
Water soluble cationic porphyrin TMPipEOPP-induced G-quadruplex and double-stranded DNA photocleavage and cell phototoxicity†
Li-Na Zhu*ab,
Shan Shiab,
Lin Yangab,
Min Zhanga,
Ke-Ke Liuab and
Li-Na Zhangab
aDepartment of Chemistry, Tianjin University, Tianjin, 300072, China. E-mail: linazhu@tju.edu.cn; Fax: +86-22-27403475
bCollaborative Innovation Center of Chemical Science and Engineering (Tianjin), Tianjin, 300072, China
First published on 26th January 2016
Abstract
Photodynamic therapy (PDT) has attracted increasing interest as a promising and non-invasive method for cancer treatment. Double-stranded DNA (dsDNA) is one of the potential bio-targets in PDT, and a PDT drug that could photocleave telomeric G-quadruplex might efficiently kill tumor cells by shortening the telomere length. Herein, a water soluble cationic porphyrin derivative 5,10,15,20-tetra-{4-[2-(1-methyl-1-piperidinyl)ethoxy]phenyl} porphyrin (TMPipEOPP) was demonstrated to be a promising PDT agent. It shows no DNA damage ability in the dark, but efficiently cleaves both double-stranded plasmid DNA and telomeric G-quadruplex DNA under irradiation by producing reactive oxygen species 1O2. However, since TMPipEOPP has much higher binding affinity to G-quadruplex than to dsDNA, it can preferentially bind to and specifically photocleave telomeric G-quadruplex in the presence of dsDNA with identical nucleotide concentrations, thus suggesting that it might be used as a human telomere-targeted photosensitizer for PDT of tumors. Correspondingly, almost no cytotoxicity is observed for TMPipEOPP in the dark, but efficient PDT-induced apoptosis is shown towards human colon carcinoma cells (HCT-8).
Introduction
A main challenge faced by traditional cancer treatments (radiotherapy and chemotherapy) is the serious side effects caused by the loss of normal organ function. As one of the promising and non-invasive methods for cancer treatment, photodynamic therapy (PDT) has attracted increasing interest over the last few years due to its potential to selectively destroy malignant cells while sparing the normal tissues.1–3 Due to the high absorption ability to light and high electron excitation energy for strong exergonic electron transfer,4 porphyrins and their derivatives have been widely used as photosensitizers of PDT.5 However, poor water solubility of many porphyrins has greatly hampered their applications in biologically relevant conditions.In PDT, double-stranded DNA (dsDNA) is one of the potential bio-targets,6 and many PDT drugs can interact with dsDNA and cleave it. In fact, beside typical double-stranded helix, more and more non-canonical DNA structures have been discovered and many of them are demonstrated to be associated with human diseases.7–9 In these, G-quadruplex is a well-known one. G-quadruplex is a four-stranded structure formed by guanine (G)-rich DNA or RNA sequence.10,11 It has been widely accepted that G-rich telomeric sequence at the 3′-end of linear chromosome might fold into G-quadruplex,12 and the formation of such a structure can inhibit the activity of telomerase,13 which is activated in approximately 85% human malignant tumors. DNA end replication problem can lead to progressive shortening of telomere. However, in malignant tumors, end replication problem is overcome, and telomere length cannot be shortened to a critical value to result in apoptosis. If G-rich telomere region could be specifically recognized and cleaved into fragments by drugs, infinite hyperplasia ability of cancer cells might be effectively inhibited.
Because of the size matching between the porphyrin core and the G-quartet plane of G-quadruplex, porphyrin derivatives are considered as promising G-quadruplex ligands that can efficiently promote the formation of G-quadruplexes and stabilize them.14–21 Recently, we designed and synthesized a cationic porphyrin derivative 5,10,15,20-tetra-{4-[2-(1-methyl-1-piperidinyl)ethoxy] phenyl} porphyrin (TMPipEOPP, Scheme 1) with four bulky side arm substituents, and demonstrated that the introduction of the four bulky, cationic side arms not only confers TMPipEOPP with good water solubility, but also greatly increases its G-quadruplex recognition specificity against dsDNA, single-stranded DNA (ssDNA) and triplex DNA.22–24 In this study, we further demonstrated that TMPipEOPP might be used as a promising photosensitizer of PDT. It could cleave supercoiled plasmid DNA under irradiation but could not work in dark. However, under the same nucleotide concentration conditions, TMPipEOPP shows good telomeric G-quadruplex photocleavage specificity against dsDNA.
 | ||
| Scheme 1 Chemical structure of TMPipEOPP. | ||
Experimental
Materials and chemicals
5,10,15,20-tetra-{4-[2-(1-methyl-1-piperidinyl)ethoxy]phenyl} porphyrin (TMPipEOPP) was prepared according to the method reported by our group.22 Fluorescein (FAM)-labelled oligonucleotides (Hum51: FAM-(TTAGGG)8TTA; DS51: FAM-TGAGTGAGTGAGTGTGAGAGTGTGTTACACACTCTCACACTCACTCACTCA) and calf thymus DNA (CtDNA) were purchased from Sangon Biotech. Co. Ltd. (Shanghai, China). The concentrations of the oligonucleotides were represented as single-stranded concentrations, which were determined by measuring the absorbance at 260 nm using the molar absorption coefficient determined by a nearest neighbour approximation (http://www.idtdna.com/analyzer/Applications/OligoAnalyzer). The concentration of CtDNA was represented as nucleotide concentration, which was determined by the absorbance at 260 nm using the molar absorption coefficient of 6600 M−1 cm−1. Supercoiled pBR322 plasmid DNA was ordered from TaKaRa Biotech. Co. Ltd. (Dalian, China). Superoxide dismutase (SOD) was purchased from Dingguo Biotech. Co. Ltd. (Beijing, China). Other reagents, including Tris (tris(hydroxymethyl)aminomethane), HCl, KCl, MgCl2, KI, DMSO, NaN3, Na2EDTA (disodium ethylenediamine tetraacetic acid) and ethanol were gained from Sigma. All chemical reagents were of analytical grade and used without further purification.Plasmid DNA photocleavage study
Photocleavage of supercoiled pBR322 plasmid DNA by TMPipEOPP was studied by agarose gel electrophoresis. 25 μL reaction mixtures containing 25 mM Tris–HCl buffer (pH 7.4), 10 ng μL−1 of pBR322 DNA, 0.75 μM of TMPipEOPP were prepared. The reaction mixtures were incubated in dark or under sunlight irradiation at 25 °C for 2 h. To which 5 μL loading buffer (0.25% bromophenol blue, 0.25% xylene cyanol, 30% glycerol) was added to quench the cleavage reaction. Then, 25 μL was loaded onto the 1% agarose gel containing ethidium bromide (1 μg mL−1). The gel was run in TBE buffer (90 mM Tris–borate, pH 8.3, 20 mM Na2EDTA) at 80 V for 1 h and photographed under UV light. The extent of DNA cleavage was analyzed by the Gel-Pro Analyzer software. Supercoiled plasmid DNA values were corrected by a factor 1.3 based on average literature estimate of lowered binding of ethidium to this structure.25G-quadruplex DNA photocleavage study
G-quadruplex DNA photocleavage activity of TMPipEOPP was evaluated by 10% polyacrylamide gel electrophoresis. 25 μL reaction mixtures containing 25 mM Tris–HCl buffer (pH 7.4), 5 μM of Hum51, 100 mM KCl and 10 mM EDTA, were prepared. The mixtures were heated at 95 °C for 5 min and then cooled down to 25 °C, and incubated at this temperature for 30 min. To which 40 μM TMPipEOPP was added and mixed. Then, the mixtures were incubated in dark or under sunlight irradiation at 25 °C for 2 h. 5 μL loading buffer (0.25% bromophenol blue, 0.25% xylene cyanol, 30% glycerol) was added to quench the cleavage reaction, and 25 μL was used for 10% polyacrylamide gel electrophoresis analysis (80 V, 40 min). The gel was visualized with UV light, and the extent of DNA cleavage was analyzed by the Gel-Pro Analyzer software. To investigate TMPipEOPP-induced DNA photocleavage of G-quadruplex/dsDNA mixture, 25 μL reaction mixtures containing 25 mM Tris–HCl buffer (pH 7.4), 5 μM of Hum51, 5 μM of DS51, 100 mM KCl and 10 mM MgCl2, were prepared. Other procedures are same as above except that 18% polyacrylamide gel was used.UV-vis absorption spectroscopy
UV-vis absorption spectra were measured by a Cary 60 UV-vis spectrophotometer (Agilent Technologies). Samples containing 10 mM Tris–HCl buffer (pH 7.4), 10 μM Hum51 or DS51 (strand concentration) or 510 μM CtDNA (nucleotide concentration), 50 mM KCl and 1 mM Na2EDTA were prepared. The samples were heated at 95 °C for 5 min and then cooled down to 25 °C for 30 min. After overnight incubation at 4 °C, 5 μM of TMPipEOPP was added and the absorption spectra in the range of 350–800 nm were recorded.In vitro cytotoxicity assay
Standard 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetraazolium bromide (MTT) assay was used to estimate the cytotoxicity of TMPipEOPP in dark or under irradiation. Human colon carcinoma cells (HCT-8) were seeded in 96-well culture plates and incubated for 24 h in a 5% CO2 incubator. After sucking out the culture medium, fresh culture medium containing different concentrations of TMPipEOPP were added to each well separately. The cells were divided into two parts, one part was incubated in dark for 24 h and the other was incubated under sunlight for 1 h. After that, the culture medium containing TMPipEOPP was sucked out. The cells were washed with phosphate buffer solution (PBS) twice and then fresh culture medium was added. MTT dye solution (20 μL, 0.1 mg) was added to each well and wells were incubated for an additional 4 h. Then, 150 μL DMSO was added to dissolve the formazan crystal and the absorbance at 490 nm was measured for the calculation of the cell survival rate.Results and discussion
TMPipEOPP-induced photocleavage of dsDNA
Since dsDNA is one of the potential bio-targets of PDT, the photocleavage activity of TMPipEOPP towards dsDNA was studied firstly using pBR322 plasmid DNA with 4363 base pairs as model dsDNA. As shown in Fig. 1, DNA cleavage was not observed in dark neither in the absence nor presence of TMPipEOPP. With the irradiation of sunlight, however, supercoiled plasmid DNA (Form I) was nearly completely converted to the nicked form (Form II) in the presence of TMPipEOPP.25 In contrast, no DNA cleavage was observed without TMPipEOPP even under irradiation, thus demonstrating that the cleavage of supercoiled plasmid DNA is really caused by TMPipEOPP, and TMPipEOPP has the potential to be used as a photosensitizer for photocleavage of dsDNA. | ||
| Fig. 1 Photocleavage of pBR322 plasmid DNA by TMPipEOPP. Lane 1, DNA + TMPipEOPP in dark; lane 2, DNA + TMPipEOPP under irradiation; lane 3, DNA in dark; lane 4, DNA under irradiation. The experiments were conducted in 25 mM Tris–HCl buffer (pH 7.4), [pBR322 DNA] = 10 ng μL−1, [TMPipEOPP] = 0.75 μM, reaction time was 2 h. | ||
Then, the dsDNA photocleavage behaviors of TMPipEOPP were studied in detail in the following experiments. To further demonstrate the photocleavage of plasmid DNA is really caused by TMPipEOPP, the effect of TMPipEOPP concentration was investigated. The results showed that the photocleavage of plasmid DNA was closely relevant with the concentration of TMPipEOPP (Fig. 2). With the increase of TMPipEOPP concentration, the amount of supercoiled DNA decreased and that of nicked DNA increased at the same time. When the concentration of TMPipEOPP exceeded 0.75 μM, the supercoiled form was nearly completely converted to the nicked one. With the further increase of TMPipEOPP concentration, nicked DNA could be further cleaved and linear DNA (Form III) could be observed.
 | ||
| Fig. 2 TMPipEOPP concentration dependence of plasmid DNA photocleavage. The concentrations of TMPipEOPP are (lanes 1 → 11) 0, 0.10, 0.25, 0.50, 0.75, 1.00, 1.25, 1.50, 1.75, 2.00 and 2.25 μM. [pBR322 DNA] = 10 ng μL−1, reaction time was 2 h. | ||
As shown in Fig. 3, photocleavage activity of TMPipEOPP towards plasmid DNA is pH-dependent. And interestingly, TMPipEOPP displayed the highest dsDNA photocleavage activity at pH of 7.4, both increasing and decreasing pH resulted in reduced photocleavage activities. Our previous work and other works demonstrated that cationic porphyrins tended to assemble into slipped face-to-face self-aggregate(s) under acidic conditions.26–28 Thus, the increase of steric hindrance might hamper the interactions between TMPipEOPP and dsDNA. Under basic conditions, the increase of hydroxyl ion concentration would certainly weaken the electrostatic interaction between positively charged TMPipEOPP and negatively charged dsDNA. Above experimental results demonstrate that TMPipEOPP might be used as an effective PDT photosensitizer at physiological or nearly physiological pH conditions.
 | ||
| Fig. 3 pH dependence of TMPipEOPP-induced pBR322 DNA photocleavage. The pH of lanes (0 → 8) are: 4.0, 5.0, 6.0, 7.0, 7.4, 8.0, 9.0, 10.0. [pBR322 DNA] = 10 ng μL−1, [TMPipEOPP] = 0.75 μM, reaction time was 2 h. | ||
The experiment to explore reaction time dependence of TMPipEOPP-induced plasmid DNA photocleavage was also carried out. As shown in Fig. 4, with the increase of reaction time, the amount of supercoiled DNA incessantly decreased, accompanied by the continuous increase of nicked one. The electrophoresis image was analyzed by the Gel-Pro Analyzer software to construct a plot of the percentage of either DNA form versus reaction time (Fig. 4B). The experimental data in Fig. 4B could be fitted well with first-order consecutive kinetic equations:

| Form I: CIt = CI0e−k1t |

where CIt, CIIt and CIIIt represent the concentration of supercoiled DNA, nicked DNA and linear DNA, respectively; CI0, CII0, and CIII0 represent the initial concentration of supercoiled DNA, nicked DNA and linear DNA, respectively; k1 and k2 represent the rate constant of conversion of supercoiled DNA to nicked DNA and nicked DNA to linear DNA, respectively.

 | ||
| Fig. 4 Time dependence of TMPipEOPP-induced pBR322 DNA photocleavage. (A) Gel electrophoresis results. The reaction times for lanes 1 → 11 are 0, 10, 20, 30, 40, 50, 60, 80, 100, 120 and 140 min, respectively. (B) Densitometric quantitative results of the gel electrophoresis. The scatters represent experimental data. The lines represent fitting curves. | ||
The rate constants of 0.0230 ± 0.0015 min−1 (k1) and 0.00087 ± 0.00032 min−1 (k2) were obtained for the conversion of supercoiled to nicked DNA and nicked to linear form, respectively. Since k1 is much larger than k2, TMPipEOPP might be considered as a single strand cleavage agent, and the cleavage reaction is non-site-specific.
It is reported that the damage of PDT drugs to biomolecules relies on irradiation-induced production of singlet oxygen 1O2, which might further lead to the generation of various reactive oxygen species (ROS).5,29,30 To disclose the mechanism of TMPipEOPP-induced DNA photocleavage, the effects of several radical scavengers were examined (Fig. 5). Superoxide anion radical (˙O2−) scavenger like superoxide dismutase (SOD) did not show any apparent effect on the DNA photocleavage behavior of TMPipEOPP, while DNA photocleavage was remarkably suppressed by NaN3 (1O2 scavenger), which is consistent with the previous reports that porphyrins and porphyrin derivatives are good photosensitizers for 1O2.5,31,32 In the commonly used hydroxyl radical ˙OH scavengers (KI, DMSO and ethanol), only KI could inhibit the DNA photocleavage in some degree. This might be attributed to the decreased light absorption ability of TMPipEOPP caused by KI (Fig. S1†), but not the scavenging of ˙OH by KI. These results demonstrated that 1O2 is responsible for TMPipEOPP-induced plasmid DNA photocleavage.
 | ||
| Fig. 5 TMPipEOPP-induced pBR322 DNA photocleavage in the presence of different ROS scavengers. Lane 1, DNA only; lane 2, DNA + TMPipEOPP; lane 3, DNA + TMPipEOPP + SOD (500 U mL−1); lane 4, DNA + TMPipEOPP + NaN3 (10 mM); lane 5, DNA + TMPipEOPP + KI (10 mM); lane 6, DNA + TMPipEOPP + DMSO (100 mM); lane 7, DNA + TMPipEOPP + ethanol (100 mM). [TMPipEOPP] = 0.75 μM, reaction time is 2 h. | ||
TMPipEOPP-induced photocleavage of telomeric G-quadruplex
It has been widely accepted that telomere length maintenance by telomerase is important for tumour cell growth and survival,33 and more and more evidences show that G-rich telomere sequence might fold into a kind of unique DNA secondary structure, G-quadruplex.12,13 In our previous works, we have demonstrated that TMPipEOPP can be used as a specific G-quadruplex ligand showing fascinating G-quadruplex recognition specificity against dsDNA and ssDNA.22–24 Since TMPipEOPP preferentially binds to telomeric G-quadruplex other than dsDNA, we have reasons to assume that the ROS produced by TMPipEOPP upon irradiation might efficiently cleave telomeric G-quadruplex, thus depriving tumor cells of immortality ability by shortening the telomere length. To investigate the photocleavage ability of TMPipEOPP towards telomeric G-quadruplex, a fluorescein (FAM)-labeled oligonucleotide (Hum51) with 51-nucleotide human telomeric sequence was used. The fluorescence of FAM was used to judge the cleavage of Hum51 in gel electrophoresis assays. As shown in Fig. 6, in the absence of TMPipEOPP, comparable signal intensities were given by Hum51 with or without irradiation. In the presence of TMPipEOPP, however, the signal intensity of the electrophoretic band under irradiation was much weaker than that in dark, suggesting that TMPipEOPP can also be used as a photosensitizer for G-quadruplex photocleavage. Such a G-quadruplex photocleavage reaction is also non-site-specific, producing cleavage products with different lengths, which cannot be observed in gel. However, the extent of cleavage reaction can still be judged by comparing the signal intensity change before and after reaction. | ||
| Fig. 6 Photocleavage of Hum51 by TMPipEOPP. Lane 1, Hum51 in dark; lane 2, Hum51 under irradiation; lane 3, Hum51 + TMPipEOPP in dark; lane 4, Hum51 + TMPipEOPP under irradiation. The experiments were conducted in 25 mM Tris–HCl buffer (pH 7.4), [Hum51] = 5 μM, [TMPipEOPP] = 40 μM, reaction time was 2 h. | ||
Similar to TMPipEOPP-induced plasmid DNA photocleavage, Hum51 photocleavage was also TMPipEOPP concentration-dependent (Fig. 7). That is, with the increase of TMPipEOPP concentration, more and more Hum51 was cleaved. When 40 μM TMPipEOPP was added, about 81% Hum51 was cleaved compared to the blank control. This result demonstrated that the photocleavage reaction of Hum51 is really triggered by TMPipEOPP.
 | ||
| Fig. 7 TMPipEOPP concentration dependence of Hum51 photocleavage reaction. The concentrations of TMPipEOPP are (lanes 1 → 7): 0, 4, 8, 16, 24, 32, 40 μM. [Hum51] = 5 μM, reaction time was 2 h. | ||
pH dependence of TMPipEOPP-induced G-quadruplex photocleavage was also investigated (Fig. 8). The results of circular dichroism (CD) showed that pH had no effect on the secondary structures of duplex and G-quadruplex DNAs, and parallel/antiparallel-mixed G-quadruplex was formed by Hum51 in the tested pH range (4.0–10.0) (Fig. S2†). Different from TMPipEOPP-induced plasmid DNA photocleavage, TMPipEOPP-induced Hum51 photocleavage was only slightly affected by pH though the highest cleavage activity was still found at physiological pH condition (pH = 7.4). This is not surprising. Under acidic conditions, the strong interactions between TMPipEOPP and G-quadruplex, including end-stacking and electrostatic interactions,26 might promote the disassembly of TMPipEOPP self-aggregates, and Hum51/TMPipEOPP complex can also be formed. Although the increase of hydroxyl ion concentration under basic conditions would weaken the electrostatic interaction between TMPipEOPP and Hum51, end-stacking interaction between TMPipEOPP and Hum51 can also promote the formation of Hum51/TMPipEOPP complex. Overall, TMPipEOPP can be used as an efficient photosensitizer for telomeric G-quadruplex cleavage in a wide pH range. This is very interesting, because it has been reported that the pH value of tumor microenvironment might be a little different from that of the normal biological one.
 | ||
| Fig. 8 pH dependence of TMPipEOPP-induced Hum51 photocleavage. Lane 1, Hum51 without TMPipEOPP; lanes 2 → 7 are (2) pH 5.0; (3) pH 6.0; (4): pH 7.0; (5) pH 7.4; (6) pH 8.0; (7) pH 9.0. [Hum51] = 5 μM, [TMPipEOPP] = 40 μM, reaction time was 2 h. | ||
It is well known that K+ plays an important role in biological system, and it can efficiently promote the formation of G-quadruplexes and stabilize them. Therefore, the effects of K+ concentration on Hum51 photocleavage were investigated. The results (Fig. 9) showed that TMPipEOPP-induced Hum51 photocleavage is insensitive to the change of K+ concentration. Similar cleavage efficiencies were obtained for the reactions conducted at 5 mM and 100 mM K+. Considering that identical G-quadruplex structure was adopted by Hum51 under the two K+ conditions (Fig. S3†), above experimental results indicate that TMPipEOPP-induced telomeric G-quadruplex photocleavage can be conducted in a wide K+ concentration range, and also suggests that such a photocleavage reaction is insensitive to ionic strength.
 | ||
| Fig. 9 TMPipEOPP-induced Hum51 photocleavage in the presence of different concentrations of K+. Lane 1, Hum51 without TMPipEOPP; lane 2, 5 mM K+; lane 3, 100 mM K+. [Hum51] = 5 μM, [TMPipEOPP] = 40 μM, reaction time was 2 h. | ||
As shown in Fig. 10, with the increase of reaction time, the signal intensity of Hum51 became weaker and weaker, indicating that such a TMPipEOPP-induced Hum51 photocleavage is time-dependent. Analysis with first-order reaction kinetic equation gave a cleavage reaction rate constant of 0.0160 ± 0.0007 min−1 for 40 μM TMPipEOPP.
 | ||
| Fig. 10 Time dependence of TMPipEOPP-induced Hum51 photocleavage. (A) Gel electrophoresis results. The reaction times for lanes 1 → 7 are 0, 10, 30, 60, 90, 120 and 150 min, respectively. (B) Densitometric quantitative results of the gel electrophoresis. The scatters represent experimental data. The line represents fitting curve. | ||
Similar to TMPipEOPP-induced pBR322 DNA photocleavage, TMPipEOPP-induced Hum51 photocleavage was almost unaffected by SOD, DMSO and ethanol. However, the addition of both NaN3 and KI could inhibit the Hum51 photocleavage reaction in some degree. Considering KI-induced decrease of TMPipEOPP light absorption ability, we could conclude that 1O2, but not ˙O2− and ˙OH, is involved in Hum51 photocleavage (Fig. 11).
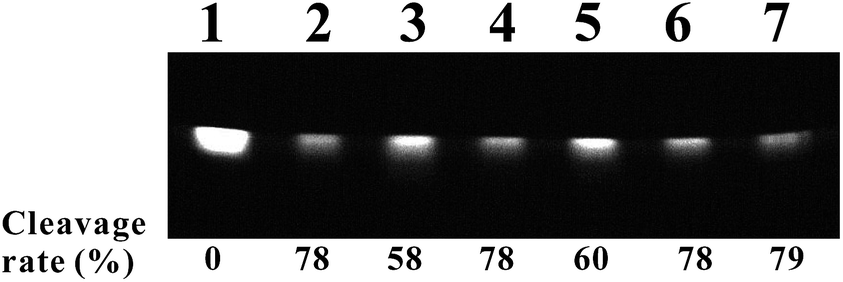 | ||
| Fig. 11 TMPipEOPP-induced Hum51 photocleavage in the presence of different ROS scavengers. Lane 1, DNA only; lane 2, Hum51 + TMPipEOPP; lane 3, Hum51 + TMPipEOPP + NaN3 (10 mM); lane 4, Hum51 + TMPipEOPP + SOD (500 U mL−1); lane 5, Hum51 + TMPipEOPP + KI (10 mM); lane 6, Hum51 + TMPipEOPP + DMSO (100 mM); lane 7, Hum51 + TMPipEOPP + ethanol (100 mM). [TMPipEOPP] = 40 μM, reaction time is 2 h. | ||
Since TMPipEOPP has high G-quadruplex recognition specificity against dsDNA, the photocleavage activities of TMPipEOPP towards G-quadruplex and dsDNA were compared. To eliminate the effects of DNA length and nucleotide concentration, a FAM-labeled double-stranded DNA (DS51) with 51 nucleotides was designed. Under the same conditions, TMPipEOPP showed high photocleavage activity towards Hum51, but only displayed very little effect on DS51 (Fig. 12), thus suggesting that TMPipEOPP has higher photocleavage activity to G-quadruplex than to dsDNA. Then, we investigated whether TMPipEOPP could specifically photocleave G-quadruplex in the presence of dsDNA. In this experiment, Hum51 and DS51 with identical nucleotide concentration were mixed. To achieve the efficient separation of Hum51 and DS51 in gel electrophoresis analysis, 18% polyacrylamide gel was used. To our pleasure, even in the presence of DS51, TMPipEOPP could still specifically recognize Hum51 and photocleave it. These results suggest that TMPipEOPP has the potential to specifically target at telomeric G-quadruplex regions even in the presence of other double-stranded gene sequences.
 | ||
| Fig. 12 (A) TMPipEOPP-induced Hum51 photocleavage in the presence of double-stranded DS51. Lane 1, Hum51 + TMPipEOPP (in dark); lane 2, Hum51 + TMPipEOPP (under irradiation); lane 3, DS51 + TMPipEOPP (in dark); lane 4, DS51 + TMPipEOPP (under irradiation); lane 5, Hum51 + DS51 + TMPipEOPP (in dark); lane 6, Hum51 + DS51 + TMPipEOPP (under irradiation). [TMPipEOPP] = 40 μM. (B) TMPipEOPP-induced pBR322 DNA photocleavage in the presence of Hum51; lane 1, pBR322 DNA + TMPipEOPP (in dark); lane 2, pBR322 DNA + TMPipEOPP (under irradiation); lane 3, pBR322 DNA + TMPipEOPP + Hum51 (in dark); lane 4, pBR322 DNA + TMPipEOPP + Hum51 (under irradiation). [TMPipEOPP] = 0.75 μM. [Hum51] = [pBR322 DNA] = 31 μM (in nucleotide). (C) TMPipEOPP-induced Hum51 photocleavage in the presence of pBR322 DNA; lane 1, Hum51 + TMPipEOPP (in dark); lane 2, Hum51 + TMPipEOPP (under irradiation); lane 3, Hum51 + TMPipEOPP + pBR322 DNA (in dark); lane 4, Hum51 + TMPipEOPP + pBR322 DNA (under irradiation). [TMPipEOPP] = 40 μM. [Hum51] = [pBR322 DNA] = 255 μM (in nucleotide). | ||
In this work, we firstly demonstrated that TMPipEOPP could trigger the photocleavage of double-stranded pBR322 plasmid DNA, and subsequently showed that TMPipEOPP could photocleave G-quadruplex Hum51 with high specificity in the presence of double-stranded DS51. The results seem to be contradictory. However, it should be noted that pBR322 DNA is a long dsDNA with 4363 base pairs (8726 nucleotides). DS51 is a short dsDNA with only 51 nucleotides. That is, the length of pBR322 DNA is about 171 times of DS51. In the 8726 nucleotides of pBR322 DNA, photocleavage at any site will result in the conversion of pBR322 DNA from supercoiled to nicked form. On the contrary, the photocleavage of DS51 needs that at least one site is damaged in 51 nucleotides. That is, the photocleavage of pBR322 DNA is much more easily observed than that of DS51.
Then, TMPipEOPP-induced DNA photocleavage was conducted on the Hum51/pBR322 DNA mixture. Because the length of pBR322 DNA is much longer than that of Hum51, TMPipEOPP-induced photocleavage of them cannot be simultaneously observed in one electrophoresis image. Therefore, the effects of Hum51 on pBR322 DNA photocleavage and pBR322 DNA on Hum51 photocleavage were investigated, respectively. As shown in Fig. 12B, the presence of Hum51 could greatly inhibit the photocleavage of pBR322 DNA. Under the same nucleotide concentrations, more than 60% pBR322 DNA photocleavage was inhibited by Hum51. On the contrary, pBR322 DNA showed much weaker effect on Hum51 photocleavage, obvious Hum51 photocleavage could also be observed in the presence of pBR322 DNA with the same nucleotide concentration (Fig. 12C), thus suggesting that effective G-quadruplex photocleavage could still be triggered by TMPipEOPP in the presence of long dsDNA.
Effects of DNAs on TMPipEOPP UV-vis absorption spectrum
To further explore the G-quadruplex photocleavage specificity of TMPipEOPP, the different TMPipEOPP-binding abilities of G-quadruplex and dsDNA were investigated by comparing the effects of them on the UV-vis absorption spectrum of TMPipEOPP. As shown in Fig. 13, free TMPipEOPP shows a strong Soret absorption band centered at 419 nm and four weak absorption bands centered at 521, 559, 593 and 650 nm. With the addition of Hum51, obvious hypochromicity was observed for the Soret band, accompanied by the emergence of two new absorption bands centered at 454 and 695 nm, respectively. The strong effects of Hum51 on TMPipEOPP absorption spectrum suggest that strong interactions occur between G-quadruplex and TMPipEOPP. That is, TMPipEOPP binds G-quadruplex by two synergetic interaction modes: π–π stacking interaction between the porphine core of TMPipEOPP and the G-quartet planes of G-quadruplex, and the electrostatic interaction between the positively charged side arms of TMPipEOPP and negatively charged phosphate back bones or grooves of G-quadruplex. On the contrary, dsDNAs, including the long dsDNA (calf thymus DNA, CtDNA) and the short one (DS51), gave much weaker effects on the TMPipEOPP absorption spectrum than Hum51. Besides no obvious hypochromicity was observed for the Soret band, only a very weak shoulder band emerged at 454 nm and no new absorption band appeared at 695 nm at all. These results suggest that the TMPipEOPP-binding ability of dsDNA is weaker than that of G-quadruplex, and the electrostatic interaction between the four positively charged side arms of TMPipEOPP and the negatively charged phosphate backbones of dsDNA might be the main binding mode between TMPipEOPP and dsDNA. As a result, TMPipEOPP can preferentially bind to G-quadruplex and photocleave it with high specificity even in the presence of dsDNA. | ||
| Fig. 13 UV-vis absorption spectra of TMPipEOPP in the absence or presence of different DNAs. [TMPipEOPP] = 5 μM. [Hum51] = [DS51] = 10 μM (strand concentration). [CtDNA] = 510 μM (nucleotide concentration). Each sample was replicated three times. | ||
Phototoxicity of TMPipEOPP to tumor cells
Standard 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetraazolium bromide (MTT) assay was used to estimate the cytotoxicity of TMPipEOPP. As shown in Fig. 14, after incubation of human colon carcinoma cell lines (HCT-8) with TMPipEOPP for 24 h in dark, the cell viability was still greater than 83%. However, under the irradiation of sunlight, one hour incubation of HCT-8 cells with TMPipEOPP would result in great decrease of the cell viability. For example, only about 30% cells survived after incubation with 2 μM TMPipEOPP for 1 h. If TMPipEOPP can be conjugated with other molecules or loaded in a nanocarrier to promote its aggregation in nucleus, the phototoxicity of TMPipEOPP to tumor cells might be further increased. These results suggest that TMPipEOPP has no or very low cytotoxicity in dark, but shows high phototoxicity towards tumor cells, thus might be used as an efficient photosensitizer in PDT. | ||
| Fig. 14 Cell viability of HCT-8 cells incubated with different concentrations of TMPipEOPP in dark for 24 h or under irradiation for 1 h. Each sample was replicated in six wells. | ||
Conclusions
In summary, the DNA cleavage ability and cytotoxicity of TMPipEOPP, a water soluble cationic porphyrin derivative with four bulky side arm substituents, were studied. The results show that no DNA cleavage ability is observed for TMPipEOPP in dark, however, irradiation of sunlight can lead to the generation of 1O2, thus render TMPipEOPP with the ability to photocleave both dsDNA and G-quadruplex DNA. Since TMPipEOPP has higher binding ability towards G-quadruplex than towards dsDNA, it can preferentially recognize and photocleave G-quadruplex with high specificity in the presence of dsDNA, thus making TMPipEOPP a potential drug targeting at human G-rich telomeric region. Cytotoxicity experiment shows that TMPipEOPP has no or very low cytotoxicity in dark, but displays high phototoxicity towards tumor cells, thus suggesting that TMPipEOPP might be used as an efficient photosensitizer in PDT.Acknowledgements
This work was supported by the Natural Science Foundation of China [No. 21371130] and the Natural Science Foundation of Tianjin [No. 15JCYBJC48300].Notes and references
- J. P. Celli, B. Q. Spring, I. Rizvi, C. L. Evans, K. S. Samkoe, S. Verma, B. W. Pogue and T. Hasan, Chem. Rev., 2010, 110, 2795–2838 CrossRef CAS PubMed.
- A. P. Castano, P. Mroz and M. R. Hamblin, Nat. Rev. Cancer, 2006, 6, 535–545 CrossRef CAS PubMed.
- L. Benov, Med. Princ. Pract., 2015, 24(1), 14–28 CrossRef PubMed.
- C. Zhou, Q. Liu, W. Xu, C. Wang and X. Fang, Chem. Commun., 2011, 47, 2982–2984 RSC.
- M. Ethirajan, Y. Chen, P. Joshi and R. K. Pandey, Chem. Soc. Rev., 2011, 40, 340–362 RSC.
- Y. Sun, Y.-J. Hou, Q.-X. Zhou, J.-R. Chen, B.-W. Zhang and X.-S. Wang, J. Inorg. Biochem., 2011, 105, 978–984 CrossRef CAS PubMed.
- O. Doluca, J. M. Withers and V. V. Filichev, Chem. Rev., 2013, 113, 3044–3083 CrossRef CAS PubMed.
- Y. Du and X. Zhou, Chem. Rec., 2013, 13, 371–384 CrossRef CAS PubMed.
- M. Métifiot, S. Amrane, S. Litvak and M. L. Andreola, Nucleic Acids Res., 2014, 42, 12352–12366 CrossRef PubMed.
- M. L. Bochman, K. Paeschke and V. A. Zakian, Nat. Rev. Genet., 2012, 13, 770–780 CrossRef CAS PubMed.
- S. Zhang, Y. Wu and W. Zhang, ChemMedChem, 2014, 9, 899–911 CrossRef CAS PubMed.
- C. Sissi and M. Palumbo, Curr. Pharm. Des., 2014, 20, 6489–6509 CrossRef CAS PubMed.
- Z. Crees, J. Girard, Z. Rios, G. M. Botting, K. Harrington, C. Shearrow, L. Wojdyla, A. L. Stone, S. B. Uppada, J. T. Devito and N. Puri, Curr. Pharm. Des., 2014, 20, 6422–6437 CrossRef CAS PubMed.
- S. Neidle and M. A. Read, Biopolymers, 2000-2001, 56, 195–208 Search PubMed.
- T. M. Ou, Y. J. Lu, J. H. Tan, Z. S. Huang, K. Y. Wong and L. Q. Gu, ChemMedChem, 2008, 3, 690–713 CrossRef CAS PubMed.
- X. X. Huang, L. N. Zhu, B. Wu, Y. F. Huo, N. N. Duan and D. M. Kong, Nucleic Acids Res., 2014, 42, 8718–8731 Search PubMed.
- L. Sabater, P. J. Fang, C. F. Chang, A. De Rache, E. Prado, J. Dejeu, A. Garofalo, J. H. Lin, J. L. Mergny, E. Defrancq and G. Pratviel, Dalton Trans., 2015, 44, 3701–3707 RSC.
- V. H. Le, N. Nagesh and E. A. Lewis, PLoS One, 2013, 8, e72462 CAS.
- J. Seenisamy, E. M. Rezler, T. J. Powell, D. Tye, V. Gokhale, C. S. Joshi, A. Siddiqui-Jain and L. H. Hurley, J. Am. Chem. Soc., 2004, 126, 8702–8709 CrossRef CAS.
- D. F. Shi, R. T. Wheelhouse, D. Sun and L. H. Hurley, J. Med. Chem., 2001, 44, 4509–4523 CrossRef CAS PubMed.
- H. Han, D. R. Langley, A. Rangan and L. H. Hurley, J. Am. Chem. Soc., 2001, 123, 8902–8913 CrossRef CAS PubMed.
- L.-N. Zhu, S.-J. Zhao, B. Wu, X.-Z. Li and D.-M. Kong, PLoS One, 2012, 7, e35586 CAS.
- L.-N. Zhu, B. Wu and D.-M. Kong, Nucleic Acids Res., 2013, 41, 4324–4335 CrossRef CAS PubMed.
- L.-N. Zhu, Y.-F. Huo and B. Wu, Anal. Methods, 2014, 6, 5067–5075 RSC.
- D.-M. Kong, J. Wang, L.-N. Zhu, Y.-W. Jin, X.-Z. Li, H.-X. Shen and H.-F. Mi, J. Inorg. Biochem., 2008, 102, 824–832 CrossRef CAS PubMed.
- Y.-Y. Liu, M. Wu, L.-N. Zhu, X.-Z. Feng and D.-M. Kong, Chem.Asian J., 2015, 10, 1304–1310 CrossRef CAS PubMed.
- K. Kano, H. Minamizono, T. Kitae and S. Negi, J. Phys. Chem. A, 1997, 101, 6118–6124 CrossRef CAS.
- S. Thyagarajan, T. Leiding, S. P. Arsköld, A. V. Cheprakov and S. A. Vinogradov, Inorg. Chem., 2010, 49, 9909–9920 CrossRef CAS PubMed.
- Y. Sun, L. E. Joyce, N. M. Dickson and C. Turro, Chem. Commun., 2010, 46, 6759–6761 RSC.
- Y. Sun, Y.-J. Hou, Q.-X. Zhou, J.-R. Chen, B.-W. Zhang and X.-S. Wang, J. Inorg. Biochem., 2011, 105, 9778–9984 Search PubMed.
- D. Ashen-Garry and M. Selke, Photochem. Photobiol., 2014, 90, 257–274 CrossRef CAS PubMed.
- R. W. Redmond and J. N. Gamlln, Photochem. Photobiol., 1999, 70, 391–475 CrossRef CAS PubMed.
- E. H. Blackburn, Angew. Chem., Int. Ed. Engl., 2010, 49, 7405–7421 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c5ra24964c |
| This journal is © The Royal Society of Chemistry 2016 |