
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceDirect organelle thermometry with fluorescence lifetime imaging microscopy in single myotubes†
Hideki
Itoh
ab,
Satoshi
Arai
cd,
Thankiah
Sudhaharan
b,
Sung-Chan
Lee
e,
Young-Tae
Chang
fg,
Shin'ichi
Ishiwata
*cdh,
Madoka
Suzuki
*cd and
E. Birgitte
Lane
*b
aDepartment of Pure and Applied Physics, Graduate School of Advanced Science and Engineering, Waseda University, Tokyo 169-8555, Japan
bInstitute of Medical Biology, Agency for Science, Technology and Research (A*STAR), Singapore 138648, Singapore. E-mail: birgit.lane@imb.a-star.edu.sg
cOrganization for University Research Initiatives, Waseda University, Tokyo 162-0041, Japan. E-mail: suzu_mado@aoni.waseda.jp; Ishiwata@waseda.jp
dWaseda Bioscience Research Institute in Singapore (WABIOS), Singapore 138667, Singapore
eAptabio Therapeutics Inc., Yongin 446-908, Korea
fSingapore Bioimaging Consortium, Agency for Science, Technology and Research (A*STAR), Singapore 138667, Singapore
gDepartment of Chemistry and MedChem Program of Life Sciences Institute, National University of Singapore, Singapore 117543, Singapore
hDepartment of Physics, Faculty of Advanced Science and Engineering, Waseda University, Tokyo 169-8555, Japan
First published on 21st January 2016
Abstract
We describe organelle thermometry using an endoplasmic reticulum-targeting small molecule dye and cytosolic mCherry, whose fluorescence lifetimes reduce with increasing temperature and can be monitored by fluorescence lifetime imaging microscopy. The results show that heat production in single myotubes is highly localized and is coupled to a Ca2+ burst.
Optical sensors that allow us to obtain the read-out of temperature with fluorescence signals have begun to emerge recently.1 We have developed microscopic thermometry methods using glass capillaries,2 nano-particles,3–5 and sheet-coated cover glass.6 For intracellular thermometry, the precise subcellular localization of these thermosensors is all-important. Because the measured temperature is a function of the distance from the heat source, the location of the probe critically affects the accuracy of the temperature measurements. To the best of our knowledge, there have been only two sensors reported that meet the spatial requirements for targeting the endoplasmic reticulum (ER) or the sarcoplasmic reticulum (SR), which produces heat in the skeletal muscle during non-shivering thermogenesis (NST).7,8 One type of sensor is the genetically-encoded protein-based ratiometric fluorescent thermosensor which can be expressed in the organelle of interest.9 Another is a small molecule-based fluorescent thermosensor that specifically localizes to ER or SR:10 this is easier to use by virtue of its small molecular size, leading to high cell permeability, and its photostability. In this study, fluorescence lifetime imaging microscopy (FLIM) was used because of its advantages over intensity-based methods, in that the fluorescence lifetime of the dye is independent of sample displacement, dye concentration and/or light source intensity.1 Currently, two types of FLIM are available. Time-domain FLIM is the most frequently used, but requires several minutes to acquire one image with relatively high spatial resolution. Frequency-domain FLIM takes about 10 s to create a single image but with a lower spatial resolution. By using both types of FLIM, we demonstrated a subcellular-level quantitative imaging of Ca2+-induced heat production in myotubes, through the use of the ER-targeted fluorescence thermometer, ER thermo yellow (Fig. 1a), as described by us previously,10 and cytosolic mCherry.
 | ||
| Fig. 1 Properties of fluorescence lifetime of ER thermo yellow. (a) Chemical structure of ER thermo yellow. (b) Fluorescence lifetime images of living C2C12 myotubes loaded with ER thermo yellow at 25 °C and 37 °C. Scale bar, 50 μm; calibration bar, 2.3–3.3 ns. | ||
As the fluorescence intensity of ER thermo yellow is temperature sensitive,10 we first confirmed that ER thermo yellow would also work as a temperature sensor with FLIM in HeLa cells. The fluorescence lifetime of ER thermo yellow measured by time-domain FLIM shortened with temperature increase (Fig. S1, ESI†). The intensity-weighted average fluorescence lifetime (τint) of ER thermo yellow was 2.50 ns (3.31 ns, 35%; 1.64 ns, 65%) at 25 °C and 2.28 ns (3.19 ns, 29%; 1.49 ns, 71%) at 37 °C. The relationship between temperature and the average fluorescence lifetime of ER thermo yellow measured by frequency-domain FLIM (Fig. S2, ESI†) was linear (R2 = 0.97) in living HeLa cells between 23 °C and 40 °C (Table S1 and Fig. S3a, ESI†). The temperature sensitivity was −26 ps per °C and the accuracy of temperature measurements was 1.6 ± 0.4 °C (average ± standard deviation (SD)) under these experimental conditions. In ideal situations, the absolute temperature could be determined by means of FLIM. However, in our setup with frequency-domain FLIM, it was observed that the accuracy of absolute temperature measurements in this study was almost the same as the magnitude of temperature changes in response to chemical stimulations, as mentioned below. It was therefore decided to limit the analysis to the temperature difference in this study. A similar thermosensitivity with better accuracy was recorded in parallel experiments using fixed HeLa cells (Table S1 and Fig. S3b, ESI†). The effects of the chemical environment on the fluorescence lifetime of ER thermo yellow were also measured by time-domain FLIM. In a physiological pH range (5.0–8.0), the fluorescence lifetime of ER thermo yellow fluctuated, but it was relatively stable at the normal pH of ER (pH 7.4 ± 0.2);11τint = 2.18 ± 0.07 ns at pH 7 and τint = 2.16 ± 0.08 ns at pH 8 (Fig. S4a, ESI†). The emission spectra show a similar trend (Fig. S4b, ESI†). τint was independent of the ionic strength (0–500 mM KCl) (Fig. S4c, ESI†). We and others have measured a temperature increase of about 1 °C that is induced by a Ca2+ shock with ionomycin, a Ca2+ ionophore.2,5,10,12 Hence we validated the thermometry with the fluorescence lifetime of ER thermo yellow in living HeLa cells by means of ionomycin stimulation. The temperature increase after the addition of 2 μM ionomycin was 0.93 ± 0.68 °C (vehicle: 0.03 ± 0.42 °C), which was consistent with previous studies (Fig. S5, ESI†).
This thermometry can be used on C2C12 myotubes (Fig. 1b, Table S1 and Fig. S6, ESI†). The NST of the skeletal muscle is mediated by sarco/endoplasmic reticulum Ca2+-ATPase (SERCA), which is a Ca2+ pump located in the SR membrane that actively transfers Ca2+ from the cytosol to the SR lumen by means of ATP hydrolysis. The Ca2+ release through the ryanodine receptor (RyR), which is a Ca2+ channel located at SR, triggers the NST.7,8 Using frequency-domain FLIM, we evaluated the efficacy of ER thermo yellow for monitoring the heat production in C2C12 myotubes. First, we stimulated RyR with 1 mM caffeine, which increases the open probability of RyR, so that Ca2+ is released from the SR lumen.13,14 The myotubes were loaded with a Ca2+ indicator (fluo-4) to visualize intracellular Ca2+ concentration. Immediately after the addition of 1 mM caffeine, a Ca2+ burst was observed, followed by a return to basal Ca2+ concentration at about 120 s (Fig. 2a and Fig. S7, ESI†). Thapsigargin, an inhibitor of SERCA,15 also induced a Ca2+ burst when applied at 1 μM due to the Ca2+ leakage through RyR and store-operated calcium entry,16 which peaked at 60 s, and the Ca2+ signal remained high for 150 s (Fig. 2b and Fig. S7, ESI†). The fluorescence lifetime of ER thermo yellow shortened after the addition of 1 mM caffeine, and returned within 10 min (Fig. 2c and Fig. S8, ESI†), but no obvious change was observed upon 1 μM thapsigargin addition (Fig. 2d). As mentioned above (Fig. S4a and b, ESI†), the fluorescence lifetime of ER thermo yellow was changed between pH 6 and pH 7, but was constant below pH 6 and over pH 7. We observed the shortening of the fluorescence lifetime of ER thermo yellow upon 1 mM caffeine stimulation; however, this cannot be explained by the decrease of pH from pH 7.4 ± 0.2.11 The myotubes were displaced due to the Ca2+ bursts (Fig. S9, ESI†). In this situation, single-colour intensity-based methods are problematic. In contrast, Fig. 2d shows that the fluorescence lifetime of ER thermo yellow is independent of the displacement of myotubes, which is one of the advantages of FLIM. Upon 1 mM caffeine stimulation, the temperature increased after a Ca2+ burst (Fig. S10, ESI†), which was consistent with the gap between the Ca2+ burst and the temperature increase of HeLa cells in response to ionomycin in our previous study.2 The average trend showed a peak at 105 s (Fig. 2c), and the temperature changes at 105 s before (ΔTbefore) and after (ΔTafter) the addition of 1 mM caffeine were 0.07 ± 0.25 °C and 1.6 ± 0.6 °C (p = 1.7 × 10−8) respectively, whereas ΔTbefore and ΔTafter for the vehicle (DMEM) only addition were −0.14 ± 0.26 °C and −0.23 ± 0.28 °C (p = 0.36), respectively (Fig. 2e). Upon 1 μM thapsigargin stimulation, ΔTbefore (at −45 s) and ΔTafter (at 45 s) were very similar, 0.10 ± 0.28 °C and 0.38 ± 0.34 °C (p = 1.9 × 10−2) respectively; 0.01 ± 0.22 °C and −0.15 ± 0.28 °C (p = 7.5 × 10−2), respectively, in the control (DMSO) measurements (Fig. 2f). The control time course data are shown in Fig. S11, ESI†. In addition, the changes of the fluorescence lifetime of ER thermo yellow were suppressed upon 1 mM caffeine stimulation in the presence of 1 μM thapsigargin (Fig. S12, ESI†), which is consistent with other results in this study.
 | ||
| Fig. 2 A key player of heat production, SERCA, in C2C12 myotubes. (a and b) Time course of the fluorescence intensity of fluo-4 in C2C12 myotubes stimulated by 1 mM caffeine or 1 μM thapsigargin. The thin lines: individual myotubes; the thick lines: the means (a, N = 4, n = 36; b, N = 4, n = 20). (c and d) Time course of the fluorescence lifetime changes of ER thermo yellow in C2C12 myotubes stimulated by 1 mM caffeine or 1 μM thapsigargin. The thin lines: individual myotubes; the thick lines: the means (c, N = 3, n = 16; d, N = 3, n = 15). (e and f) Comparison of the ΔT of SR in C2C12 myotubes between 1 mM caffeine (red circles) and DMEM (blue circles) stimulations, or between 1 μM thapsigargin (red circles) and DMSO (blue circles) stimulations. The red and blue diamonds are the mean values in each condition, respectively. | ||
The expression levels of RyR and SERCA increase with the differentiation process of C2C12 cells.9,17 Consistently, 1 mM caffeine induced neither detectable Ca2+ burst in C2C12 myoblasts (Fig. 3a and Fig. S13, ESI†), nor the fluorescence lifetime change of ER thermo yellow (Fig. 3b). These results also confirm that caffeine itself has no significant effect on the fluorescence lifetime of ER thermo yellow. The temperature sensitivity of ER thermo yellow in myoblasts was −25 ps per °C (Table S1 and Fig. S14a, ESI†). The ΔTbefore (at −105 s) and ΔTafter (at 105 s) were 0.10 ± 0.28 °C and 0.09 ± 0.36 °C (p = 0.86), respectively (Fig. 3c). The control measurements showed essentially the same trend (Fig. S14b, ESI†); ΔTbefore (at −105 s) and ΔTafter (at 105 s) were 0.05 ± 0.35 °C and −0.10 ± 0.40 °C (p = 1.5 × 10−2), respectively (Fig. 3c). We therefore concluded that there was no substantial temperature increase of ER in single C2C12 myoblasts upon 1 mM caffeine stimulation, which could be detected using these methods.
 | ||
| Fig. 3 No substantial events in C2C12 myoblasts (undifferentiated cells) stimulated by 1 mM caffeine. (a) Time course of the fluorescence intensity of fluo-4 in C2C12 myoblast stimulated by 1 mM caffeine. The thin lines: individual myoblasts; the thick line: the mean (N = 2, n = 39). (b) Time course of the fluorescence lifetime changes of ER thermo yellow in C2C12 myoblasts stimulated by 1 mM caffeine. The thin line: individual myoblasts; the thick line: the mean (N = 3, n = 74). (c) Comparison of the ΔT of ER in C2C12 myoblasts between 1 mM caffeine (red circles) and DMEM (blue circles) stimulations. The red and blue diamonds are the mean values in each condition, respectively. | ||
We observed the Ca2+-induced temperature increase (∼1.6 °C) at the SR, a target organelle that we expected to be a major heat source, in C2C12 myotubes. We expected that the heat should diffuse out from the SR to the surrounding environment within the cell. Thus, the temperature around the SR should vary inversely with the distance from the SR. To examine the temperature increase in the cytosol, we expressed mCherry (a fluorescent protein) in the cytosol of C2C12 myotubes. Since the fluorescence quantum yield decreases with temperature increase,18 we expected that the fluorescence lifetime of mCherry would also be temperature dependent. In fact, the τint of cytosolic mCherry was 1.42 ns (1.85 ns, 35%; 1.01 ns, 65%) at 25 °C and 1.30 ns (1.77 ns, 29%; 0.93 ns, 71%) at 37 °C (Fig. 4a). These τint values were consistent with a literature value (∼1.46 ns at 20 °C).19 The relationship between temperature and the fluorescence lifetime of cytosolic mCherry was linear (R2 = 0.98) between 24 °C and 40 °C, and the temperature sensitivity was −14 ps per °C (Fig. S15, ESI†). We then measured temperature changes, by means of the fluorescence lifetime of mCherry, in the cytosol of C2C12 myotubes exposed to 1 mM caffeine (Fig. S16a, ESI†). The temperature differences at 105 s before (ΔTbefore) and 105 s after (ΔTafter) the addition of 1 mM caffeine were −0.03 ± 0.12 °C and −0.07 ± 0.18 °C (p = 0.74), respectively (Fig. 4b). In the control measurements, the ΔTbefore and ΔTafter were −0.20 ± 0.14 °C and −0.15 ± 0.20 °C (p = 0.68), respectively (Fig. 4b and Fig. S16b, ESI†). We could detect no significant changes of the fluorescence lifetime of mCherry by 1 mM caffeine in the cytosol of C2C12 myotubes.
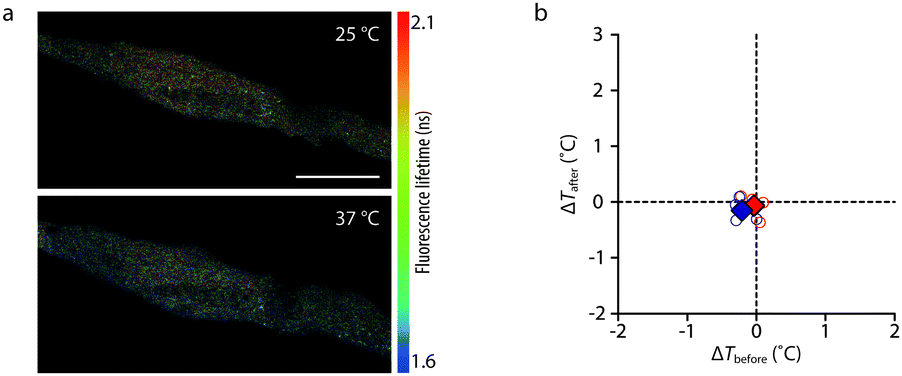 | ||
| Fig. 4 Temperature change measurements of cytosol in C2C12 myotubes. (a) Fluorescence lifetime images of living C2C12 myotubes expressing cytosolic mCherry captured at 25 °C and 37 °C. Scale bar, 50 μm; calibration bar, 1.6–2.1 ns. (b) Comparison of the ΔT of cytosol in myotubes between 1 mM caffeine (red circles) and DMEM (blue circles) stimulations. The red and blue diamonds are the mean values in each condition, respectively. | ||
Small-molecule dyes such as ER thermo yellow have advantages in handling compared to microinjection,20 transfection,9 and osmotic pressure.21,22 In comparison, the expression efficiency of mCherry in C2C12 myotubes was low, while ER thermo yellow was incorporated evenly into almost all cells (Fig. 1b). This supports the general versatility of ER thermo yellow with FLIM in a variety of cells and tissues, including primary culture cells. The temperature sensitivity of the fluorescence lifetime of ER thermo yellow was between −24 and −26 ps per °C in living cells regardless of cell types (Table S1, ESI†). The fluorescence lifetime of ER thermo yellow was independent of pH (Fig. S4a and b, ESI†) and ionic strength (Fig. S4c, ESI†) within the reported range of ER, and cytosolic Ca2+ dynamics had no obvious effect on the fluorescence lifetime of ER thermo yellow either (Fig. 2b and d). Moreover, the calibration data in living (Fig. S3a, ESI†) and fixed (Fig. S3b, ESI†) cells were quite similar to each other, strongly suggesting that the fluorescence lifetime of ER thermo yellow is independent of viscosity in cells. From these results, we conclude that the fluorescence lifetime of ER thermo yellow is most probably resistant to the environmental changes within the known physiological range.
There is an ongoing debate as to whether heat production is measurable in living cells.23–26 It has been reported that thermogenesis of brown adipocytes (1.63 ± 0.73 nW per cell) and cardiomyocytes (1.71 ± 0.94 nW per cell)27 is greater than that of other types of cells (e.g. H35 cells: 15 pW per cell),28 and cells which naturally have a thermogenic role in the body would be expected to produce more heat. Another aspect considered is the heterogeneous temperature increase in cells. No detectable temperature increase was observed in the fluorescence lifetime measurement of cytosolic mCherry of C2C12 myotubes stimulated by 1mM caffeine (Fig. 4b). This result also shows an analogy with previous studies,5,12 where heterogeneous thermogenesis was detected in HeLa and NIH3T3 cells in response to ionomycin stimulation, although their nanothermometers could not demonstrate the location specificity of the thermogenesis.
This study demonstrates, for the first time, to the best of our knowledge, Ca2+-induced heat production at the single live cell level in a C2C12 myotube model, which may partially represent NST of the skeletal muscle.7,8 Contrary to our results showing no detectable temperature changes with 1 μM thapsigargin treatment (Fig. 2d and f), Kiyonaka et al. reported that cyclopiazonic acid (CPA), an inhibitor of SERCA, induced a temperature drop of SR,9 suggesting that heat production is continuously occurring in untreated cells. Although the reason for these differences is not immediately clear, our results suggest that the substantial heat production at the SR happens only when Ca2+ bursts occur in the cytosol.
In conclusion, we have monitored heat production coupled with a Ca2+ burst in myotubes by means of frequency-domain FLIM. This SERCA-mediated heat production induced by Ca2+ release through RyR matches the physiological mechanism of NST,7,8 thus potentially leading to a deeper understanding of its mechanism.
This study was partially supported by the Biomedical Research Council of Singapore (H. I. and E. B. L.) and the Japan Society for the Promotion of Science (JSPS) KAKENHI through Grant Number 22227005 (to S. I.) and 15K05251 (to M. S.). H. I. was supported by the A*STAR Research Attachment Programme (ARAP). The authors thank the Institute of Medical Biology Microscopy Unit (IMU, Singapore) for microscope maintenance and Ms Yanyan Hou (WABIOS; Singapore) for technical support.
Notes and references
- X. Wang, O. S. Wolfbeis and R. J. Meier, Chem. Soc. Rev., 2013, 42, 7834–7869 RSC.
- M. Suzuki, V. Tseeb, K. Oyama and S. Ishiwata, Biophys. J., 2007, 92, L46–L48 CrossRef CAS PubMed.
- K. Oyama, M. Takabayashi, Y. Takei, S. Arai, S. Takeoka, S. Ishiwata and M. Suzuki, Lab Chip, 2012, 12, 1591–1593 RSC.
- S. Arai, F. Ferdinandus, S. Takeoka, S. Ishiwata, H. Sato and M. Suzuki, Analyst, 2015, 140, 7534–7539 RSC.
- Y. Takei, S. Arai, A. Murata, M. Takabayashi, K. Oyama, S. Ishiwata, S. Takeoka and M. Suzuki, ACS Nano, 2014, 8, 198–206 CrossRef CAS PubMed.
- H. Itoh, K. Oyama, M. Suzuki and S. Ishiwata, Biophysics, 2014, 10, 109–119 CrossRef CAS.
- N. C. Bal, S. K. Maurya, D. H. Sopariwala, S. K. Sahoo, S. C. Gupta, S. A. Shaikh, M. Pant, L. A. Rowland, S. A. Goonasekera, J. D. Molkentin and M. Periasamy, Nat. Med., 2012, 18, 1575–1579 CrossRef CAS PubMed.
- L. P. Kozak and M. E. Young, Nat. Med., 2012, 18, 1458–1459 CrossRef CAS PubMed.
- S. Kiyonaka, T. Kajimoto, R. Sakaguchi, D. Shinmi, M. Omatsu-Kanbe, H. Matsuura, H. Imamura, T. Yoshizaki, I. Hamachi, T. Morii and Y. Mori, Nat. Methods, 2013, 10, 1232–1238 CrossRef CAS PubMed.
- S. Arai, S.-C. Lee, D. Zhai, M. Suzuki and Y.-T. Chang, Sci. Rep., 2014, 4, 6701 CrossRef CAS PubMed.
- M. M. Wu, M. Grabe, S. Adams, R. Y. Tsien, H. P. H. Moore and T. E. Machen, J. Biol. Chem., 2001, 276, 33027–33035 CrossRef CAS PubMed.
- J. M. Yang, H. Yang and L. Lin, ACS Nano, 2011, 5, 5067–5071 CrossRef CAS PubMed.
- F. Protasi, A. Shtifman, F. J. Julian and P. D. Allen, Am. J. Physiol.: Cell Physiol., 2004, 286, C662–C670 CrossRef CAS PubMed.
- H. Kong, P. P. Jones, A. Koop, L. Zhang, H. J. Duff and S. R. W. Chen, Biochem. J., 2008, 414, 441–452 CrossRef CAS PubMed.
- A. Wrzosek, H. Schneider, S. Grueninger and M. Chiesi, Cell Calcium, 1992, 13, 281–292 CrossRef CAS PubMed.
- Y. Gutierrez-Martin, F. J. Martin-Romero and F. Henao, Biochim. Biophys. Acta, 2005, 1711, 33–40 CrossRef CAS PubMed.
- P. Tarroni, D. Rossi, A. Conti and V. Sorrentino, J. Biol. Chem., 1997, 272, 19808–19813 CrossRef CAS PubMed.
- A. Dhar, K. Girdhar, D. Singh, H. Gelman, S. Ebbinghaus and M. Gruebele, Biophys. J., 2011, 101, 421–430 CrossRef CAS PubMed.
- B. Seefeldt, R. Kasper, T. Seidel, P. Tinnefeld, K. J. Dietz, M. Heilemann and M. Sauer, J. Biophotonics, 2008, 1, 74–82 CrossRef CAS PubMed.
- K. Okabe, N. Inada, C. Gota, Y. Harada, T. Funatsu and S. Uchiyama, Nat. Commun., 2012, 3, 705 CrossRef PubMed.
- T. Tsuji, S. Yoshida, A. Yoshida and S. Uchiyama, Anal. Chem., 2013, 85, 9815–9823 CrossRef CAS PubMed.
- T. Hayashi, N. Fukuda, S. Uchiyama and N. Inada, PLoS One, 2015, 10, e0117677 Search PubMed.
- G. Baffou, H. Rigneault, D. Marguet and L. Jullien, Nat. Methods, 2014, 11, 899–901 CrossRef CAS PubMed.
- S. Kiyonaka, R. Sakaguchi, I. Hamachi, T. Morii, T. Yoshizaki and Y. Mori, Nat. Methods, 2015, 12, 801–802 CrossRef CAS PubMed.
- M. Suzuki, V. Zeeb, S. Arai, K. Oyama and S. Ishiwata, Nat. Methods, 2015, 12, 802–803 CrossRef CAS PubMed.
- G. Baffou, H. Rigneault, D. Marguet and L. Jullien, Nat. Methods, 2015, 12, 803 CrossRef CAS PubMed.
- E. A. Johannessen, J. M. R. Weaver, L. Bourova, P. Svoboda, P. H. Cobbold and J. M. Cooper, Anal. Chem., 2002, 74, 2190–2197 CrossRef CAS PubMed.
- C. Loesberg, J. C. Van Miltenburg and R. Van Wijk, J. Therm. Biol., 1982, 7, 87–90 CrossRef.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c5cc09943a |
| This journal is © The Royal Society of Chemistry 2016 |