DOI:
10.1039/C5RA22371G
(Paper)
RSC Adv., 2016,
6, 5249-5258
Reparative activity of costunolide and dehydrocostus in a mouse model of 5-fluorouracil-induced intestinal mucositis
Received
26th October 2015
, Accepted 24th December 2015
First published on 5th January 2016
Abstract
The aim of the study was to investigate the protective effects of costunolide (Co) and dehydrocostus (De) in 5-fluorouracil (5-FU)-induced intestinal mucositis (IM) as well as the potential mechanisms involved. Male Kunming mice were given 5-FU (60 mg kg−1 per day) by intraperitoneal injections for 5 consecutive days and IM was evaluated histochemically. Co (5, 20 mg kg−1) and De (5, 20 mg kg−1) were orally administered once daily for 8 days. Repeated 5-FU treatment caused severe IM including morphological damage, which was accompanied by feeding reduction, body weight loss and diarrhea. Daily intragastric administration of Co or De significantly relieved the severity of IM through promoting intestinal mucosal recovery, inhibiting reactive oxygen species and ameliorating the inflammatory responses. Accordingly, Co and De may be promising therapeutic candidates and clinically used for the prevention of IM during cancer chemotherapy.
1 Introduction
5-Fluorouracil (5-FU) is a main backbone agent in clinical malignant cancer practice.1 It exerts an anti-tumor effect by acting on the enzyme thymidylate synthetase, which in turn blocks DNA synthesis. However, 5-FU is burdened with toxicities that can induce dose-limiting, cause costly side effects and threaten the effectiveness of therapy. The targets of 5-FU rapidly divide cells indiscriminately, either malignant or healthy, including intestinal basal stem cells in crypts.2,3 Mucositis is one of the resultant damages induced by chemotherapy, it limits the clinical applications of 5-FU and no definitive prophylaxis or treatment exists.4 Approximately 50–80% of patients undergoing 5-FU treatment suffer from intestinal mucositis (IM) as a result of ulceration through to, and including, gastrointestinal symptoms such as abdominal bloating, vomiting and diarrhoea.5,6 Therefore, it is vital to do some pharmacological interventions in IM during 5-FU therapy. With a disruption of villi length and crypt cell homeostasis, IM induced by 5-FU is a consequence of oxidative stress, apoptosis, hypoproliferation and abnormal inflammation.3,7 Reactive oxygen species (ROS) is firstly stimulated by 5-FU-challenged, and then it activates transcription factors that lead to a suite of phenomena defined as acute tissue reactions. Nuclear factor kappa-B (NF-κB), as a key mediator, triggers the production of pro-inflammatory cytokines such as tumor necrosis factor-α (TNF-α) and interleukin (IL)-1β, which further amplify inflammatory signals via positive feedback loops, thus prolonging tissue injury and resulting in intestinal homeostasis disequilibrium and dysfunctions.
Costunolide (Co) and dehydrocostus (De) with a guaiane skeleton belonging to sesquiterpene lactones are the major active components of Radix Aucklandiae (Fig. 1). They possess a variety of pharmacological activities including anti-carcinogenesis, anti-ulcer, antimicrobial, anti-inflammatory and antifungal.8–10 Further mechanisms implicated in above activities are mainly focused on the inhibition of ROS, the amelioration of inflammatory responses and the improvement of immune system. In more specific terms, Co exerts inhibitory effect on ROS liberation, NF-κB activation, mitogen-activated protein kinase level and nitric oxide synthase (NOS) expression.11–14 De prevents the augment of TNF-α, activation of transcription-3 (STAT3), productions of nitric oxide (NO) with inducible nitric oxide synthase (iNOS) and levels of cyclooxygenase-2 (COX-2) as well as prostaglandin E2 (PGE2).13–16 What is notable that, our previous researches on gastrointestinal diseases manifested that Co and De could improve gastrointestinal function, relieve the spasm of smooth muscle as well as protect against intestinal carcinogenesis.17,18 Consequently, it is remarkable to explore the pharmacological action of Co and De in IM caused by chemotherapeutic agents, which is also significant to the further cancer treatment.
 |
| | Fig. 1 The chemical structures and essential group of Co and De. | |
In view of the potential anti-inflammatory advantage and gastrointestinal regulation, we set forward this study to investigate the possible effects of Co and De on IM induced by 5-FU as well as to explore their putative pathways of actions.
2 Material and methods
2.1 Drugs and reagents
5-FU injection (250 mg/10 mL) was purchased from Kingyork Amino Acid Co., Ltd (Tianjin, China). Loperamide (loper) was produced by Geri-care Pharmaceuticals Corp. (New York, America). Co and De were purchased from the National Institute for the Control of Pharmaceutical and Biological Products (Beijing, China). Purity of all reference compounds were over 98%.
2.2 Animals
Mice experiments were conducted under ethics approval from Institutional Animal Care and Use Committee of Institute of Radiation Medicine Chinese Academy of Medical Sciences (permit number: IACUC2014-010). Ninety-eight adult male Kunming (KM) mice (18–22 g) (license no. 0042643, SCXK (Jun)-2012-004) were provided by the Laboratory Animal Center of Health Science, Peking University, Beijing, China. Housed in polycarbonate cages (fourteen animals in each cage) with white wood chips for bedding, animals had free access to water and food. This study was carried out in accordance with the Regulation for the Administration of Affairs Concerning Experimental Animals.
2.3 Physical manifestations
IM was induced by intraperitoneal injection (i.p.) of 5-FU at a dose of 60 mg kg−1 per day for 5 consecutive days (1st day to 5th day) in model and drugs treatment groups. A 8 day treatment protocol (1st day to 8th day), Co H (high dose of Co, 20 mg kg−1 day), Co L (low dose of Co, 5 mg kg−1 per day), De H (high dose of De, 20 mg kg−1 per day), De L (low dose of De, 5 mg kg−1 per day), loper (4 mg kg−1 per day) and sodium carboxymethylcellulose (CMC-Na, model group, 0.2 mL/20 g per day) via the oral administration route was carried out from the same day that the mice were first given 5-FU. As control, mice were i.p. with normal saline (NS) and intragastric administration (i.g.) with CMC-Na. The experimental protocol is shown in Fig. 2. Disease severity was monitored daily by measuring food intake, water consumption, body weight and scoring the stool consistency: 0, normal (normal stool or absent); 1, slight (slightly wet and soft stool); 2, moderate (wet and unformed stool with moderate perianal staining of the coat); and 3, severe (watery stool with severe perianal staining of the coat).19,20 The incidence of diarrhea score 0 to 3 and average diarrhea score were used to evaluate the severity of diarrhea.
 |
| | Fig. 2 The experimental protocol. IM was induced in KM mice by i.p. with 5-FU at a dose of 60 mg kg−1 per day for 5 days (1st day to 5th day), then treatment with Co H (20 mg kg−1 per day), Co L (5 mg kg−1 per day), De H (20 mg kg−1 per day), De L (5 mg kg−1 per day), loper (2 mg kg−1 per day) and CMC-Na (model group, 0.2 mL/20 g per day) via the oral administration route were carried out starting from 1st day to 8th day. As control, mice in control group were i.p. with NS (0.2 mL/20 g per day) and i.g. with CMC-Na (0.2 mL/20 g per day). | |
2.4 Tissues collection
After treatment for 8 days, mice peripheral blood samples were obtained by retro-orbital puncture. Each blood sample was coagulated in the refrigerator at −4 °C, and then centrifuged at 3500 rpm for 15 min. The plasma was collected and stored at −80 °C for analysis. Aliquots were used once only for the test and were not subjected to repeated freeze–thaw cycles. The animals were finally sacrificed by cervical dislocation, and the intestinal tract from the pyloric sphincter to the rectum was dissected out and flushed with NS.
2.5 Histologic examination of intestines
Segments of duodenum, jejunum, ileum and colon were harvested and fixed in 4% paraformaldehyde for 24 h. Tissues were embedded in paraffin and 7 μm-thick cross-sections were prepared for hematoxylin and eosin (HE) staining. The sections were dewaxed, rehydrated, stained in Lillie-Mayer's haematoxylin for 10 min, and differentiated in 1% acid alcohol before being counterstained in eosin. The morphological assessment was performed by measuring villus height and crypts depth under a light microscope at a magnification of 100, which was connected to a computer with an image analysis program (Nikon NIS-Elements). The villus height was measured from the baseline to the villus tip and the crypt length was measured from the baseline to the crypt bottom. Fifteen villus and crypts in longitudinal tissue sections were counted per mouse.
2.6 Enzyme-linked immunosorbent assay (ELISA) for cytokines
Methane dicarboxylic aldehyde (MDA), superoxide dismutase (SOD), TNF-α, NF-κB, IL-10 and NO in plasma were quantified by ELISA according to the manufacturer's protocols (Enzyme-linked biological technology Co., Ltd, Shanghai, China).
2.7 Immunohistochemical analysis
COX-2, iNOS, occludin and proliferating cell nuclear antigen (PCNA) on ileum sections were performed by immunohistochemical staining as previously described.21 Paraformaldehyde-fixed and paraffin-embedded intestinal sections were dried on poly lysine-treated glass slides, which were further deparaffinized and rehydrated. Endogenous peroxidase was quenched in 3% hydrogen peroxide in methanol, and then covered with normal goat serum for 20 min, incubated with the mouse monoclonal antibodies against COX-2 (dilution of 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :100), iNOS (dilution of 1:100), occludin (dilution of 1:150) and PCNA (dilution of 1:100) (Boster Biological Technology, Wuhan, China) for overnight at 4 °C. After washing, the slides were stained with a biotin-conjugated goat-anti-mouse antibody and followed by horseradish peroxidase-conjugated streptavidin (Boster Biological Technology, Wuhan, China) for 30 min. The diaminobenzidine was used as the immunodetection substrate. Staining was observed using a light microscope. Quantification of immunohistochemical staining was processed by an Image Pro-Plus program with average optical density (AOD).22 All analyses were done in a blinded fashion by one investigator.
:100), iNOS (dilution of 1:100), occludin (dilution of 1:150) and PCNA (dilution of 1:100) (Boster Biological Technology, Wuhan, China) for overnight at 4 °C. After washing, the slides were stained with a biotin-conjugated goat-anti-mouse antibody and followed by horseradish peroxidase-conjugated streptavidin (Boster Biological Technology, Wuhan, China) for 30 min. The diaminobenzidine was used as the immunodetection substrate. Staining was observed using a light microscope. Quantification of immunohistochemical staining was processed by an Image Pro-Plus program with average optical density (AOD).22 All analyses were done in a blinded fashion by one investigator.
2.8 Statistical analysis
Data were expressed as mean ± standard error or percentage. Statistical significance between control and model group was analyzed with Student's t test and One-way analysis of variance (ANOVA) followed by Dunnett's test was used to test the effect of each treatment with respect to the model. Tests were performed using SPSS 17.0 system. The p-value less than or equal to 0.05 was considered to be statistically significant.
3 Results
3.1 Physical manifestations
Physical statuses of mice were examined during the experimental period (Fig. 3). Anepithymia, body weight loss and severe diarrhea were caused by 5-FU on 2nd day. The symptom severity was associated with increased days, when watery, even purulent and bloody stools appeared and was not recovered after 5-FU withdrawal. These could be attributed to the ongoing i.p. with 5-FU throughout the experimental early period. Co, De and loper relieved the weak physical statuses, but there were no significant difference compared to model group during the first 3–4 days (p > 0.05). In 5th day, the cessation day of the 5-FU i.p., physical manifestations induced by 5-FU were significantly ameliorated after Co and De treatment, especially for Co H and De H (p < 0.01, compared to 5-FU model group).
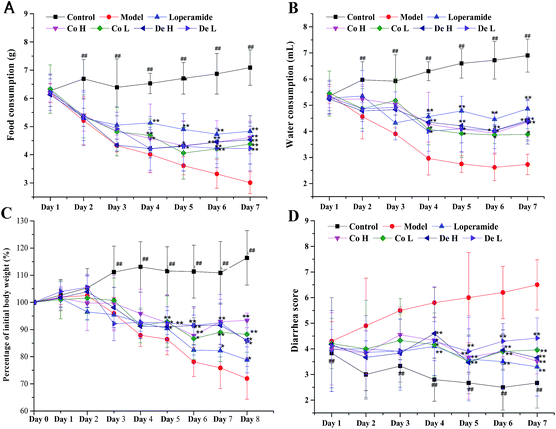 |
| | Fig. 3 The changes of physical statuses in experiment mice. (A) The index of food consumption; (B) the index of water consumption; (C) the change of body weight; (D) the diarrhea score of the experiment mice. 5-FU (60 mg kg−1 per day) were i.p. for 5 days (day 1 to 5), and loper (4 mg kg−1 per day), Co and De (5, 20 mg kg−1 per day) were i.g. from day 1 to 8. Details of experimental procedures were given in Materials and methods. Values are mean ± standard error (n = 14). *p < 0.05, **p < 0.01, compared to model group using ANOVA followed by Dunnett's test; ##p < 0.01, compared to model group using Student' t test. | |
3.2 Histopathology changes of intestines
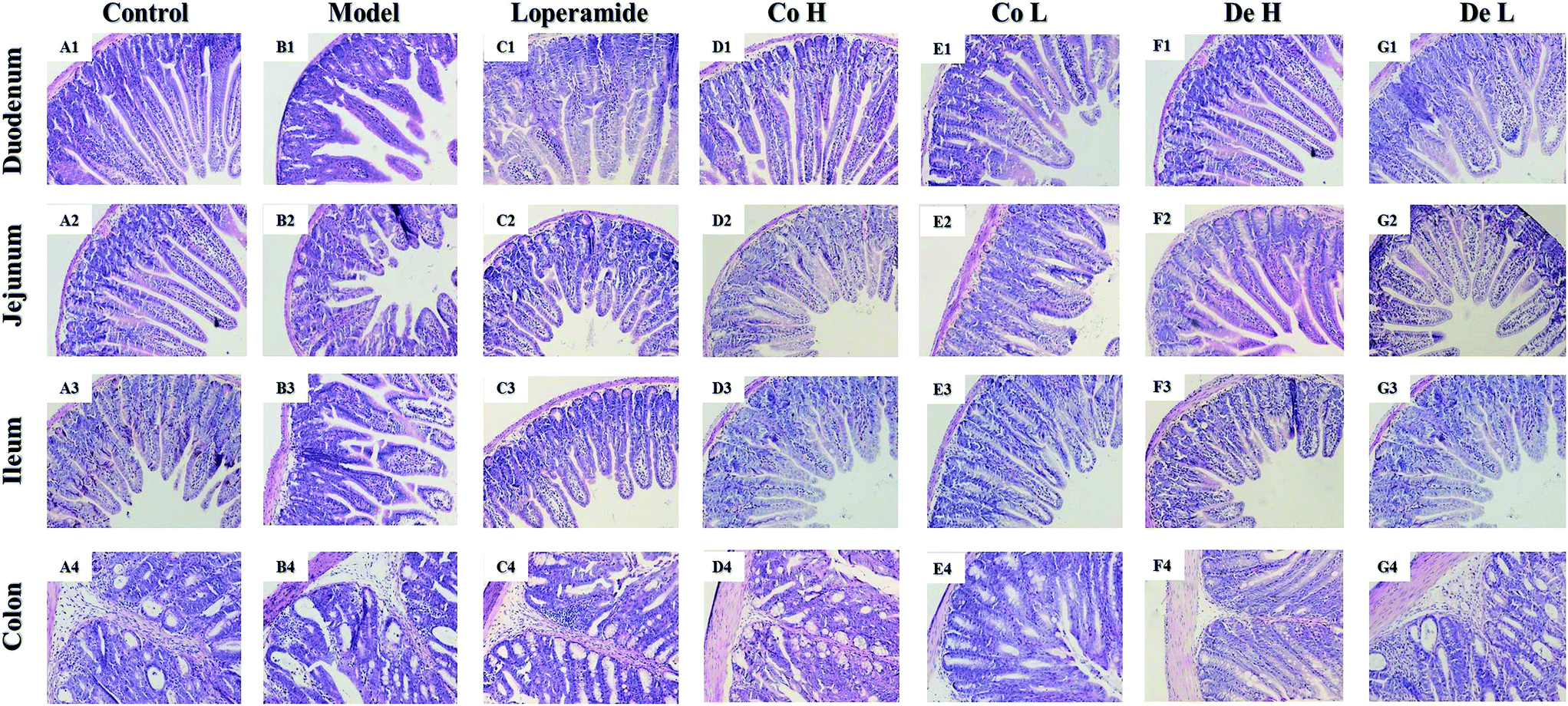
On 8th day, after killing, the intestines (duodenum, jejunum, ileum and colon) were dissected for histopathological analysis. Fig. 4 shows the morphological changes of intestines in all groups. Severely injured epithelial mucosa could be observed in small intestines, especially in ileum, after repeated administration of 5-FU (Fig. 4B1–B3), whereas significantly observable damage could rarely be found in colon section (Fig. 4B4) 5-FU induced rapid and complete loss of crypts, severe epithelial atrophy of villus, chronic inflammatory infiltration in small intestines. In contrast, ongoing Co or De treatment mitigated the morphological changes extensively characterized by distinctly longer of villi and obviously shallower of crypts (Fig. 4D1–D3, E1–E3, F1–F3 and G1–G3). Notably, intervention by Co or De showed better protective effects on 5-FU-induced morphological damage than that of loper (Fig. 4C1–C3). For further insight, the ratio of villi height/crypt depth (V/C) also verified the tendency (Table 1). Morphological analysis showed that the V/C index was shortened to −49.3% in 5-FU model mice than the control (p < 0.01). Treatment with Co or De at 5–20 mg kg−1 significantly inhibited the shortening effect induced by 5-FU in ileum (p < 0.01).
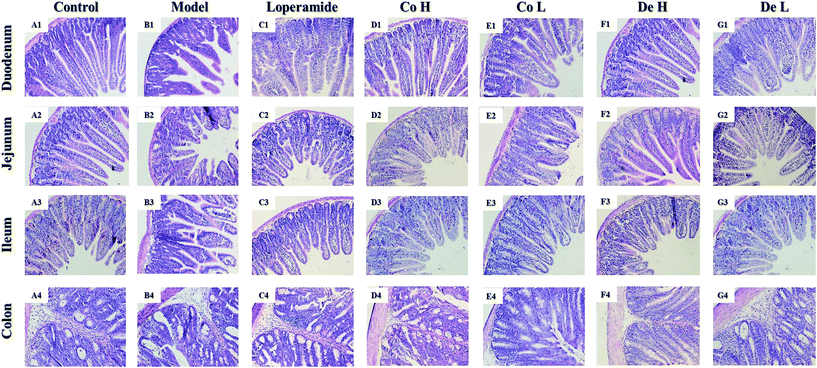 |
| | Fig. 4 Histological examination of the small intestines (duodenum, jejunum, ileum and colon). Sections were stained with HE and observed under a light microscopy. Magnification 100×. Photos were representative images (n = 6 per group). | |
Table 1 Villus height, crypts length and V/C index in ileuma
| Group |
Villus height (μm) |
Crypts length (μm) |
V/C |
| Six mice in each group were sacrificed for the morphometry analysis. Fifteen villus and crypts in longitudinal tissue sections were counted per mouse. Values are mean ± standard error (n = 6). *p < 0.05, **p < 0.01, compared to model group using ANOVA followed by Dunnett's test; ##p < 0.01, compared to model group using Student's t test. |
| Control |
59.31 ± 3.35## |
20.05 ± 3.41## |
2.96 ± 0.69## |
| Model |
36.39 ± 4.55 |
24.24 ± 2.32 |
1.50 ± 1.02 |
| Loper |
49.41 ± 1.09** |
22.04 ± 2.56** |
2.24 ± 1.29** |
| Co H |
53.71 ± 2.09** |
21.04 ± 3.68** |
2.55 ± 0.30** |
| Co L |
47.76 ± 4.01** |
20.66 ± 3.21** |
2.31 ± 0.12** |
| De H |
55.66 ± 2.33** |
21.91 ± 1.68** |
2.54 ± 0.09** |
| De L |
45.32 ± 5.12** |
19.92 ± 4.18** |
2.28 ± 0.23** |
3.3 Cytokines ELISA tests of the plasma
5-FU-induced IM is associated with the alteration of antioxidant defense mechanism through the generation of ROS.4 During the ROS process, SOD forms hydrogen peroxide through its scavenging action on the superoxide radical.23 The existing experimental evidence suggested that the 5-FU group gave a conspicuous decrease in SOD level of the plasma, and this was restored by Co or loper administration (Fig. 5A, p < 0.01, compared to the model group). De groups also presented a rising tendency in plasma SOD activity, but showed no significant difference compared to the model group. MDA, the product of phospholipid peroxidation, causes toxic stress in cells.24 We observed noticeable increasing concentration of MDA in plasma after i.p. with 5-FU for 5 days (Fig. 5B). The MDA level was alleviated after Co, De or loper treatment, especially for Co H and loper (p < 0.01, compared to the model group). De treatment showed a weaker down-regulation action in MDA level than Co, and such effect was statistically non-significant compared with model group.
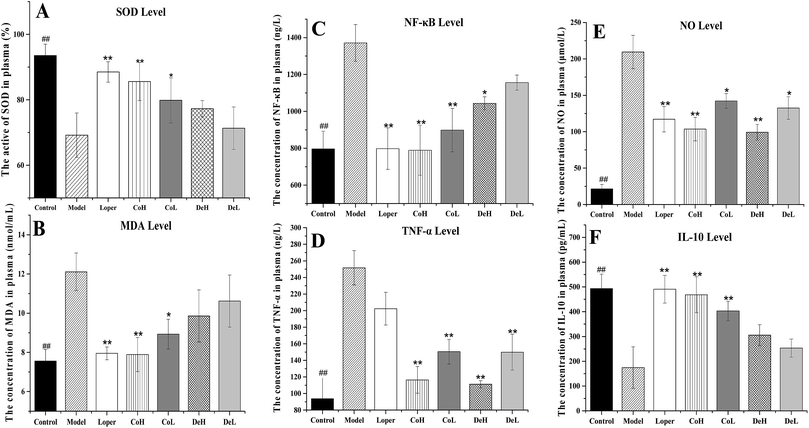 |
| | Fig. 5 Cytokines ELISA tests of the plasma experimental mice. (A) The activity of SOD in plasma; (B) the concentration of MDA in plasma; (C) the concentration of TNF-α in plasma; (D) the concentration of NF-κB in plasma; (E) the concentration of NO in plasma. (F) the concentration of IL-10 in plasma. Values are mean ± standard error (n = 10). *p < 0.05, **p < 0.01, compared to model group using ANOVA followed by Dunnett's test; ##p < 0.01, compared to model group using Student's t test. | |
As the induction and amplification of inflammatory processes likely contribute to IM,4 we further determined the effect of Co and De treatment on the expressions of relevant cytokines. Firstly, NF-κB, one of the key regulators of genes, is involved in the immune and inflammatory responses and leads to IM.25 According to the results, the level of NF-κB was obviously increased in the 5-FU model group (1371.1 ng mL−1 compared to control group 796.1 ng mL−1) (Fig. 5C). Nevertheless, Co and loper exerted inhibitory effects on NF-κB activity to ameliorate the IM induced by 5-FU (p < 0.01). Notably, De showed less potent inhibition than Co in the NF-κB activation. The concentration of NF-κB in plasma was 1043.3 ng mL−1 (p < 0.05) at De H and 1155.8 ng mL−1 (p > 0.05) at De L, respectively. Secondly, the pro-inflammatory cytokine, such as TNF-α is recognized as a mediator of activation in many systemic inflammatory reactions.26 Approximately 3-fold elevation of TNF-α expression in plasma was detected after 5-FU i.p. (Fig. 5D, 251.7 ng L−1 compared to control group 93.7 ng L−1). As expected, Co or De treatment (5, 20 mg kg−1) markedly suppressed the elevated expression of TNF-α (compared to 5-FU model group, p < 0.01), whereas loper exerted weak inhibition. Thirdly, we focus on the NO, which is a pleiotropic free radical messenger molecule. NO seems to be a key mediator of the antineoplastic drug toxicity that plays a role on the pathogenesis of IM.27 The treatment of 5-FU significantly increased the expression of NO and administration with Co, De or loper trended towards a prevention in the expansion of NO induced by 5-FU (Fig. 5E, especially for Co H, De H and loper group, p < 0.01). Last but not least, the anti-inflammatory cytokine, IL-10 was examined, and decreasing expression was observed after 5-FU treatment. On the contrary, the IL-10 expression was reversed in Co or loper-treated mice with fold changes of 490.73, 468.54 and 402.77 pg mL−1 (Co H, Co L and loper group respectively) compared to the model group (174.5 pg mL−1) (Fig. 5F). Besides, the improvement of Co on IL-10 level was superior to De after i.p. with 5-FU.
3.4 Immunohistochemistry stain of tissue
As the results shown, 5-FU increased the expressions of COX-2 and iNOS in ileum whereas decreased levels of occludin and PCNA. However, drugs treatment reversed the tendency of above factors to varying degrees (Fig. 6). COX-2 is an inducible enzyme involved in inflammation that has a key role in the production of prostaglandins from arachidonic acid.26 Not surprisingly, 5-FU-treated mice presented intense COX-2 immunostaining in ileum. The administration of Co, De and loper markedly decreased this inflammation staining (p < 0.01) (Fig. 6A1–G1). The AOD index also verified the tendency (Table 2). The iNOS expression is associated with intestinal inflammation, and the blockade of iNOS expression would protect against the exacerbation of IM by 5-FU treatment.28 The results showed that iNOS immunoexpression in ileum of mice treated with 5-FU was upregulated and, additionally reduced in drugs intervention mice (Fig. 6A2–G2).
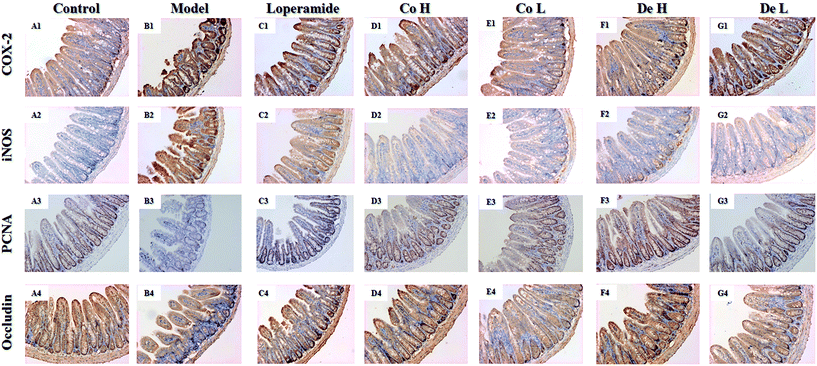 |
| | Fig. 6 Immunohistochemical staining of ileum which following 5-FU, Co, De (5, 20 mg kg−1 per day dosage) and loper (4 mg kg−1 per day dosage) administration. Sections were stained by immunohistochemistry using antibodies against (A) COX-2, (B) iNOS, (C) occludin and (D) PCNA. Magnification 100×. Photos are representative images (n = 6 per group). | |
Table 2 Quantification of immunohistochemical staining was processed by an Image Pro-Plus program with AODa
| Group |
AOD (IOD/area) |
| COX-2 |
iNOS |
PCNA |
Occludin |
| Six photos in each group were quantification analysis. Values are mean ± standard error (n = 6). *p < 0.05, **p < 0.01, compared to model group using ANOVA followed by Dunnett's test; ##p < 0.01, compared to model group using Student's t test. |
| Control |
0.072 ± 0.037## |
0.040 ± 0.021## |
0.059 ± 0.010## |
0.168 ± 0.131## |
| Model |
0.184 ± 0.033 |
0.092 ± 0.050 |
0.024 ± 0.009 |
0.073 ± 0.023 |
| Loper |
0.103 ± 0.055** |
0.061 ± 0.024* |
0.043 ± 0.010** |
0.104 ± 0.09* |
| Co H |
0.092 ± 0.037** |
0.050 ± 0.010** |
0.041 ± 0.014** |
0.120 ± 0.102** |
| Co L |
0.091 ± 0.013** |
0.056 ± 0.016** |
0.040 ± 0.011** |
0.093 ± 0.056 |
| De H |
0.087 ± 0.023** |
0.052 ± 0.020** |
0.049 ± 0.012** |
0.131 ± 0.038** |
| De L |
0.093 ± 0.014* |
0.059 ± 0.015** |
0.038 ± 0.010* |
0.099 ± 0.035** |
Especially for Co and De groups, the AOD indexes of these two groups were significantly lower than model group (p < 0.01, Table 2). PCNA is used to assess the proliferation capability of crypt cells in sections of small intestine. PCNA-positive cells were expressed in the generative zone of the control mice ileum and were strikingly decreased after treatment with 5-FU, particularly at the bottom of the crypts (p < 0.01). Conversely, all groups treated with Co, De or loper were associated with a further enhancement of PCNA expression, although uncomparable to that of control group, was much higher than the faint staining of the 5-FU group (Fig. 6A3–G3, Table 2). Occludin is the first protein identified as strand-forming component.29 It was highly expressed in the intestine and the level was significantly decreased in 5-FU-treated mice versus control mice (Fig. 6A2, p < 0.01). Co H and De treatment displayed more positive staining in the intestine as well as promoted AOD indexes compared to 5-FU-induced mice (Fig. 6A4–G4, Table 2). Loper and Co L but had only a moderate effect on elevating occludin response.
4 Discussion
5-FU is notable for significant intestine toxicities during chemotherapy and the manifestations induced by 5-FU are mainly attributed to disruptions of the architecture and integrity in the intestine.30 This destruction includes altering gut motility, mucosal flattening, epithelial cell apoptosis, inflammatory cytokines infiltration in the intestine which lead to weak physical manifestations.7,31 Administration with 5-FU for 5 consecutive days induced a severe IM which was observed as indicated by anorexia, body weight loss and diarrhea accompanied by morphological damage to the small intestine in our present study, these were consistent with results from other 5-FU-induced mucositis studies in rodents.32–35 Immediate and effective pharmacological interventions are considerable.
In our study, we presented two naturally occurring terpenes, Co and De, which were contributed to the mitigation of the severity in 5-FU-induced IM. Since Co and De are currently used in the world for the tests of various diseases such as cancer, inflammatory sickness as well as gastrointestinal modulation, these findings are interesting and relevant.36–38 Co and De have potential anti-cancer activity in many cancer models or cells. Co exerts anti-proliferative activity via forming well-organized microtubule polymers in the human breast cancer MCF-7 cells.17 De also inhibits the proliferation of A549, NCI-H460 and NCI-H520 cells by triggering endoplasmic reticulum stress, releasing cytosol-calcium levels which subsequently activate ROS, p38, JNK and ERK1/2.36 Moreover, abundant studies demonstrate that Co and De have anti-inflammatory effect on various inflammatory diseases. Activation of NF-κB in biopsies of pleurisy-model is suppressed by Co and De, suggesting that they would be potent inhibitors of NF-κB activation.38 Similarly, in human THP-1 cell, Co and De inhibit IL-6-elicited tyrosine phosphorylation of STAT3 and concomitant down-regulation of the phosphorylation of the tyrosine Janus kinases JAK1, JAK2 and Tyk2.39 Besides, the compatibility of Co and De exhibits gastrointestinal modulation, which is probably mediated through the inhibition of muscarinic receptors, 5-hydroxytryptamine receptors, and calcium influx.17 Therefore, it is likely that Co and De may be permissioned candidate drugs for ameliorating IM on the anti-tumor action of 5-FU during chemotherapy.
From the physical manifestations, anorexia and dehydration due to painful mucosal lesions were significantly alleviated by the interventions of Co, De and loper. The weight loss and severe diarrhea were acceleratedly recovered with the drugs treatment after withdrawal of 5-FU. Also, the small intestine mucosa with drugs intervention appeared to be restored with slight edema or hyperemia, under light microscopy, a relatively integral mucosal architecture was seen with aligned villus and deep crypts compared to 5-FU model mice. As above, Co and De would be potential drugs possess protective properties of the intestine that restrict the development of chemotherapy-induced IM and its related clinical symptoms. On the one hand, tight junctions (TJs) have been linked with many known hall marks of toxicity including pro-inflammatory cytokines and pathogenic bacteria.40 Occludin, a transmembrane protein plays a role in maintaining TJs integrity and regulates paracellular diffusion.41 In gut inflammatory dysregulation, down-regulation of occludin expression is associated with poor TJs integrity and increases paracellular permeability.42 In present research, decreased expression of occludin was detected in the intestine of mice by 5-FU treatment while it was reserved by Co and De. The results strengthened the hypothesis that compromised interaction of occludin was linked to intestinal barrier dysfunction, and these two naturally occurring terpenes indeed exhibited protective effects in the TJs of integral intestine. On the other hand, continuous enterocyte proliferation and the subsequent enhancement of tissue repair can attenuate gut inflammation.43 PCNA, a 36 kd protein involved in protein synthesis within intestinal epithelia, can provide an indication of deviations to normal functioning. Our results further confirmed that the notable property of Co and De was their abilities to promote the recovery of damaged intestinal tissue by increasing proliferative activity in the epithelial and crypts after 5-FU treatment.
Mechanisms involved in the pathogenesis of IM by 5-FU-challenged are the result of complex biological events and still poorly clear. Several pathogenic elements are considered to be involved, including ROS, inflammatory pathways and immune system.4 Initially, alterations are associated with the antioxidant defense mechanism through the generation of ROS that promotes cytotoxic agent exposing in intestinal mucosa, thereby resulting in cellular DNA damage and cell death.4 During the ROS process, SOD exerts scavenging action on the superoxide radical assisted by catalase to resist against ROS. In addition, MDA, one of the final products of lipid peroxidation, is used frequently to confirm the involvement of free radicals in cell damage.44 The existing results suggested that obvious degradation of SOD and apparent soar of MDA were observed after 5-FU treatment whereas revealed a pronounced recovery by administration of Co, De and loper. Moreover, the regulation of Co treatment (in particular in Co H group) on SOD and MDA expressions was similar to that of loper intervention and stronger than De administration. These results elucidated that Co and De could repress the ROS generation in 5-FU-induced IM through regulating SOD and MDA levels. The production of ROS directly triggers a cascade of inflammatory pathways and causes the tissue injury. Increasing expression of inflammatory mediators is observed, and NF-κB is thought to be pivotal in this process. Baldwin et al. confirms that NF-κB is identified in the promoter regions of several genes implicated in the pathogenesis of inflammation through consensus binding sequences.25 It has been reported that 5-FU can activate the transcription factor NF-κB in intestinal mucosa and result in up-regulating expressions of many genes, including those responsible for the production of the pro-inflammatory cytokines.45 TNF-α is acted in a positive feedback loop to reinforce NF-κB activation, and elevation of these inflammatory cytokines in plasma is usually tested during IM.46 5-FU causes over expressions of inflammatory cytokines such as NF-κB and TNF-α. These factors are associated with various metabolic disturbances and further lead to intestinal lesions.4 Interestingly, the mice of 5-FU treatment failed to induce NF-κB activity when treated with Co or loper. However, De exerted weaker inhibitive action than the former two. The release of TNF-α was effectively abrogated by treatment with Co and De but not in loper administration. Excessive iNOS expression which is triggered by activated inflammatory cytokines and further with consequent production of NO causes to generate oxidative damage.27 Ribeiro demonstrates that NO participates in the pathogenesis of oral mucositis induced by 5-FU.47 The participation of NO was supported by the increased iNOS expression in the intestinal tissue following 5-FU treatment. The variation in iNOS expression in the local intestinal tissue was in accordance with NO concentration observed in the plasma. These results indicated that they played critical roles in development of IM after 5-FU therapy. Moreover, NF-κB also increases the expression of COX-2 which is the upstream of PGE2 and regarded as the mediator of gut diarrhea and inflammation.48 Immunochemistry on mucosal tissue showed the elevation of COX-2 staining in ileum by 5-FU-challenged, whereas Co and De treatment suppressed the expression of COX-2 markedly. Besides, IL-10, an important immunoregulatory factor, is secreted by CD4+ Th2 cell, Treg, monocyte, and macrophage cells of the immune system.49 It notably contributes to decreasing the intensity of inflammatory response by inhibiting the up-stream of NF-κB transcription factor and attenuating TNF-α expression at the site of tissue damage.50 Co and De elevated the expression of IL-10 in plasma, especially for Co administration compared to the 5-FU model group. Although further studies are required, we hypothesized that 5-FU could break the inflammatory homeostasis and the protective effects of Co and De in 5-FU-treated mice could be extended to relieve inflammatory pathways (Fig. 7).
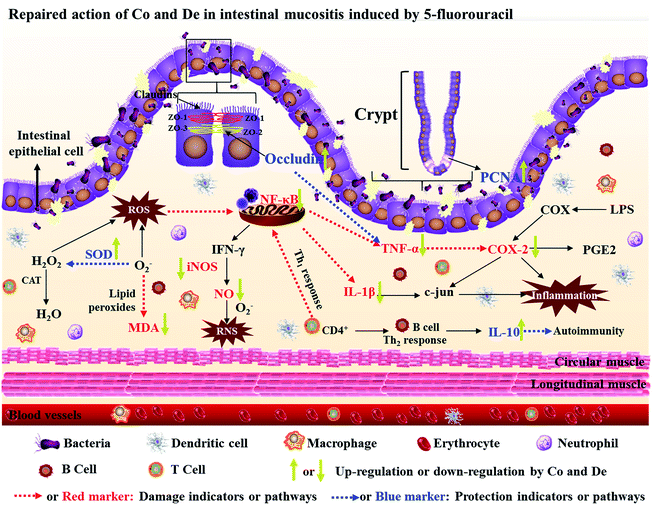 |
| | Fig. 7 The pathogenesis involved in IM and potential mechanisms protected by Co and De. | |
All above analysis strongly supported the conclusion that systemic administration of Co or De accelerated the intestinal recovery after 5-FU treatment. Collectively, we compared the cytokines regulation between these two sesquiterpene lactones and loper. As shown in Table 3, the repaired activity of Co and De are mainly focused on the oxidative stress and inflammatory reactions to varying degrees. With the paramount role in the pathogenesis of IM, the capability of Co and De to reduce poisonous effects could be promising therapeutic candidates.
Table 3 The repaired activity degrees of Co, De and loper in cytokinesa
| Factors |
Co |
De |
Loper |
| ++, exert stronger effect; +, exert strong effect; +−, exert weaker effect. |
| SOD |
++ |
+ |
++ |
| MDA |
++ |
+ |
++ |
| TNF-α |
++ |
++ |
+− |
| NF-κB |
++ |
+ |
++ |
| NO |
+ |
++ |
++ |
| IL-10 |
++ |
+ |
++ |
| iNOS |
+ |
++ |
+ |
| PCNA |
++ |
+ |
++ |
| Ocluddin |
+ |
++ |
+ |
5 Conclusions
On the whole, Co and De could reduce 5-FU induced IM through restricting related clinical symptoms, promoting the TJs, proliferating activity in the epithelial and crypts, suppressing oxidative stress and regulating the inflammatory homeostasis. Of whole supported the conclusion that Co and De could effectively ameliorate 5-FU-induced gastrointestinal toxicity.
Conflict of interest
The authors declare no competing financial interest.
Acknowledgements
The work was supported by Special Financial Grant from the China Postdoctoral Science Foundation (No. 2015T81140), PhD research startup foundation of Logistics University of Chinese People's Armed Police Forces (No. WHB201509) and China Postdoctoral Science Foundation.
References
- S. Sartori, L. Trevisani, I. Nielsen, D. Tassinari, I. Panzini and V. Abbasciano, Randomized trial of omeprazole or ranitidine versus placebo in the prevention of chemotherapy-induced gastroduodenal injury, J. Clin. Oncol., 2000, 18, 463–467 CAS.
- L. S. Elting, C. Cooksley, M. Chambers, S. B. Cantor, E. Manzullo and E. B. Rubenstein, The burdens of cancer therapy, Cancer, 2003, 98, 1531–1539 CrossRef PubMed.
- M. Duncan and G. Grant, Oral and intestinal mucositis causes and possible treatments, Aliment. Pharmacol. Ther., 2003, 18, 853–874 CrossRef CAS.
- S. T. Sonis, Pathobiology of mucositis, Semin. Oncol. Nurs., 2004, 20, 11–15 CrossRef PubMed.
- C. G. Moertel, T. R. Fleming, J. S. Macdonald, D. G. Haller, J. A. Laurie, C. M. Tangen, J. S. Ungerleider, W. A. Emerson, D. C. Tormey, J. H. Glick, M. H. Veeder and J. A. Mailliard, Fluorouracil plus levamisole as effective adjuvant therapy after resection of stage iii colon carcinoma: a final report, Ann. Intern. Med., 1995, 122, 321–326 CrossRef CAS PubMed.
- D. M. K. Keefe, J. Brealey, G. J. Goland and A. G. Cummins, Chemotherapy for cancer causes apoptosis that precedes hypoplasia in crypts of the small intestine in humans, Gut, 2000, 47, 632–637 CrossRef CAS PubMed.
- J. M. Bowen, R. J. Gibson, A. G. Cummins and D. M. K. Keefe, Intestinal mucositis: the role of the Bcl-2 family, p53 and caspases in chemotherapy-induced damage, Supportive Care in Cancer, 2006, 14, 713–731 CrossRef PubMed.
- J. Q. Gu, J. J. Gills, E. J. Park, E. Mata-Greenwood, M. E. Hawthorne, F. Axelrod, P. I. Chavez, H. H. Fong, R. G. Mehta, J. M. Pezzuto and A. D. Kinghorn, Sesquiterpenoids from Tithonia diversifolia with potential cancer chemopreventive activity, J. Nat. Prod., 2002, 65, 532–536 CrossRef CAS.
- M. Yoshikawa, S. Hatakeyama, Y. Inoue and J. Yamahara, Saussureamines A, B, C, D, and E, new anti-ulcer principles from Chinese Saussureae Radix, Chem. Pharm. Bull., 1993, 41, 214–216 CrossRef CAS PubMed.
- D. E. Wedge, J. C. G. Galindo and F. A. MacíAs, Fungicidal activity of natural and synthetic sesquiterpene lactone analogs, Phytochemistry, 2000, 53, 747–757 CrossRef CAS PubMed.
- T. H. Koo, J. H. Lee, Y. J. Park, Y. S. Hong, H. S. Kim, K. W. Kim and J. J. Lee, A sesquiterpene lactone, costunolide, from Magnolia grandiflora inhibits NF-kappa B by targeting I kappa B phosphorylation, Planta Med., 2001, 67, 103–107 CrossRef CAS PubMed.
- Y. L. Hsu, L. Y. Wu and P. L. Kuo, Dehydrocostuslactone, a medicinal plant-derived sesquiterpene lactone, induces apoptosis coupled to endoplasmic reticulum stress in liver cancer cells, J. Pharmacol. Exp. Ther., 2009, 329, 808–819 CrossRef CAS PubMed.
- P. L. Kuo, W. C. Ni, E. M. Tsai and Y. L. Hsu, Dehydrocostuslactone disrupts signal transducers and activators of transcription 3 through up-regulation of suppressor of cytokine signaling in breast cancer cells, Mol. Cancer Ther., 2009, 8, 1328–1339 CrossRef CAS PubMed.
- H. Matsuda, T. Kagerura, I. Toguchida, H. Ueda, T. Morikawa and M. Yoshikawa, Inhibitory effects of sesquiterpenes from bay leaf on nitric oxide production in lipopolysaccharide-activated macrophages: structure requirement and role of heat shock protein induction, Life Sci., 2000, 22, 2151–2157 CrossRef.
- H. J. Lee, N. Y. Kim, M. K. Jang, H. J. Son, K. M. Kim, D. H. Sohn, S. H. Lee and J. H. Ryu, A sesquiterpene, dehydrocostus lactone, inhibits the expression of inducible nitric oxide synthase and TNF-alpha in LPS-activated macrophages, Planta Med., 1999, 65, 104–108 CrossRef CAS PubMed.
- J. Cho, K. Baik and J. M. Jung, In vitro anti-inflammatory effects of cynaropicrin,
a sesquiterpene lactone, from saussurea lappa., Eur. J. Pharmacol., 2000, 398, 399–407 CrossRef CAS.
- H. M. Guo, J. Z. Zhang, W. Y. Gao, Z. Qu and C. X. Liu, Gastrointestinal effect of methanol extract of radix aucklandiae and selected active substances on the transit activity of rat isolated intestinal strips, Pharm. Biol., 2014, 52, 1141–1149 CrossRef PubMed.
- H. Mori, T. Kawamori, T. Tanaka, M. Ohnishi and J. Yamahara, Chemopreventive effect of costunolide, a constituent of oriental medicine, on azoxymethane-induced intestinal carcinogenesis in rats, Cancer Lett., 1994, 83, 171–175 CrossRef CAS PubMed.
- C. Farrell, J. Bready, K. Rex, J. Chen, C. Dipalma, K. Whitcomb, S. Yin, D. C. Hill, B. Wiemann, C. O. Starnes, A. M. Havill, Z. N. Lu, S. L. Aukerman, G. F. Pierce, A. Thomason, C. S. Potten, T. R. Ulich and D. L. Lacey, Keratinocyte growth factor protects mice from chemotherapy and radiation-induced gastrointestinal injury and mortality, Cancer Res., 1998, 58, 933–939 CAS.
- R. J. Gibson, J. M. Bowen, M. R. Inglis, A. Cummins and D. M. Keefe, Irinotecan causes severe small intestinal damage, as well as ileum ic damage, in the rat with implanted breast cancer, J. Gastroenterol. Hepatol., 2003, 18, 1095–1100 CrossRef CAS PubMed.
- R. M. Logan, A. M. Stringer, J. M. Bowen, R. J. Gibson, S. T. Sonis and D. M. K. Keefe, Is the pathobiology of chemotherapy-induced alimentary tract mucositis influenced by the type of mucotoxic drug administered?, Cancer Chemother. Pharmacol., 2009, 63, 239–251 CrossRef CAS PubMed.
- E. M. Brey, Z. Lalani, C. Johnston, M. Wong, L. V. McIntire, P. J. Duke and C. W. Patrick, Automated selection of dab-labeled tissue for immunohistochemical quantification, J. Histochem. Cytochem., 2003, 51, 575–584 CrossRef CAS PubMed.
- G. Singh, A. T. Singh, A. J. Abraham, B. Bhat, A. Mukherje, R. Verma, S. K. Agarwal, S. Jha, R. Mukherjee and A. C. Burman, Protective effects of Terminalia arjuna against Doxorubicin-induced Cardiotoxicity, J. Ethnopharmacol., 2008, 117, 123–129 CrossRef PubMed.
- E. Farmer and C. Davoine, Reactive electrophile species, Curr. Opin. Plant Biol., 2007, 10, 380–386 CrossRef CAS PubMed.
- A. S. Baldwin, The NF-kappaB and I kappa B proteins: new discoveries and insights, Annu. Rev. Immunol., 1996, 14, 649–683 CrossRef CAS PubMed.
- J. R. Vane, Y. S. Bakhle and R. M. Botting, Cyclooxygenases 1 and 2, Annu. Rev. Immunol., 1998, 38, 97–120 CAS.
- R. C. P. Lima-Júnior, A. A. Figueiredo, H. C. Freitas, M. L. P. Melo, D. V. T. Wong, C. A. V. Leite and R. A. Ribeiro, Involvement of nitric oxide on the pathogenesis of irinotecan-induced intestinal mucositis: role of cytokines on inducible nitric oxide synthase activation, Cancer Chemother. Pharmacol., 2012, 69, 931–942 CrossRef PubMed.
- K. C. Raymond and T. Keith, Nitric oxide in inflammatory bowel disease, Inflammatory Bowel Dis., 2003, 9, 179–189 CrossRef.
- M. Furuse, T. Hirase, M. Itoh, A. Nagafuchi, S. Yonemura and S. Tsukita, Occludin: a novel integral membrane protein localizing at tight junctions, J. Cell Biol., 1993, 123, 1777–1788 CrossRef CAS PubMed.
- J. Andreyev, P. Ross, C. Donnellan, E. Lennan, P. Leonard, C. Waters, L. Wedlake, J. Bridgewater, R. Glynne-Jones, W. Allum, I. Chau, R. Wilson and D. Ferry, Guidance on the management of diarrhoea during cancer chemotherapy, Lancet Oncol., 2014, 15, e447–e460 CrossRef PubMed.
- T. Y. Huang, H. C. Chu, Y. L. Lin, W. H. Ho, H. S. Hou, Y. C. Chao and L. L. Ching, Minocycline attenuates 5-fluorouracil-induced small intestinal mucositis in mouse model, Biochem. Biophys. Res. Commun., 2009, 389, 634–639 CrossRef CAS PubMed.
- T. H. Wright, R. Yazbeck, K. A. Lymn, E. J. Whitford, K. Y. Cheah, R. N. Butler, C. F. Bisset, A. N. Pilichiewicz, S. Mashtoub and G. S. Howarth, The herbal extract, iberogast, improves jejunal integrity in rats with 5-fluorouracil (5-fu)-induced mucositis, Cancer Biol. Ther., 2009, 8, 923–929 CrossRef.
- R. K. L. Leu, I. L. Brown, H. Ying, A. R. Bird, J. Michelle, E. Adrian and G. P. Young, A synbiotic combination of resistant starch and bifidobacterium lactis facilitates apoptotic deletion of carcinogen-damaged cells in rat colon, J. Nutr., 2005, 135, 996–1001 Search PubMed.
- K. L. Tooley, G. S. Howarth, K. A. Lymn, A. Lawrence and R. N. Butler, Oral ingestion of Streptococcus thermophilus diminishes severity of small intestinal mucositis in methotrexate treated rats, Cancer Biol. Ther., 2006, 5, 593–600 CrossRef CAS PubMed.
- E. J. Whitford, A. G. Cummins, R. N. Butler, L. D. Prisciandaro, J. K. Fauser, R. Yazbeck, A. Lawrence, K. Y. Cheah, T. H. Wright, K. A. Lymn and G. S. Howarth, Effects of streptococcus thermophilus th-4 on intestinal mucositis induced by the chemotherapeutic agent, 5-fluorouracil (5-fu), Cancer Biol. Ther., 2009, 8, 505–511 CrossRef CAS PubMed.
- J. Y. Hung, Y. L. Hsu, W. C. Ni, Y. M. Tsai, C. J. Yang, P. L. Kuo and M. S. Huang, Oxidative and endoplasmic reticulum stress signaling are involved in dehydrocostuslactone-mediated apoptosis in human non-small cell lung cancer cells, Lung Cancer, 2010, 68, 355–365 CrossRef PubMed.
- E. Butturini, R. D. Paola, H. Suzuki, I. Paterniti, A. Ahmad, S. Mariotto and S. Cuzzocrea, Costunolide and Dehydrocostuslactone, two natural sesquiterpene lactones, ameliorate the inflammatory process associated to experimental pleurisy in mice, Eur. J. Pharmacol., 2014, 730, 107–115 CrossRef CAS PubMed.
- C. Bocca, L. Gabriel, F. Bozzo and A. Miglietta, A sesquiterpene lactone, costunolide, interacts with microtubule protein and inhibits the growth of mcf-7 cells, Chem.-Biol. Interact., 2004, 147, 79–86 CrossRef CAS PubMed.
- E. Butturini, E. Cavalieri, A. C. de Prati, E. Darra, A. Rigo, K. Shoji, N. Murayama, H. Yamazaki, Y. Watanabe, H. Suzuki and S. Mariotto, Two Naturally Occurring Terpenes, Dehydrocostuslactone and Costunolide, Decrease Intracellular GSH Content and Inhibit STAT3 Activation, PLoS One, 2011, 6, e20174 CAS.
- R. M. Logan, A. M. Stringer, J. M. Bowen, A. S. Yeoh, R. J. Gibson, S. T. Sonis and D. M. Keefe, The role of pro-inflammatory cytokines in cancer treatment-induced alimentary tract mucositis: pathobiology, animal models and cytotoxic drugs, Cancer Treat. Rev., 2007, 33, 448–460 CrossRef CAS PubMed.
- P. J. Moughan, J. M. Wells and N. C. Roy, Regulation of tight junction permeability by intestinal bacteria and dietary components, J. Nutr., 2011, 141, 769–776 CrossRef.
- K. R. Groschwitz and S. P. Hogan, Intestinal barrier function: molecular regulation and disease pathogenesis, J. Allergy Clin. Immunol., 2009, 124, 3–20 CrossRef CAS.
- M. J. Andres, C. S. Williams, J. M. Amann, T. C. Ellis, M. M. Isabel, W. M. Kay, P. Gregoli and S. W. Hiebert, Deletion of mtgr1 sensitizes the colonic epithelium to dextran sodium sulfate-lnduced colitis, Gastroenterology, 2006, 131, 579–588 CrossRef PubMed.
- S. K. Gupta, S. Gautam, J. K. Rawat, M. Singh, S. A. Saraf and G. Kaithwas, Efficacy of variable dosage of aspirin in combating methotrexate-induced intestinal toxicity, RSC Adv., 2015, 5, 9354–9360 RSC.
- S. T. Sonis, The biologic role for nuclear factor-kappab in disease and its potential involvement in mucosal injury associated with anti-neoplastic therapy, Crit. Rev. Oral Biol. Med., 2002, 13, 380–389 Search PubMed.
- K. Shinichi, H. Shusaku, K. Yumeno, N. Koyo, A. Hitomi, S. Junichiro, U. Daichi, A. Kikuko and K. Makoto, Saireito (tj-114), a japanese traditional herbal medicine, reduces 5-fluorouracil-induced intestinal mucositis in mice by inhibiting cytokine-mediated apoptosis in intestinal crypt cells, PloS One, 2015, 10, e0116213 Search PubMed.
- R. F. C. Leitão, R. A. Ribeiro, E. A. L. Bellaguarda, F. D. B. Macedo, L. R. Silva, R. B. Oriá, M. L. Vale, F. Q. Cunha and G. A. C. Brito, Role of nitric oxide on pathogenesis of 5-fluorouracil induced experimental oral mucositis in hamster, Cancer Chemother. Pharmacol., 2007, 59, 603–612 CrossRef.
- Y. Kase, T. Hayakawa, Y. Togashi and T. Kamataki, Relevance of irinotecan hydrochloride-induced diarrhea to the level of prostaglandin E2 and water absorption of large intestine in rats, Jpn. J. Pharmacol., 1997, 75, 399–405 CrossRef CAS.
- K. Asadullah, W. Sterry and H. D. Volk, Interleukin-10 therapy–review of a new approach, Pharmacol. Rev., 2003, 55, 241–269 CrossRef CAS PubMed.
- M. Sultani, A. M. Stringer, J. M. Bowen and R. J. Gibson, Anti-inflammatory cytokines: important immunoregulatory factors contributing to chemotherapy-induced gastrointestinal mucositis, Chemother. Res. Pract., 2012, 490804 Search PubMed.
Footnote |
| † These two authors contributed equally to this work. |
|
| This journal is © The Royal Society of Chemistry 2016 |
Click here to see how this site uses Cookies. View our privacy policy here.