
Novel bivalent spermine-based neutral neogalactolipids for modular gene delivery systems†
Abstract
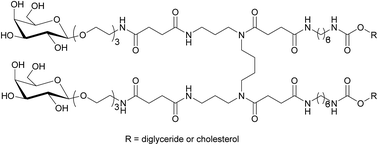
New bivalent spermine-based neutral neogalactolipids have been synthesized to develop effective modular gene delivery systems (MGDS) targeting hepatocyte asialoglycoprotein receptors. MGDS composed of new neogalactolipids, cationic gemini-amphiphile, and DOPE were agglutinated in the presence of lectin RCA120 and provided efficient delivery of fluorescently-labeled oligodeoxyribonucleotides into HepG2 cells.
 Please wait while we load your content...
Please wait while we load your content...

