
Hierarchical assembly of enzyme-inorganic composite materials with extremely high enzyme activity†
Abstract
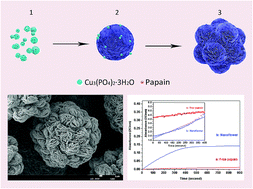
We have synthesized a novel composite material with a hierarchical flower-like structure and extremely high enzyme activity. The spherical, hierarchical flower-like structure with numerous small flowers was formed through self-assembly using papain as the organic component and Cu3(PO4)2·3H2O as the inorganic component. This hybrid-nanoflower structure was confirmed by Fourier transform infrared spectroscopy, and X-ray diffraction. The enzyme activity of papain embedded in the hybrid nanoflowers was further evaluated using BAEE as a substrate. Compared with free papain in solution, the hybrid materials exhibited extremely high enzyme activity (∼4510%). A study on the relationship of enzyme weight percentage and pattern structure with enzyme activity revealed that enzyme activity was mainly affected by the material structure. These results demonstrate that this hierarchical structure can effectively increase enzyme activity.
 Please wait while we load your content...
Please wait while we load your content...

