
FTIR metabolomic fingerprint reveals different modes of action exerted by active pharmaceutical ingredient based ionic liquids (API-ILs) on Salmonella typhimurium†
P.
Mester
a,
A. K.
Jehle
a,
C.
Leeb
a,
R.
Kalb
b,
T.
Grunert‡
c and
P.
Rossmanith‡
*a
aDepartment of Veterinary Public Health and Food Science, Christian Doppler Laboratory for Monitoring of Microbial Contaminants, University of Veterinary Medicine, Veterinaerplatz 1, 1210 Vienna, Austria. E-mail: peter.rossmanith@vetmeduni.ac.at
bProionic Production of Ionic Substances GmbH, Proionic GmbH Parkring 18, 8074 Grambach, Austria
cDepartment of Pathobiology, Functional Microbiology, Institute of Microbiology, University of Veterinary Medicine, Veterinaerplatz 1, 1210 Vienna, Austria
First published on 18th March 2016
Abstract
Since their incorporation into various chemical and biochemical processes, ionic liquids (ILs) have now been found useful for biomedical applications, including active pharmaceutical ingredients (APIs) such as antimicrobial agents or antibiotics. Recently, synergistic API-ILs with great potential have been reported, which show either increased antimicrobial activity or the ability to overcome bacterial resistance. In this study a total of 19 API-ILs, based on the antibiotic nalidixic acid, combined with different cation species, were investigated for synergistic effects against the important foodborne pathogen Salmonella. Furthermore, 19 respective ILs with chloride as the anion were used to control the effects of the different cation species. The antimicrobial activities of all 38 ILs against six different Salmonella species, as well as two nalidixic acid-resistant S. typhimurium strains, were determined via the microbroth dilution assay. The response pattern of the main cellular constituents, namely proteins, carbohydrates, and lipids of the bacterial cells to the most promising API-ILs was further investigated by Fourier transform infrared (FTIR) spectroscopy. While a number of active API-ILs based on nalidixic acid could be synthesized, no evidence for synergistic effects, such as increased antimicrobial activity or the ability to overcome resistance was found with either microbiological or spectroscopic methods. However, it could be demonstrated for the first time that while the different IL species ([TC8MA]+ and [TMC16A]+) showed similar antimicrobial activity, the FTIR spectral patterns indicated changes in bacterial membrane fluidity suggesting different modes of action.
Introduction
Since their first introduction as novel solvents and enhanced reaction media, ionic liquids (ILs) have found their way into many different applications covering the entire spectrum of the natural sciences. In the context of this study, their recent application as “active” compounds as antimicrobials, disinfectants or active pharmaceutical ingredients is of particular interest.1–8 In respect of such applications, two major fields of IL research have emerged concerning ILs as active pharmaceuticals, leading to an increasing number of publications and patent applications. In one research field different groups over the last years have shown that the IL form can positively influence active pharmaceutical ingredients (APIs), rendering them more effective and bioavailable. This so called ionic liquid prodrug approach was demonstrated by Cojocaru et al. in 2013 and since then a number of applications have been published.2,9 This prodrug approach aims to utilize the unique properties of the IL molecule to improve APIs. The second research field focuses on creating ILs consisting of biological active cations and on biologically active anions to develop new advantageous, synergistic and especially tunable API-ILs. This approach significantly benefits from increased knowledge of IL ecotoxicity, especially acute toxicity, biostability and biodegradability, which have received increased attention over the last few years secondary to increasing IL applications.10–12 One of the first examples for this approach is didecyl dimethylammonium ibuprofen,7 which combines an antibacterial cation with an anti-inflammatory anion. More recently, the synthesis of ILs with an antibiotic anion moiety was reported independently by two research groups.6,13–15 Both groups reported that the combination of a hydrophobic organic cation with an antibiotic anion ([C16Pyr][ampicillin] and [TMC16A][nalidixate]) resulted in API-ILs that were more effective at inhibiting bacterial growth than their single components alone. For [C16Pyr][ampicillin] it was reported that in the IL form, bacterial resistance to ampicillin could actually be overcome. Given the fact that the spread of antibiotic resistance among bacteria is considered one of the most important health issues worldwide, the great potential of effective API-ILs becomes clear. Antibiotic misuse and treatment neglect over the past decades has already resulted in a dramatic increase in multidrug-resistant bacteria species that cannot be treated effectively anymore.16 The serious issue of bacterial resistance to active pharmaceuticals such as nalidixic acid is now of utmost importance.17 In the case of nalidixic acid, several different bacterial resistance mechanisms are known. These include substrate specific efflux pumps as well as point mutations in the enzyme gyrase, which is the primary target of nalidixic acid.Until now no detailed investigations concerning any possible synergistic effect of API-ILs at the cellular level have been reported. One method applicable for such an investigation is Fourier transform infrared (FTIR) spectroscopy, which has been previously applied in microbiological studies to whole cell metabolomic analysis.18,19 Unlike Nuclear Magnetic Resonance (NMR) spectroscopy and mass spectrometry (MS)-based metabolomics (GC-MS, LC-MS), in most cases, FTIR spectroscopy is unable to identify single metabolites by instead providing a global image of the sample.20 Bacterial metabolic fingerprints reflect the overall composition of the main cellular biochemical constituents and originate from overlapping absorption bands, which are assigned to the following cellular biomolecules: (1) 3000 to 2800 cm−1, dominated by fatty acid chains of various amphiphilic membrane components (e.g. phospholipids); (2) 1800 to 1500 cm−1, dominated by the conformation-sensitive amide I and amide II bands of proteins and peptides and averages the predominance of α-helix or β-sheet protein secondary structures of all proteins in the bacterial cell; (3) 1500 to 1200 cm−1, a region with complex absorption profiles from proteins as well as fatty acids; (4) around 1230 cm−1 and 1085 cm−1, the region of ‘phosphorus-containing biomolecules’ detects vibrations of phosphodiester functional groups of DNA/RNA polysaccharide backbone structures and/or phospholipids or phosphorus-containing carbohydrates such as teichoic and lipoteichoic acids of bacterial membranes; (5) 1200 to 800 cm−1, dominated by vibrations of various oligo- and polysaccharides and their specific type of glycosidic linkages present within the cell wall and potentially the bacterial capsule.21,22 It can be used directly on the sample at short acquisition time (<1 minute per spectrum) and low running costs (reagent-free, reusable sample holders).20 This makes FTIR spectroscopy perfectly suited as a high-throughput metabolic fingerprinting technique and due to its high discriminatory power it has been successfully applied to species level identification and discrimination, and particularly as a tool to follow phenotypic responses caused by abiotic and biotic perturbations.23–25 For instance, the influence of growth temperature, growth media composition, pH and surfactants were demonstrated to alter significantly the metabolic state as monitored by FTIR spectroscopy.26–30 The objective of this study was to investigate if ILs with nalidixate as anion demonstrate a synergistic effect against resistant Salmonella. In this way 19 different cations were combined with chloride or nalidixate and their respective minimal inhibitory concentrations (MIC) determined against eight Salmonella serovars and two Salmonella enterica strains possessing defined nalidixic acid resistance. In addition to traditional antimicrobial susceptibility, the metabolic stress response of Salmonella strains to API-ILs was investigated. FTIR spectroscopy-based bioassay was used in this respect to determine the presence and the extent of cellular stress, under the rationale that stressing conditions can alter the cell metabolome.
Results and discussion
Synergistic ILs
To examine the effects of nalidixate in the form of an ionic liquid, 19 different API-ILs were produced via the CBILS™ route from nalidixic acid. It was our objective to investigate their antimicrobial activities and possible synergistic effects, as well as their potential to overcome bacterial resistance to nalidixic acid, as has been reported for ampicillin-based API-ILs.6,13 Since the potential antibacterial activity of the respective API-IL cationic counter ion of should not be discounted, 19 respective ILs with a chloride anion were included in this study. To assess antimicrobial activity, minimal inhibitory concentrations (MICs) of all 38 ILs were determined against eight different Salmonella strains, including six distinct Salmonella subspecies as well as two S. typhimurium strains with distinct resistance against nalidixic acid. As there were no clear differences in the MIC patterns between either the six non-resistant or the two strains with distinct nalidixic acid resistance, only the respective MICs of S. typhimurium and ΔttrC are shown in Table 1. Results for the other six strains are shown in ESI Table S1.†| IL cation | MIC (μM) 24 h | ||||
|---|---|---|---|---|---|
| S. typhimurium | ΔttrC | ||||
| IL anion | |||||
| Chloride | Nalidixate | Chloride | Nalidixate | ||
| a n.t.-not tested. | |||||
| DMIM | [C1mim]+ | >1000 | 79.3 ± 27.5 | >1000 | >1000 |
| EMIM | [C2mim]+ | >1000 | 91.3 ± 0.0 | >1000 | >1000 |
| EDMIM | [C1C1mim]+ | >1000 | 38.4 ± 9.5 | >1000 | >1000 |
| BMIM | [C4mim]+ | >1000 | 98.4 ± 64.4 | >1000 | >1000 |
| PMIM | [C5mim]+ | >1000 | 94.8 ± 62.1 | >1000 | >1000 |
| OMIM | [C8mim]+ | >1000 | 161.8 ± 124.3 | >1000 | >1000 |
| TMA | [TMC1A]+ | >1000 | 50.5 ± 32.5 | >1000 | >1000 |
| TMBA | [TMC4A]+ | >1000 | 87.7 ± 20.1 | >1000 | >1000 |
| TMOA | [TMC8A]+ | >1000 | 50.0 ± 26.7 | >1000 | >1000 |
| TMC10A | [TMC10A]+ | >1000 | 72.4 ± 62.7 | >1000 | 579.2 ± 0.0 |
| TMC12A | [TMC12A]+ | >1000 | 74.4 ± 67.0 | >1000 | >1000 |
| TMC16A | [TMC16A]+ | 109.9 ± 14.1 | 14.7 ± 3.9 | 94.6 ± 25.2 | 87.1 ± 43.5 |
| TBMA | [TC4MA]+ | >1000 | 40.0 ± 30.7 | >1000 | >1000 |
| TOMA | [TC8MA]+ | 38.7 ± 0.0 | 34.7 ± 15.0 | 38.7 ± 0.0 | 52.1 ± 0.0 |
| EMMor | [Emmor]+ | >1000 | 104.5 ± 61.5 | >1000 | >1000 |
| Empip | [Empip]+ | >1000 | 50.7 ± 33.2 | >1000 | >1000 |
| BMPyr | [Bmpyr]+ | >1000 | 177.5 ± 143.8 | >1000 | >1000 |
| TBMP | [TC4MP]+ | >1000 | 87.1 ± 34.8 | >1000 | >1000 |
| TOMP | [TC8MP]+ | 37.1 ± 0.0 | 22.2 ± 5.5 | 37.1 ± 0.0 | 22.2 ± 5.5 |
| Nalidixin | Na+ | n.t. | 38.9 ± 19.9 | n.t. | >1000 |
Only three ILs with a chloride anion inhibited bacterial growth within the given test parameters (<1000 μM): [TMC16A][Cl], [TC8MA][Cl] and [TC8MP][Cl] had MICs between 30 and 110 μM for all Salmonella strains, which is in good accordance with previously published results correlating the lipophilicity of IL cations, e.g. the number of carbon atoms, with increasing toxicity.31,32 For the ILs with nalidixate as anion, two trends were clearly observed. Firstly, the activity of nalidixate was not compromised by being in IL form, due to the fact that the nalidixate ILs showed MICs in the same range as pure nalidixic acid. This indicates that the antimicrobial effectivity of “normal” sodium nalidixate and nalidixate in IL form is roughly equivalent. However, in the case of the nalidixic acid resistant Salmonella strains, the data revealed that none of the nalidixate ILs, with non-toxic cations, actually overcame the inherent resistance. For both inherent resistant strains none of these 16 ILs showed a MIC within the experimental parameters (Tables 1 and S1†).
However, for the resistant strains, three nalidixate ILs had lower MICs than sodium nalidixate alone. As shown in Fig. 1, the nalidixate-based ILs with antimicrobial activity against all tested strains belong to the same IL species that also demonstrated antimicrobial activity with chloride as anion. Indeed, the MICs of the nalidixate and chloride ILs show no distinct differences from each other.
 | ||
| Fig. 1 MICs of selected ILs against eight different Salmonella strains. All ILs were tested against Salmonella typhimurium, the two Nal resistant strains ΔinvA and ΔttrC, and the five Salmonella subspecies S. bongori, S. arizonae, S. indica, S. houtenae and S. salamae. | ||
Based on the results obtained by the microbiological tests it is not possible to claim that the nalidixate ILs show any synergistic activity as was reported by Seter et al.13,14 The group reported a one log scale lower optimal inhibitory concentration of [TMC16A][Nal] compared to either [TMC16A][lactate] or sodium nalidixate against different bacterial species, which are associated with biofouling in marine environments. However, in this study, no evidence of such a synergistic effect against the different Salmonella subspecies was found for any of the tested nalidixate-based ILs. Although it could be demonstrated that nalidixate is still active when in IL form (at least with non-toxic cations), the microbiological tests did not reveal any evidence of nalidixate-based ILs overcoming defined bacterial resistance. Thus, solely from the MIC data, it is not possible to determine if, in the case of [TMC16A][Nal], [TC8MA][Nal] and [TC8MP][Nal], the anion or the cation is the determining factor.
In light of the results obtained, at this point it seems questionable as to whether API-ILs based on nalidixic acid are a promising approach to counter microbe influenced corrosion as previously suggested. Although the current literature suggests that it is clear that the physico-chemical properties of APIs can be specifically altered and improved as an IL, too little is known about the bacterial response or the impact on their genetic background if exposed to ILs. Therefore, to avoid mistakes made in the past, from our perspective a better understanding of possible synergistic effects of API-ILs is absolutely necessary before releasing them into the environment. Otherwise the uncontrolled release of antibiotics will inevitably lead to a further increase in bacterial resistance to such compounds.
No evidence for synergistic effects between ILs either containing [Nal] or [Cl] anions as examined by FTIR spectroscopy
To determine whether the anion or the cation is the determining factor of the ILs with the lower MICs, the metabolic response of two Salmonella strains exposed to nalidixate ILs was investigated by FTIR spectroscopy. With the help of the FTIR data, it is possible to detect and determine metabolic changes of bacteria exposed to different growth conditions or different stresses. For all FTIR spectroscopy experiments, the ILs [TC8MA][Nal]/[Cl] and [TMC16A][Nal]/[Cl] were chosen as representatives for ILs that have similar MICs with either chloride or nalidixate as anion. [TMC4A][Nal]/[Cl] were selected as ILs that did not influence bacterial growth in its chloride form. Sodium nalidixate-treated cells and cells treated only with PBS were used as respective controls. Two different approaches were chosen in this study: in the long-term approach, the wild type and the nalidixic acid-resistant ΔttrC strains were grown in the presence of either sodium nalidixate or the respective ILs under the same conditions as described for the microbroth dilution assay, but in larger volumes. For each tested substance the sample with the highest antimicrobial concentration, which still permitted growth, was used for FTIR spectroscopic analysis. For the second, short-term approach, the same two Salmonella strains were exposed for one hour to the different ILs at toxic concentrations to investigate a short-term effect on the bacterial cells as previously described.30,31Visual spectral analysis revealed that the spectral windows from 3000 to 2800 cm−1 and 1800 to 1500 cm−1 were the regions with the most prominent effects of the different treatments on the Salmonella cells (Fig. 2). The respective spectral window from 3000 to 2800 cm−1 contains biochemical information about the membrane fatty acids, while the spectral window from 1800 to 1500 cm−1 can be assigned to amide I and amide II groups of proteins and peptides.21,22 Hierarchical cluster analysis (HCA) of vector normalized 2nd derivatives of FTIR spectral data was employed to assess the overall similarity between the different treatments on Salmonella cells. In order to extract spectroscopically relevant information and to increase discriminatory power, the whole recorded spectral range of 4000 to 500 cm−1 was limited to either 3000 to 2800 cm−1 or 1800 to 1500 cm−1.
 | ||
| Fig. 2 FTIR spectra of S. typhimurium after short-term stress treatment. (A) Exemplary spectra derived from PBS (green), [TC8MA][Cl] (blue) and [TMC16A][Cl] (red) are shown from the whole spectrum. (B and C) Spectra of the second derivative and vector-normalized are shown in the regions where most pronounced differences were observed (spectral range of 3000 to 2800 cm−1 (membrane lipids, B) and 1800 to 1500 cm−1 (protein region, C)). | ||
For the long-term approach, where S. typhimurium and ΔttrC cells were grown overnight in the presence of the six different ILs or controls, no spectral differences in the respective regions were observed and HCA revealed no distinct clustering between treatments (ESI Fig. S1†). These results demonstrate that the presence of the respective ILs in the growth media for both Salmonella strains (the concentration just below the respective MIC) did not lead to distinct cell responses detectable by FTIR spectroscopy.
Accordingly, for the short-term approach, IL or sodium nalidixate concentrations were chosen that led to a significant CFU reduction after one hour of exposure (ESI Table S2†). Due to the fact that in the preliminary experiments, no spectral differences could be found between S. typhimurium and ΔttrC cells after any treatment, we decided to perform the approach only with S. typhimurium (data not shown). In contrast to the results of the long-term approach, the results obtained in the short-term approach revealed a great deal of information.
Firstly, no spectral differences between the respective ILs with either chloride or nalidixate as anion could be found. For both investigated spectral regions, fatty acids and proteins, HCA did not differentiate between either [TMC16A][Cl]/[Nal] or [TC8MA][Cl]/[Nal] when compared to the PBS and sodium nalidixate-treated controls (Fig. 3 and S2†). However, a clear discrimination between the IL treated cells and the controls was found (Fig. 3) for both IL species in the fatty acid spectral region. In the protein spectral region there was no such clear distinction which correlated with the found effects of the different IL or control treatments (ESI Fig. S2†). One explanation for these findings might be the fact that the recorded spectra for most incubation conditions are mixtures from living and dead cells (ESI Table S2†). While the observed clustering effects in the in the fatty acid region correlated with the respective ILs with lipophilic cations, such correlation could not be observed in the protein region either because there was no IL specific effect or it was masked by effects of overall cell viability.
 | ||
| Fig. 3 Clustering and concentration dependent effects of [TC8MA][Cl] and [TMC16A][Cl] treatment on S. typhimurium in the spectral region of fatty acids (3000 to 2800 cm−1). (A and C) FTIR spectroscopy-based dendrogram of S. typhimurium after short-term treatments with [TC8MA][Cl]/[Nal]. (A) The HCA plot demonstrates a clear separation of cells treated with [TC8MA][Cl]/[Nal] from cells treated with sodium nalidixate or PBS in the spectral region of fatty acids. The cluster of [TC8MA][Cl]/[Nal] treated cells, however, shows no further subdivision depending on the anion chloride or nalidixate. (C) Similarly, the treatment with ionic liquids based on [TMC16A] leads to a clear separation from the controls, but no further subdivision is caused by the anion Cl or Nal. (B and D) FTIR spectra of S. typhimurium treated with three different concentrations of [TC8MA][Cl] (B) and [TMC16A][Cl] (D) in the spectral range of 3000 to 2800 cm−1, showing the concentration dependency of spectral changes. | ||
The differences between the two examined spectral regions are even more evident for the HCA and PCA of all investigated conditions depicted in Fig. 4 and S3.† The two main clusters found in the HCA for the protein spectral region correlate well with the respective viability of the treated cells without being able to differentiate between either the different IL species or the controls, while no clustering at all was found in the PCA (ESI Fig. S3†).
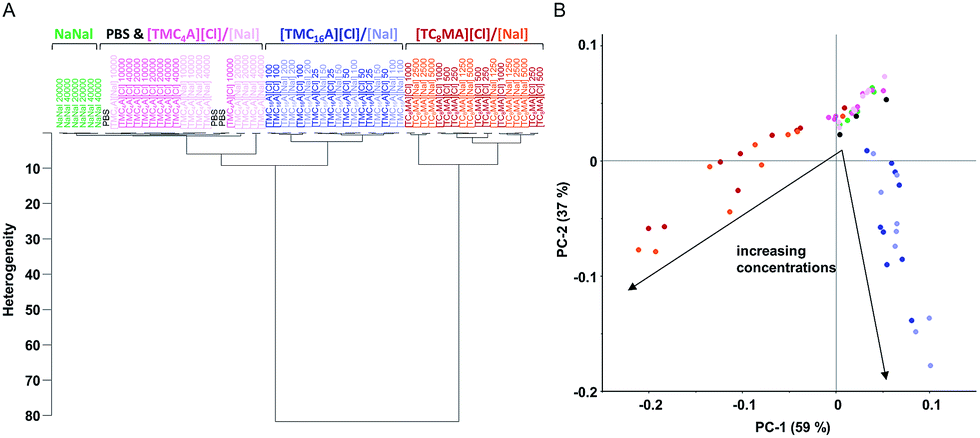 | ||
| Fig. 4 Clustering of different IL effects. FTIR spectroscopy-based analyses of S. typhimurium after short-term treatments. (A) In the region of fatty acids (3000 to 2800 cm−1), there is a clear clustering effect correlated with the cation. The bacteria treated with ionic liquids based on [TC8MA] and [TMC16A] clearly separate from the controls and from each other. In contrast, cells treated with [TMC4A][Cl]/[Nal] cluster with non-treated (PBS) and sodium nalidixate treated cells. In all three clusters ([TC8MA], [TMC16A] and [TMC4A] with controls), there is no subdivision depending of the anion Cl or Nal. (B) The principal component analysis demonstrates a clear distinction of [TC8MA] (red) and [TMC16A] (blue) treated cells between each other and the controls (PBS (black), NaNal (green) and [TMC4A] (pink)). Additionally, a clear concentration dependent effect is visible ([TC8MA] increasing concentrations from right to left and [TMC16A] increasing concentrations top down). | ||
In contrast, the HCA of the fatty acid spectral region revealed clear clustering of the different IL species, which can be correlated with their respective chemical structure (Fig. 4A). The PCA of the respective treatments additionally shows that the first two principal components, PC1 and PC2, revealed a clear concentration dependent clustering of spectral data according to treatment by [TC8MA] and [TMC16A] based ILs, respectively (Fig. 4B).
As is depicted in Fig. 4, incubation with the small and less lipophilic [TMC4A] cation had no effect in the fatty acid region, while strong effects were found for both the [TMC16A] and [TC8MA] based ILs. For both IL species clear qualitative and quantitative effects were found (Fig. 3), which were not very surprising given the surfactant-like structure of [TMC16A] and the strong lipophilicity of [TC8MA]. However, to our surprise, the HCA and PCA also revealed clearly distinguishable effects between these two IL cations.
FTIR spectroscopy indicates different modes of action of IL cations
Fig. 5 shows the spectral range dominated by fatty acids (3000 to 2800 cm−1) and demonstrates that S. typhimurium treated with ILs, containing either [TMC16A] or [TC8MA] as cations, resulted not only in a strong effect on cellular lipids compared to controls (PBS, sodium nalidixate), but also between the two distinctive cationic species. The most prominent differences were observed near 2923 cm−1 and near 2852 cm−1 (Fig. 5A), which correspond to the asymmetric νas(CH2) and symmetric νs(CH2) stretching mode of![[double bond splayed left]](https://www.rsc.org/images/entities/char_e009.gif) CH2 functional groups in fatty acids, respectively.19 Changes in band frequency of the conformation-sensitive νs(CH2) band near 2852 cm−1 have been used in previous publications to monitor membrane fluidity as a function of growth temperature, growth medium, pH, salt concentration in S. typhimurium and relate to desiccation and rehydration in E. coli, which, like S. typhimurium, is also a Gram-negative organism.33,34
CH2 functional groups in fatty acids, respectively.19 Changes in band frequency of the conformation-sensitive νs(CH2) band near 2852 cm−1 have been used in previous publications to monitor membrane fluidity as a function of growth temperature, growth medium, pH, salt concentration in S. typhimurium and relate to desiccation and rehydration in E. coli, which, like S. typhimurium, is also a Gram-negative organism.33,34
 | ||
| Fig. 5 Effects of different IL treatments in the spectral range of fatty acids. FTIR spectra of S. typhimurium treated with [TMC16A][Cl]/[Nal] or [TC8MA][Cl]/[Nal] in the spectral range of 3000 to 2800 cm−1 (A). Short-term treatment with [TC8MA][Cl]/[Nal] resulted in higher frequencies of peak maxima of νas(CH2) (B) and νs(CH2) (C) compared to PBS control and [TMC16A]Cl. | ||
According to the literature, an increasing frequency of approximately 2 to 5 cm−1 upon transition of the membrane lipids from an ordered to a disordered state corresponds to higher membrane fluidity.35,36 In the case of [TC8MA] based ILs, an increased peak frequency by 2 cm−1 (2852 to 2854 cm−1) of the νs(CH2) peak after treatment was found, which likewise indicates increased membrane fluidity (Fig. 5C).
While treatment of S. typhimurium with [TMC16A] based ILs did not lead to a shift in the peak maxima to either higher or lower frequencies, as in the case of [TC8MA] based ILs, an increased peak intensity of the νs(CH2) and νas(CH2) spectral bands was detected (Fig. 5B and C). This indicates the presence of longer and/or more saturated acyl chains as previously shown in E. coli intact cells during recombinant protein expression and aggregation.37 Thus, these results suggest that [TC8MA] and [TMC16A] based ILs have different modes of action against bacterial cells, or, to be more specific, the bacterial membrane, despite having similar MICs.
Conclusions
Ionic liquids as active pharmaceutical ingredients have been reported as promising substances that can show synergistic effects in terms of their antimicrobial activity or by overcoming resistance. In this study, the antimicrobial activity of 19 different nalidixate-based ILs and their respective chloride counterparts were investigated for such synergistic effects. Microbiological as well as spectroscopic methods revealed that, in the case of the foodborne pathogen Salmonella, no synergistic effects of the API-ILs could be found. In the case of API-ILs with non-toxic cations, the activity of nalidixate as an antibiotic remained unchanged, both against susceptible and resistant strains. FTIR spectroscopic analysis of two IL species with similar high antimicrobial activity ([TC8MA] and [TMC16A]) did not distinguish between the anions (nalidixate and chloride) but differentiated clearly between the cations, indicating alterations in the bacterial membrane fluidity. Thus, FTIR spectroscopy revealed different modes of action of differentially modified ILs with similar MICs against S. typhimurium. The present study demonstrates that FTIR spectroscopy is a very informative, high-throughput method to determine IL-induced changes in bacteria on macromolar and metabolic levels. It is anticipated that FTIR spectroscopy, followed by chemometrics, is also suitable for developing statistical models capable of predicting changes in membrane fluidity or other cellular biochemical constituents associated with ILs.Experimental
Ionic liquids
In this study, 38 ILs in total were tested, comprising 19 different cations combined with either chloride ([Cl]) or nalidixate [Nal]) anions. The majority of the ILs were based either on the 1-alkyl-3-methylimidazolium cation [Cnmim]+ (n = 12) or the trimethylalkylammonium cation ([TMCnA]+ (n = 12). The other cation species included in this study were tributylmethylammonium [TC4MA]+, trioctylmethylammonium [TC8MA]+, tributylmethylphosphonium [TC4MP]+, trioctylmethylphosphonium [TC8MP]+, 1-ethyl-3-methylpiperidinium [Empip]+, 1-ethyl-1-methylmorpholinium [Emmor]+ as well as 1-butyl-3-methylpyrrolidinium [Bmpyr]+. All ILs were provided by Proionic GmbH (Grambach, Austria) with a nominal purity higher than 95%, or synthesized using the CBILS® route.38 A complete list of all ILs used in this study can be found in ESI Table S1.†Bacterial strains and growth conditions
The bacterial strains Salmonella enterica ssp. enetrica serovar Typhimurium, Salmonella bongori, Salmonella arizonae, Salmonella indica, Salmonella houtenae and Salmonella salamae were maintained at −80 °C using MicroBank™ technology (Pro-Lab Diagnostics, Richmont Hill, Canada) and were part of the collection of bacterial strains at the Institute of Milk Hygiene, Milk Technology and Food Science, University of Veterinary Medicine, Vienna, Austria. All bacterial strains were grown overnight in tryptone soya broth with 0.6% (w/v) yeast extract (TSB-Y; Oxoid, Hampshire, United Kingdom) at respective growth temperatures (37 °C for Salmonella).The S. typhimurium mutants ΔinvA and ΔttrC were provided by Andreas Bäumler (California, USA39,40) and in the nalidixic acid-resistant derivative of S. typhimurium strain was ATCC 14028 (IR715).
Minimum inhibitory concentration (MIC) assessment
Minimum inhibitory concentrations of the examined ILs were determined by the serial 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :2 dilution microtiter plate method in TSB-Y medium. The highest tested concentration of each substance was 1000 μM. To ensure a constant cell status at the beginning of each experiment, one ml of the respective overnight culture of either of each Salmonella strain was transferred into nine ml fresh TSB-Y medium and incubated for three hours at 37 °C to ensure that the cells were in the logarithmic growth-phase. Afterwards each well, containing serial diluted ILs, was inoculated with 5 × 105 CFU per ml of the respective bacterial cells. After inoculation with the respective bacteria, the 96-well microtiter plates (Corning B.V. Life Sciences, Amsterdam, Netherlands) were measured at 610 nm wavelength in a TECAN F100 microplate reader (Tecan Austria GmbH, Groeding, Austria) to check for possible interference of the ILs at the given wavelength. The microtiter plates were then incubated for 24 h at 37 °C and afterwards microbial growth was assessed by measuring absorbance at 610 nm. The MIC was defined as the lowest IL concentration where no growth of the respective bacteria could be measured within 24 h. Each experiment was performed on three different days and the calculated average MIC values, as well as the standard deviations, are presented. Each experiment included positive and negative controls for the respective Salmonella strains.41
:2 dilution microtiter plate method in TSB-Y medium. The highest tested concentration of each substance was 1000 μM. To ensure a constant cell status at the beginning of each experiment, one ml of the respective overnight culture of either of each Salmonella strain was transferred into nine ml fresh TSB-Y medium and incubated for three hours at 37 °C to ensure that the cells were in the logarithmic growth-phase. Afterwards each well, containing serial diluted ILs, was inoculated with 5 × 105 CFU per ml of the respective bacterial cells. After inoculation with the respective bacteria, the 96-well microtiter plates (Corning B.V. Life Sciences, Amsterdam, Netherlands) were measured at 610 nm wavelength in a TECAN F100 microplate reader (Tecan Austria GmbH, Groeding, Austria) to check for possible interference of the ILs at the given wavelength. The microtiter plates were then incubated for 24 h at 37 °C and afterwards microbial growth was assessed by measuring absorbance at 610 nm. The MIC was defined as the lowest IL concentration where no growth of the respective bacteria could be measured within 24 h. Each experiment was performed on three different days and the calculated average MIC values, as well as the standard deviations, are presented. Each experiment included positive and negative controls for the respective Salmonella strains.41
FTIR spectroscopy-based bioassay
Each cell suspension was centrifuged (5 min at 8000g), washed twice with PBS and re-suspended in 1.5 ml tubes with one ml of the respective test solution (standardized optical density OD610 = 5). The control was obtained by re-suspending the cells directly in PBS. All tests were carried out in triplicate in three independent experiments on different days. Tubes were incubated for one hour at 30 °C in a shaking incubator set at 250 rpm. After the incubation, each sample was centrifuged (5 min at 8000g), washed twice with one ml PBS to effectively remove free IL and finally re-suspended in 100 μl PBS (OD610 = 50).30 90 μl suspension aliquots were sampled for three independent FTIR measurements (30 μl each), while the remaining suspension was used for biocidal activity determination. The FTIR spectroscopy experiments were carried out with a TENSOR 27 FTIR spectrometer, equipped with an HTS-XT accessory for rapid automation of the analysis (BRUKER Optics GmbH, Ettlingen, Germany). FTIR measurements were performed in transmission mode. All spectra were recorded in the range between 4000 and 400 cm−1. Spectral resolution was set at 4 cm−1, sampling 256 scans per sample. The software OPUS version 6.5 (BRUKER Optics GmbH, Ettlingen, Germany) was used to carry out the quality test, baseline correction, vector normalization and the calculation of the first and second derivatives of spectral values.24,42Unsupervised multivariate statistics
The OPUS software was used for hierarchical cluster analysis (HCA). Only the spectral region that offered the maximum information and discriminatory power for characteristic cellular macromolecules was examined. The normalized 2nd-derivative of the membrane lipid region (3000 to 2800 cm−1) and protein region (1800 to 1500 cm−1) were selected. Dendrograms were generated by using Ward's algorithm with normalization to repro level 30.For further HCAs and principal component analyses (PCAs), triplicates of each day or nonuplicates of each treatment were combined using the averages of the 2nd derivative.
The PCA computations were based on the NIPALS algorithm and were performed using the software Unscrambler X applying the same spectral windows as above mentioned (CAMO Software, Oslo, Norway).43
Biocidal activity test
The biocidal activity tests were performed in parallel with the FTIR spectroscopy-based bioassay to compare the metabolomic damages with the loss of viability. Ten μl of each cell suspension, prepared for the FTIR analysis, were serial diluted to determine the viable cell count on TSA + Y plates. The biocidal effect of the tested compounds was highlighted as logCFU reduction induced at different concentrations compared to the CFU number in the PBS control sample.
Acknowledgements
Financial support by the Austrian Federal Ministry of Science, Research and Economy and the National Foundation of Research, Technology and Development is gratefully acknowledged. The authors thank Martin Bobal, Tobias Gundolf and Christian Robben for their technical assistance.References
- A. Busetti, D. E. Crawford, M. J. Earle, M. A. Gilea, B. F. Gilmore, S. P. Gorman, G. Laverty, A. F. Lowry, M. McLaughlin and K. R. Seddon, Green Chem., 2010, 12, 420–425 RSC.
- O. A. Cojocaru, K. Bica, G. Gurau, A. Narita, P. D. McCrary, J. L. Shamshina, P. S. Barber and R. D. Rogers, MedChemComm, 2013, 4, 559–563 RSC.
- D. Coleman, M. Spulak, M. T. Garcia and N. Gathergood, Green Chem., 2012, 14, 1350–1356 RSC.
- R. Ferraz, L. C. Branco, C. Prudencio, J. P. Noronha and Z. Petrovski, ChemMedChem, 2011, 6, 975–985 CrossRef CAS PubMed.
- R. Ferraz, L. C. Branco, I. M. Marrucho, J. M. M. Araujo, L. P. N. Rebelo, M. N. Da Ponte, C. Prudêncio, J. P. Noronha and E. Petrovski, MedChemComm, 2012, 3, 494–497 RSC.
- R. Ferraz, V. Teixeira, D. Rodrigues, R. Fernandes, C. Prudêncio, J. P. Noronha, Z. Petrovski and L. C. Branco, RSC Adv., 2014, 4, 4301–4307 RSC.
- W. L. Hough, M. Smiglak, H. Rodr¡guez, R. P. Swatloski, S. K. Spear, D. T. Daly, J. Pernak, J. E. Grisel, R. D. Carliss, M. D. Soutullo, J. Davis and R. D. Rogers, New J. Chem., 2007, 31, 1429–1436 RSC.
- T. J. Kemp, Sci. Prog., 2012, 95, 224–230 CrossRef CAS PubMed.
- D. Dobler, T. Schmidts, I. Klingenhöfer and F. Runkel, Int. J. Pharm., 2013, 441, 620–627 CrossRef CAS PubMed.
- M. C. Bubalo, K. Radoseviç, I. R. Redovnikoviç, J. Halambek and V. G. Srcek, Ecotoxicol. Environ. Saf., 2014, 99, 1–12 CrossRef PubMed.
- S. Morrissey, B. Pegot, D. Coleman, M. T. Garcia, D. Ferguson, B. Quilty and N. Gathergood, Green Chem., 2009, 11, 475–483 RSC.
- G. Quijano, A. Couvert, A. Amrane, G. Darracq, C. Couriol, P. Le Cloirec, L. Paquin and D. Carrie, Chem. Eng. J., 2011, 174, 27–32 CrossRef CAS.
- M. Seter, M. J. Thomson, J. Stoimenovski, D. R. MacFarlane and M. Forsyth, Chem. Commun., 2012, 48, 5983–5985 RSC.
- M. Seter, M. J. Thomson, A. Chong, D. R. MacFarlane and M. Forsyth, Aust. J. Chem., 2013, 66, 921–929 CrossRef CAS.
- C. Florindo, J. M. M. Araujo, F. Alves, C. Matos, R. Ferraz, C. Prudêncio, J. P. Noronha, Z. Petrovski, L. Branco, L. P. N. Rebelo and I. M. Marrucho, Int. J. Pharm., 2013, 456, 553–559 CrossRef CAS PubMed.
- K. Hegstad, S. Langsrud, B. T. Lunestad, A. A. Scheie, M. Sunde and S. P. Yazdankhah, Microb. Drug Resist., 2010, 16, 91–104 CrossRef CAS PubMed.
- J. E. Stevenson, K. Gay, T. J. Barrett, F. Medalla, T. M. Chiller and F. J. Angulo, Antimicrob. Agents Chemother., 2007, 51, 195–197 CrossRef CAS PubMed.
- D. Naumann, D. Helm and H. Labischinski, Nature, 1991, 351, 81–82 CrossRef CAS PubMed.
- D. Naumann, Encyclopedia of Analytical Chemistry, 2000, pp. 102–131 Search PubMed.
- P. Vernocchi, L. Vannini, D. Gottardi, F. Del Chierico, D. I. Serrazanetti, M. Ndagijimana and M. E. Guerzoni, Front. Cell. Infect. Microbiol., 2000, 2, 156 Search PubMed.
- D. Helm, H. Labischinski, G. Schallehn and D. Naumann, J. Gen. Microbiol., 1991, 137, 69–79 CrossRef CAS PubMed.
- M. Wenning, S. Scherer and D. Naumann, Handbook of Vibrational Spectroscopy, 2008, pp. 71–95 Search PubMed.
- A. Alvarez-Ordonez, D. J. M. Mouwen, M. Lopez and M. Prieto, J. Microbiol. Methods, 2011, 84, 369–378 CrossRef CAS PubMed.
- T. Grunert, A. Monahan, C. Lassnig, C. Vogl, M. Müller and M. Ehling-Schulz, PLoS One, 2014, 9(12), e115959 Search PubMed.
- M. Wenning and S. Scherer, Appl. Microbiol. Biotechnol., 2013, 97, 7111–7120 CrossRef CAS PubMed.
- O. Preisner, J. A. Lopes, R. Guiomar, J. MacHado and J. C. Menezes, Anal. Bioanal. Chem., 2007, 387, 1739–1748 CrossRef CAS PubMed.
- P. M. Wehrli, E. Lindberg, O. Svensson, A. Sparén, M. Josefson, R. H. Dunstan, A. E. Wold and J. Gottfries, J. Chemom., 2014, 28, 681–686 CrossRef CAS.
- C. L. Winder and R. Goodacre, Analyst, 2004, 129, 1118–1122 RSC.
- H. M. Al-Qadiri, M. Lin, M. A. Al-Holy, A. G. Cavinato and B. A. Rasco, J. Food Sci., 2008, 73, M54–M61 CrossRef CAS.
- L. Corte, M. Tiecco, L. Roscini, S. De Vincenzi, C. Colabella, R. Germani, C. Tascini and G. Cardinali, PLoS One, 2015, 10(1), e0115275 Search PubMed.
- P. Mester, M. Wagner and P. Rossmanith, Ecotoxicol. Environ. Saf., 2014, 111, 96–101 CrossRef.
- S. Stolte, M. Matzke, J. Arning, A. Boschen, W. R. Pitner, U. Welz-Biermann, B. Jastorff and J. Ranke, Green Chem., 2007, 9, 1170–1179 RSC.
- A. Alvarez-Ordez, J. Halisch and M. Prieto, Int. J. Food Microbiol., 2010, 142, 97–105 CrossRef.
- C. M. Scherber, J. L. Schottel and A. Aksan, Biochim. Biophys. Acta, Biomembr., 2009, 1788, 2427–2435 CrossRef CAS.
- Z. Kota, M. Debreczeny and B. Szalontai, Biospectroscopy, 1999, 5, 169–178 CrossRef CAS.
- B. Szalontai, Y. Nishiyama, Z. Gombos and N. Murata, Biochim. Biophys. Acta, Biomembr., 2000, 1509, 409–419 CrossRef CAS.
- D. Ami, A. Natalello, T. Schultz, P. Gatti-Lafranconi, M. Lotti, S. M. Doglia and A. de Marco, Biochim. Biophys. Acta, Proteins Proteomics, 2009, 1794, 263–269 CrossRef CAS.
- R. Kalb, W. Wesner, R. Hermann, M. Kotschan, M. Schelch, and W. Staber, WO2005021484, 2005.
- M. Raffatellu, M. D. George, Y. Akiyama, M. J. Hornsby, S. P. Nuccio, T. A. Paixao, B. P. Butler, H. Chu, R. L. Santos, T. Berger, T. W. Mak, R. M. Tsolis, C. L. Bevins, J. V. Solnick, S. Dandekar and A. J. Bäumler, Cell Host Microbe, 2009, 5, 476–486 CAS.
- S. E. Winter, P. Thiennimitr, M. G. Winter, B. P. Butler, D. L. Huseby, R. W. Crawford, J. M. Russell, C. L. Bevins, L. G. Adams, R. M. Tsolis, J. R. Roth and A. J. Bäumler, Nature, 2010, 467, 426–429 CrossRef CAS PubMed.
- A. G. Santos, B. D. Ribeiro, D. S. Alviano and M. A. Z. Coelho, RSC Adv., 2014, 4, 37157 RSC.
- T. Grunert, M. Wenning, M. S. Barbagelata, M. Fricker, D. O. Sordelli, F. R. Buzzola and M. Ehling-Schulz, J. Clin. Microbiol., 2013, 51, 2261–2266 CrossRef CAS PubMed.
- H. Martens and T. Næs, J. Chemom., 1991, 138, 147–156 Search PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c5ra24970h |
| ‡ These authors have contributed equally to this work. |
| This journal is © The Royal Society of Chemistry 2016 |