
Improved biomass production by humic analog anthraquinone-2-sulfonate from kitchen waste in a two-phase system†
Abstract
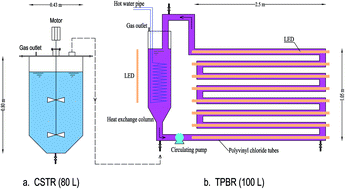
Fermentation products of volatile fatty acids from kitchen waste were explored as substrates of anoxygenic photosynthetic bacteria (APB) in a dark-photo fermentation reactor, and anthraquinone-2-sulfonate (AQS) was firstly applied in photofermentation process to boost the biomass yield. During the hydrolysis and acidification phase, the production of SCOD and volatile fatty acids (VFAs) reached 75.26 g L−1 and 14.2 g L−1 within 72 h, respectively. During the photofermentation phase, the VFAs and NH3–N removal rate in AQS test was 1.44- and 3.0-fold higher than that of control test without AQS addition, respectively. Additionally, AQS increased biomass production by 120% (0.46 g biomass/g COD) than that of control test (0.21 g biomass/g COD). Pyrosequencing results revealed that AQS lead to the highest bacterial diversity and abundance of purple non-sulfur bacteria (22.9%) and denitrifier (8.5% of Planctomyces). This study successfully established synergetic biomass production from fermentation products of kitchen waste with aid of AQS.
 Please wait while we load your content...
Please wait while we load your content...

