am. J. Clin. Nutr., 1993, 57, 715S–724S CAS.
- R. Polavarapu, D. R. Spitz, J. E. Sim, M. H. Follansbee, L. W. Oberley, A. Rahemtulla and A. A. Nanji, Hepatology, 1998, 27, 1317–1323 CrossRef CAS PubMed.
- R. Anderson and A. R. Barron, J. Am. Chem. Soc., 2005, 127, 10458–10459 CrossRef CAS PubMed.
- J. E. Klaunig, L. M. Kamendulis and B. A. Hocevar, Toxicol. Pathol., 2010, 38, 96–109 CrossRef CAS PubMed.
- R. F. Schwabe and D. A. Brenner, Am. J. Physiol., 2006, 290, G583–G589 CAS.
- L. Xu, Y. Wei, D. Dong, L. Yin, Y. Qi, X. Han and J. Peng, RSC Adv., 2014, 4, 30704–30711 RSC.
- S. Ceccarelli, N. Panera, M. Mina, D. Gnani, C. De Stefanis, A. Crudele, C. Rychlicki and A. Alisi, Oncotarqet., 2015, 6, 41434–41452 Search PubMed.
- F. Balkwill, Cancer Metastasis Rev., 2006, 25, 409–416 CrossRef CAS PubMed.
- H. Walczak, Immunol. Rev., 2011, 244, 9–28 CrossRef CAS PubMed.
- B. A. Holm, E. J. Bergey, T. De, D. J. Rodman, R. Kapoor, L. Levy and P. N. Prasad, Mol. Cryst. Liq. Cryst., 2002, 374, 589–598 CrossRef CAS.
- J. P. M. Almeida, A. L. Chen, A. Foster and R. Drezek, Nanomedicine, 2011, 6, 815–835 CrossRef CAS PubMed.
- J. Grebowski, P. Kazmierska and A. Krokosz, Biomed Res. Int., 2013, 2013, 1–9 CrossRef PubMed.
- Z. Q. Ji, H. Sun, H. Wang, Q. Xie, Y. Liu and Z. Wang, J. Nanopart. Res., 2006, 8, 53–63 CrossRef CAS.
- R. Injac, N. Radic, B. Govedarica, M. Perse and A. Cerar, Pharmacol. Rep., 2009, 61, 335–342 CrossRef CAS PubMed.
- S. Trajković, S. Dobrić, V. Jaćević, V. Dragojević-Simić, Z. Milovanović and A. Đorđević, Colloids Surf., B, 2007, 58, 39–43 CrossRef PubMed.
- R. Kubota, M. Tahara, K. Shimizu, N. Sugimoto, A. Hirose and T. Nishimura, Toxicol. Lett., 2011, 206, 172–177 CrossRef CAS PubMed.
- J. Y. Xu, Y. Y. Su, J. S. Cheng, S. X. Li, R. Liu, W. X. Li and Q. N. Li, Carbon, 2010, 48, 1388–1396 CrossRef CAS.
- D. Wu, J. Lu and Y. Zhang, Toxicol. Appl. Pharmacol., 2013, 271, 127–136 CrossRef CAS PubMed.
- X. Liu, Y. Zhang, J. Li, D. Wang, Y. Wu, Y. Li and X. Yang, Int. J. Nanomed., 2014, 11, 823–839 Search PubMed.
| This journal is © The Royal Society of Chemistry 2016 |