DOI:
10.1039/C5RA15301H
(Paper)
RSC Adv., 2015,
5, 79874-79881
Molecular interaction of inorganic mercury(II) with catalase: a spectroscopic study in combination with molecular docking†
Received
31st July 2015
, Accepted 14th September 2015
First published on 14th September 2015
Abstract
The interaction between inorganic mercury(II) (Hg(II)) and catalase (CAT) was investigated using fluorescence, UV-visible absorption (UV-vis), circular dichroism (CD) spectroscopic techniques and molecular docking methods under simulated physiological conditions (in Tris–HCl buffer, pH = 7.40). The fluorescence quenching analysis showed that the intrinsic fluorescence of CAT was quenched by Hg(II) through a static quenching mechanism. Hg(II) can bind with CAT to form a Hg(II)–CAT complex, with a binding constant of 13.24 L mol−1 at 295 K. Thermodynamic analysis indicated that electrostatic force and van der Waals forces were the dominant intermolecular forces in stabilizing the complex. The results of UV-vis absorption and CD spectral analysis indicated that the formation of the Hg(II)–CAT complex induced some conformational changes in CAT, increasing and decreasing its α-helical content at low and high concentrations of Hg(II), respectively. The CAT activity can be inhibited by Hg(II) significantly, about a 67.2% drop with the presence of 5.0 × 10−4 mol L−1 Hg(II), and the relative activity values of CAT showed a good linear relationship with its fluorescence intensity. Molecular docking was employed to further investigate the interaction of CAT with different species of Hg(II) (HgCl2, [HgCl3]− and [HgCl4]2−), to seek the optimum binding sites of Hg(II) in CAT, and to obtain detailed binding information. This study contributes to the understanding of the interaction mechanism between Hg(II) and CAT at the molecular level in vitro, which is helpful for clarifying the toxicity mechanism of Hg(II) on an antioxidant enzyme system in vivo.
1. Introduction
Catalase (CAT, EC 1.11.1.6) is one of the most important proteins of the antioxidant defense system in plant and animal tissues, which can catalyze the decomposition of hydrogen peroxide into molecular oxygen and water.1,2 Recently, many studies showed that pathological states such as diabetes, aging, oxidative stress, and cancer were correlated with the denaturation of CAT.3,4 Meanwhile, the intake of contaminants is likely to influence the catalytic activity of CAT in tissues.5 Though some studies on the interactions between CAT and contaminants in vitro have been performed,6,7 the toxicity mechanism of some important environmental pollutants on CAT is far from being fully understood. Hence, we had paid close attention to the molecular toxicity of persistent toxic substance (such as heavy metal) on CAT. To understand the toxicity mechanism, we should make it clear that how the pollutant bond to CAT, also the structural changes and activity inhibition of CAT induced by the pollutant.
Mercury is one of the most toxic heavy metals presenting a serious threat with respect to polluting the environment and damaging human health.8,9 It is known that catalase in human red blood cells is responsible for the oxidation of elemental mercury to divalent mercury.10 However, inorganic mercury salts, especially mercury(II) (Hg(II)) salts, are more toxic than elemental mercury due to their greater water solubility.11 Both acute and chronic exposure to Hg(II) may cause damage to organs, including the lungs, kidneys, brain and liver.12 Furthermore, mercuric chloride is the most common form of Hg(II) compounds in nature.8 Mercuric chloride intoxication can cause a significant depletion of liver catalase (CAT) activity in mice.13 Hg(II) can also induce oxidative stress and make a significant contribution to the molecular mechanism for liver injury.14 Durak et al. reported that mercuric chloride can induce oxidative stress in erythrocytes through the generation of free radicals and alteration of the cellular antioxidant defense system.15 However, as these reports only focused on the effect of Hg(II) on CAT activity in vivo, little work has focused on the interaction mechanisms between Hg(II) and CAT at the molecular level. Dai et al. studied the interaction between mercuric chloride and bovine serum albumin by spectroscopic methods at the molecular level; the binding parameters and the effect of mercuric chloride on the conformation of bovine serum albumin were investigated.9 However, the mercuric chloride in Dai's experimental system actually existed as different species, such as HgCl2, [HgCl3]−, [HgCl4]2− and so on. The assay methodology within their report could not distinguish between these distinct Hg(II) species. To address this underlying issue, we proposed the use of molecular docking to study the binding interaction of different Hg(II) species with CAT.
In brief, we aimed to use spectroscopic methods combined with molecular docking to study the interaction mechanism of Hg(II) with CAT in vitro, obtain the binding parameters (binding constants, number of binding sites, thermodynamic parameters and binding forces) of the interaction and the effect of Hg(II) on the conformation of CAT, and distinguish between the interactions of CAT with different species of Hg(II). By this study, we are hoping to further understand the mechanism of the toxicity of Hg(II) with respect to CAT at the molecular level.
2. Materials and methods
2.1 Materials
Catalase (from bovine liver) was provided by Sigma Chemical Company, USA. Mercuric chloride of 99.5% purity was purchased from Guizhou Tongren Chemical Reagent Factory, China. H2O2 (30%) was purchased from Xilong Chemical Company, Ltd. Tris–HCl buffer (0.05 mol L−1, containing 0.10 mol L−1 NaCl) was used to maintain the pH of the solution at 7.40. The CAT stock solution (5.0 × 10−5 mol L−1) was prepared by dissolving CAT in Tris–HCl buffer. The mercuric chloride stock solution (1.0 × 10−2 mol L−1) was prepared by dissolving mercuric chloride in Tris–HCl buffer. All chemicals were of analytical reagent grade, and Milli-Q water was used throughout the study.
2.2 Fluorescence spectra measurements
The fluorescence measurements were carried out on a Cary Eclipse fluorescence spectrophotometer (Varian, USA). The excitation and emission slit widths were set to 5 nm and 10 nm, respectively. The excitation wavelength was set at 280 nm, and the emission scans ranged from 300 to 400 nm. The excitation synchronous fluorescence spectra were scanned from 260 to 310 nm (Δλ = 15 nm) and from 250 to 310 nm (Δλ = 60 nm).
2.3 UV-vis absorption measurements
The UV-vis absorption spectra were measured from 200 to 500 nm at room temperature (295 K) on an Agilent 8453 UV-visible spectroscopy system (Agilent Technologies, USA).
2.4 CD spectra measurements
CD spectra were recorded from 200 to 250 nm at a scan rate of 500 nm min−1 with a JASCO-810 spectrometer (Shimadzu, Japan). Three scans were measured and averaged for each CD spectrum. All of the observed spectra were baseline corrected by subtracting the spectrum of the buffer solution.
2.5 CAT activity determination
The activity of CAT was measured by monitoring the decrease in the absorbance values at 240 nm, resulting from the consumption of H2O2. The relative activity of CAT was calculated by the equation ΔA1/ΔA0 × 100%, where ΔA1 and ΔA0 are the reduction of the absorption values at 240 nm in a 2 min interval after the addition of CAT, with or without the presence of Hg(II), respectively.
2.6 Molecular docking study
Docking calculations were carried out with AutoDock 4.2 and the AutoDock Tools (ADT) software based on the method by Xu et al.16 The crystal structure of CAT was retrieved from the Protein Data Bank (http://www.rcsb.org/pdb/home/home.do, code: 1TGU). The 3D structure of ligand was generated by GaussView 5.08, and optimized by DFT/B3LYP method combined with LANL2DZ basis set using Gaussian 09 package. To reorganize the binding sites of the ligands in CAT, blind docking was carried out by setting the grid box size to 126, 126 and 126 Å along the X, Y and Z axes, with a 0.375 Å grid spacing. The Lamarckian genetic algorithm method was applied for docking simulations. The number of genetic algorithm runs, the population size and the maximum number of energy evaluations were set to 10, 150 and 250![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000, respectively. For each docking case, the lowest energy docked conformation was selected as the binding mode. Then, the docked conformations were visualized using the PyMOL software package.17
000, respectively. For each docking case, the lowest energy docked conformation was selected as the binding mode. Then, the docked conformations were visualized using the PyMOL software package.17
3. Results and discussion
3.1 Effect of Hg(II) on CAT fluorescence
The intrinsic fluorescence of CAT arises mainly from its tryptophan (Trp), tyrosine (Tyr), and phenylalanine (Phe) residues.18 Fig. 1 shows the fluorescence emission spectra of CAT with the presence of varying concentrations of Hg(II). As observed from Fig. 1, pure CAT displays a strong fluorescence emission peak at 350 nm when excited at 280 nm, while the emission fluorescence of Hg(II) can be ignored between 300 and 400 nm with identical excitation. Moreover, the fluorescence intensity of CAT decreased with the addition of Hg(II), which indicated that the fluorescence of CAT could be quenched by Hg(II).19
 |
| | Fig. 1 Fluorescence quenching spectra of CAT in the presence of various amounts of Hg(II) (pH = 7.40). c(CAT) = 1.0 × 10−6 mol L−1;104 c(Hg(II))/(mol L−1), (a–f): 0, 1.0, 2.0, 3.0, 4.0, 5.0; (g): 5.0 × 10−4 mol L−1 Hg(II) only. | |
3.2 Fluorescence quenching mechanisms
Fluorescence quenching can be caused by a dynamic or static quenching process.16,20 To interpret the quenching mechanism of Hg(II) with CAT, the fluorescence quenching spectra of CAT in the presence of various concentrations of Hg(II) were measured at three temperatures (295 K, 305 K and 315 K), and the fluorescence intensity data were analyzed by the modified Stern–Volmer eqn (S1).†19,21
The plots of F0/(F0 − F) versus [Q]−1 are shown in Fig. 2, and the values of Ksv (Table 1) can be calculated from the values of the slope. The quenching rate constant of the biomolecule Kq was evaluated using the equation Kq = Ksv/τ0. The average lifetime (τ0) of a biopolymer has been reported as 10−8 s.22 It can be observed in Fig. 2 and Table 1 that the Ksv values decrease at higher temperature and that the Kq is greater than 2.0 × 1010 L mol−1 s−1 (the maximum dynamic quenching constant of the various quenchers).23 These results indicated that the fluorescence quenching induced by Hg(II) was initiated by the formation of the Hg(II)–CAT complex.6,24
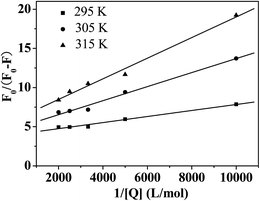 |
| | Fig. 2 Modified Stern–Volmer plots of the Hg(II)–CAT system at three temperatures. | |
Table 1 Modified Stern–Volmer quenching constants and the correlation coefficient at different temperatures
| T (K) |
Ksv (L mol−1) |
kq (L mol−1 s−1) |
R2 |
| 295 |
1.03 × 104 |
1.03 × 1012 |
0.992 |
| 305 |
5.29 × 103 |
5.29 × 1011 |
0.994 |
| 315 |
4.43 × 103 |
4.43 × 1011 |
0.994 |
To further clarify the fluorescence quenching mechanisms, the fluorescence lifetimes of the Hg(II)–CAT system were measured, and the results are shown in Table 2. The data were found to fit well to the double-exponential decay model with χ2 values close to 1.00. With the addition of Hg(II), the average lifetimes (τAV) of CAT scarcely changed. These observations further demonstrated that the quenching of CAT by Hg(II) mainly followed a static mode, which was consistent with the result from the modified Stern–Volmer equation.7
Table 2 Fluorescence lifetimes of CAT in the presence of different concentrations of Hg(II)
| Molar ratio of CAT to Hg(II) |
τAV |
χ2 |
| 1:0 |
4.44 |
1.043 |
| 1:200 |
4.47 |
0.952 |
| 1:300 |
4.51 |
1.013 |
| 1:400 |
4.50 |
0.962 |
| 1:500 |
4.49 |
1.046 |
3.3 Binding constant (Kb) and number of binding sites (n)
For the static quenching interaction, the Kb and n values can be obtained from the double logarithm eqn (S2).†25–27 The calculated Kb and n values at different temperatures are shown in Table 3. The results showed that the binding constants of the Hg(II)–CAT complex were 13.24, 8.90 and 8.04 L mol−1 at 295 K, 305 K and 315 K, respectively, with the numbers of binding sites all approaching 0.5. The binding constants decreased at higher temperatures, which indicated that the formation of the Hg(II)–CAT complex was hindered at higher temperatures.28
Table 3 Binding constants (Kb) and binding sites (n) of the Hg(II)–CAT interaction
| T (K) |
Kb (L moL−1) |
n |
R2 |
| 295 |
13.24 |
0.490 |
0.983 |
| 305 |
8.90 |
0.508 |
0.973 |
| 315 |
8.04 |
0.535 |
0.983 |
3.4 Determination of the binding forces
To determine the binding forces between Hg(II) and CAT, the thermodynamic analyses were performed based on Ross and Subramanian's theory.29 As the temperature variation range was not too wide (from 295 K to 315 K), the interaction enthalpy change (ΔH) can be regarded as a constant.24 The thermodynamic parameters (the free energy change (ΔG), ΔH and the entropy change (ΔS)) can be calculated by the Van't Hoff equation and the thermodynamic eqn (S3).†30,31
The ΔH and ΔS values were calculated from the slope and intercept values of the plot of lnK versus 1/T (Fig. 3), respectively. The thermodynamic parameters of the Hg(II)–CAT system are shown in Table 4. As Table 4 indicates, the values of ΔG at three temperatures were all negative, which indicated that the binding process of Hg(II) with CAT was spontaneous.1 Furthermore, because the ΔH (−19.38 kJ mol−1) and ΔS (−44.57 J mol−1 K−1) values were all negative in the binding reaction, the reaction was enthalpy driven, revealing that hydrogen bonds or van der Waals forces played major roles in the formation of the Hg(II)–CAT complex.9,32 However, from the structure of the main species of Hg(II) in our experimental system (S4), hydrogen bonds cannot be formed between Hg(II) and CAT. Furthermore, as the isoelectric point of CAT is 5.4,33 it should have a negative charge in the neutral pH (7.40) environment. Therefore, the electrostatic force should not be negligible between the negatively charged CAT and the charged species of Hg(II). Hence, it can be concluded that electrostatic force and van der Waals forces both played important roles in the binding reaction.
 |
| | Fig. 3 Van't Hoff plot for the interaction between Hg(II) and CAT. | |
Table 4 Thermodynamic parameters of the Hg(II)–CAT interaction
| T (K) |
ΔG (kJ mol−1) |
ΔS (J mol−1 K−1) |
ΔH (kJ mol−1) |
| 295 |
−6.23 |
−44.57 |
−19.38 |
| 305 |
−5.78 |
| 315 |
−5.33 |
3.5 Investigation on the conformational changes in CAT
Though it was confirmed that the binding of Hg(II) to CAT caused the fluorescence quenching of CAT, it was still unknown whether the binding may affect the conformation and/or micro-environment of CAT. To further evaluate this, UV-vis absorption, synchronous fluorescence, and CD spectroscopy were employed.
3.5.1 UV-vis absorption spectroscopy. As a simple but effective method, UV-vis absorption spectroscopy can be used to explore the structural changes in CAT.5,34 Fig. 4 shows the UV-vis absorption spectra of the Hg(II) and CAT mixtures (curves c and d), CAT (curve e), and different concentrations of Hg(II) (curves a and b). Fig. 4(A) illustrates that the absorption bands of Hg(II) and CAT overlap strongly at approximately 213 nm, which reflects the framework conformation of CAT.35,36 So, the subtraction spectra (curves f and g) in Fig. 4(B) were obtained by deducting the spectra of Hg(II) from the spectra of the mixed Hg(II) and CAT. It can be observed from Fig. 4(B) that the absorption peak at 213 nm decreases with the addition of Hg(II), indicating that the interaction between Hg(II) and CAT leads to the loosening and unfolding of the CAT skeleton.37,38 Furthermore, the weak absorption bands around 280 nm and 405 nm were nearly unchanged, which demonstrated that the binding of Hg(II) to CAT did not drastically change the microenvironment around the tryptophan residues and the porphyrin ring of the heme.16,39
 |
| | Fig. 4 UV-vis absorption spectra of CAT in the absence and presence of different concentrations of Hg(II) (pH = 7.40). (A) Curve a: 1.0 × 10−4 mol L−1 Hg(II); curve b: 2.0 × 10−4 mol L−1 Hg(II); curve c: 1.0 × 10−6 mol L−1 CAT + 1.0 × 10−4 mol L−1 Hg(II); curve d: 1.0 × 10−6 mol L−1 CAT + 2.0 × 10−4 mol L−1 Hg(II). (B) Curve e: 1.0 × 10−6 mol L−1 CAT; curve f: [1.0 × 10−6 mol L−1 CAT + 1.0 × 10−4 mol L−1 Hg(II)] − [1.0 × 10−4 mol L−1 Hg(II)]; curve g: [1.0 × 10−6 mol L−1 CAT + 2.0 × 10−4 mol L−1 Hg(II)] − [2.0 × 10−4 mol L−1 Hg(II)]. | |
3.5.2 Synchronous fluorescence spectroscopy. Synchronous fluorescence spectroscopy was further utilized to study the microenvironment changes in CAT induced by Hg(II) based on the possible shift in the maximum excitation wavelength.40 When the wavelength intervals (Δλ) were set as 15 nm or 60 nm, the synchronous fluorescence spectra of CAT characterized the polarity changes of the tyrosine (Tyr) or tryptophan (Trp) residues of CAT, respectively.41 Fig. 5 shows the synchronous fluorescence spectra of CAT in the presence of various amounts of Hg(II). As illustrated by Fig. 5, the synchronous fluorescence intensity of Tyr and Trp both decreased with the addition of Hg(II), and the emission peaks showed no shift over the investigated concentration range. This finding indicated that Hg(II) had no obvious effect on the microenvironment of the Tyr and Trp residues in CAT,42 which was in good agreement with the conclusions drawn from the UV-vis absorption spectral analysis.
 |
| | Fig. 5 Synchronous fluorescence spectra of the Hg(II)–CAT system. (A) Δλ = 15 nm; (B) Δλ = 60 nm. c(CAT) = 1.0 × 10−6 mol L−1;104 c(Hg(II))/(mol L−1), (a–f): 0, 1.0, 2.0, 3.0, 4.0, 5.0. | |
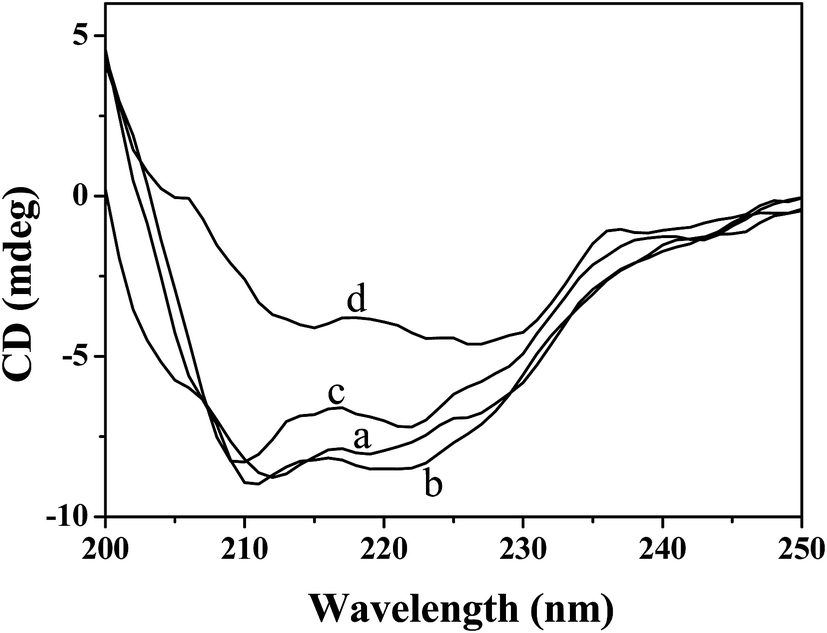
3.5.3 CD spectroscopy. To further understand the influence of Hg(II) on the secondary structure of CAT, CD spectroscopy was used. The CD spectra of CAT in the absence and presence of Hg(II) are shown in Fig. 6. As Fig. 6 indicates, the CD spectrum of pure CAT contains two main negative bands at approximately 211.0 and 219.0 nm, which are characteristic of the α-helical structure of the protein.43 It was also found that with the addition of Hg(II), the ellipticity of CAT changed significantly. Furthermore, the CDPro software package was employed to analyze the CD spectra, and the proportions of four secondary structures of CAT were obtained (Table 5). Table 5 shows that the secondary structures of pure CAT consist of 17.6% α-helix, 28.9% β-sheet, 24.1% β-turn and 24.4% random coil. After the addition of small amounts of Hg(II) to CAT (10:1, 100:1), the α-helix content of CAT increased to 23.6% and 24.7%, and the β-sheet content decreased to 27.4% and 23.6%. It was possible that the charged Hg(II) bonded with the surface charges of the CAT, which enhanced the helical structure by the dipole–dipole interaction between or within the CAT.44 However, when the molar ratio of Hg(II) to CAT increased to 1000, the α-helix content decreased to 4.9% and the β-sheet content increased to 38.7% rapidly, which may be because Hg(II) conjugated with certain amino acid residues within the CAT and therefore destroyed its hydrogen bonding networks.9 Meanwhile, the decrease in α-helix content indicated that a high concentration of Hg(II) can cause unfolding of the CAT skeleton,45 which was in good agreement with the results of the absorption study.
 |
| | Fig. 6 Effects of Hg(II) on the CD spectra of CAT. c(CAT) = 5.0 × 10−8 mol L−1;108 c(Hg(II))/(mol L−1), (a–d): 0.0, 10.0, 100.0, 1000.0. | |
Table 5 CAT secondary structure contents in the presence of different molar ratios of Hg(II)
| Molar ratio of Hg(II) to CAT |
Secondary structural elements in CAT |
| α-Helix (%) |
β-Sheet (%) |
β-Turn (%) |
Random coil (%) |
| 0:1 |
17.6 |
28.9 |
24.1 |
29.7 |
| 10:1 |
23.6 |
27.4 |
22.2 |
26.9 |
| 100:1 |
24.7 |
23.6 |
20.8 |
30.4 |
| 1000:1 |
4.9 |
38.7 |
26.8 |
28.6 |
3.6 Effects of Hg(II) on CAT activity
It is well known that the structural change of a protein is closely related to its biological function.46,47 According to the above results, the addition of Hg(II) changed the conformation of CAT remarkably, but the relevant activity changes were still unknown. Hence, the effects of different concentrations of Hg(II) on the activity of CAT were investigated. As shown in Fig. 7, the CAT activity decreased rapidly with Hg(II) concentration increasing from 0 to 5.0 × 10−4 mol L−1. As the Hg(II) concentration increased to 5.0 × 10−4 mol L−1, the CAT activity decreased to a minimum of approximately 32.8% of the initial level. This result suggested that the activity of CAT decreased in the presence of Hg(II), which may be caused by the conformational changes, as reported previously for a graphene oxide and CAT system.16 Meanwhile, it was likely that Hg(II) would have an acute toxic effect on an antioxidant enzyme system in organisms.
 |
| | Fig. 7 Effects of different concentrations of Hg(II) on the activity of CAT (pH = 7.40). c(CAT) = 1.0 × 10−6 mol L−1; 104 c(Hg(II))/(mol L−1): 0, 1.0, 2.0, 3.0, 4.0, 5.0. | |
3.7 Correlativity of CAT activity and CAT fluorescence intensity
Both the CAT activity and the CAT fluorescence intensity decreased significantly with the addition of Hg(II), and it was interesting to determine their inter-dependent relation within this study. Fig. 8 demonstrates a clear linear relation between the CAT activity and the CAT fluorescence intensity. The linear regression equation was determined to be Y = 1.36X − 248.09, and the correlation coefficient (r) was 0.9453. This phenomenon may be ascribed to the fact that both the CAT activity and CAT fluorescence intensity changed mainly as a result of the conformational changes of CAT.
 |
| | Fig. 8 Linear relationship between the CAT activity and CAT fluorescence intensity. | |
3.8 The species of Hg(II) in the experimental system
It is well known that Hg2+ shows a strong trend to form coordination complexes with chloride ion (such as [HgCl]+, HgCl2, [HgCl3]−, and [HgCl4]2−) and hydroxides (such as [HgOH]+, Hg(OH)2, and [Hg(OH)3]−).48 The buffer solution used in our experimental system contained 0.10 mol L−1 chloride ion, and the pH value was 7.40. So, coordination equilibrium was used to calculate the contents of different Hg(II) species (S4). The results suggest that the main species of Hg(II) in our experimental system were HgCl2, [HgCl3]−, and [HgCl4]2−, at 41.35%, 29.27%, and 29.27%, respectively.
3.9 Molecular docking
It is apparent that different species of Hg(II) may have different effects on CAT. However, it is difficult to determine the difference through the experimental method. Hence, computational chemistry could be employed to resolve this question.49,50 Molecular docking was used to understand the interaction of CAT with different species of Hg(II) (HgCl2, [HgCl3]− and [HgCl4]2−).51 For each species of Hg(II), the lowest binding energy conformer was determined from 10 different conformers for further investigation.52 Fig. 9 shows the most possible interaction modes between CAT and different species of Hg(II), and the related data are shown in Tables 6 and 7. Fig. 9(A) shows that the binding sites of HgCl2, [HgCl3]− and [HgCl4]2− with CAT are significantly different. For HgCl2, the probe molecule is surrounded by amino acid residues Val 322, Glu 327, Pro 373, Val 374, Met 394, and Asp 395. The probe molecule of [HgCl3]− is located adjacent to amino acid residues Trp 14, Arg 18, Gln 21, Asp 24, and Arg 381. In the case of [HgCl4]2−, the amino acid residues consist of Pro 107, Arg 319, and Tyr 378. As shown in Table 6, electrostatic forces play a more important role in the binding interactions of CAT with HgCl2 and [HgCl3]−. In contrast, the dominant binding forces for [HgCl4]2− are van der Waals forces. In addition, no hydrogen bonds are observed between CAT and different species of Hg(II). Overall, electrostatic and van der Waals forces are the dominant intermolecular forces in the binding interactions of CAT with Hg(II). The binding energy values calculated from the docking studies for HgCl2–CAT, [HgCl3]−–CAT and [HgCl4]2−–CAT systems were −6.15, −3.06 and −1.92 kJ mol−1, with binding constants of 11.97, 3.42 and 2.17 L mol−1, respectively. Meanwhile, the experimentally calculated binding energy and binding constant were −6.23 kJ mol−1 and 13.24 L mol−1 for the Hg(II)–CAT system. The experimental values were close to the docking values of the HgCl2–CAT system, which may be because the content of HgCl2 was the highest in the experimental system.
 |
| | Fig. 9 Docking results of the Hg(II) and CAT system. (A) Binding sites of HgCl2, [HgCl3]− and [HgCl4]2− to CAT. The interaction mode of (B) HgCl2–CAT; (C) [HgCl3]−–CAT; (D) [HgCl4]2−–CAT. | |
Table 6 Docking summary of CAT with HgCl2, [HgCl3]− and [HgCl4]2−
| |
Binding energy (kJ mol−1) |
van der Waals energy (kJ mol−1) |
Electrostatic energy (kJ mol−1) |
Inhibition constant (mmol L−1) |
Binding constant (L mol−1) |
| HgCl2 |
−6.15 |
0.30 |
−6.45 |
83.55 |
11.97 |
| [HgCl3]− |
−3.06 |
−0.13 |
−2.93 |
292.27 |
3.42 |
| [HgCl4]2− |
−1.92 |
−1.17 |
−0.75 |
461.71 |
2.17 |
Table 7 Distance between the amino acid residues and the Hg(II) species
| Hg(II) species |
Atom of the Hg(II) |
Residue |
Atom of the residuea |
Distance (Å) |
| The first one character of the atom name consists of the chemical symbol for the atom type. All the atom names beginning with “C” are carbon atoms; “N” indicates a nitrogen and “O” indicates oxygen. The next character is the remoteness indicator code, which is transliterated according to: “B” stands for (∼) “β”; “G”∼“γ”; “D”∼“δ”; “E”∼“ε”; “Z”∼“ζ”; “H”∼“η”. |
| HgCl2 |
Hg |
Asp 395 |
OD |
2.2 |
| Cl 1 |
Val 322 |
CG |
3.8 |
| Cl 2 |
Glu 327 |
CG |
3.3 |
| Cl 2 |
Val 374 |
CG |
3.7 |
| Cl 2 |
Pro 373 |
CB |
3.7 |
| Cl 2 |
Met 394 |
O |
3.9 |
| [HgCl3]− |
Hg |
Asp 24 |
OD |
2.3 |
| Cl 1 |
Arg 381 |
NE |
3.3 |
| Cl 2 |
Gln 21 |
OE |
4.0 |
| Cl 3 |
Arg 18 |
NH |
3.1 |
| Cl 3 |
Trp 14 |
CZ |
3.7 |
| [HgCl4]2− |
Cl 1 |
Arg 319 |
NH |
3.2 |
| Cl 2 |
Tyr 378 |
OH |
3.2 |
| Cl 2 |
Pro 107 |
CB |
3.6 |
In this study, firstly, fluorescence, UV-vis and CD spectra were used to obtain the binding parameters of the interaction of Hg(II) with CAT and to confirm the significant structural changes and activity inhibition of CAT induced by Hg(II). Secondly, we focused on the fact which was too easy to be ignored that the metal ions may interact with other ions or molecules and then exist as different species. In order to distinguish between the interactions of CAT with different species of Hg(II), molecular docking was further employed. So, this work has significant implications for the research about the interactions of proteins with metal ions.
4. Conclusions
The above results showed that by combining multiple spectroscopic techniques and molecular docking simulation methods, the interaction mechanism of Hg(II) with CAT can be revealed in depth. The results indicated that Hg(II) can interact with CAT to form a complex through electrostatic and van der Waals forces. Low and high concentrations of Hg(II) can induce different conformational changes in CAT. The CAT activity was inhibited after binding with Hg(II), and the relative activity values were linearly associated with the CAT fluorescence intensity. Molecular docking results revealed that different species of Hg(II) were located at different sites on CAT, and detailed binding information was also explored. In conclusion, this study successfully furthered the understanding of the toxicity mechanism of Hg(II) on CAT at the molecular level.
It is well known that organic mercury is more toxic and bioavailable than inorganic mercury and can be biomagnified through trophic transfer. Hence, to fully understand the toxicity mechanism of mercury on an antioxidant enzyme system, further research should be performed to investigate the interaction mechanism of organic mercury with CAT in vitro.
Acknowledgements
This work was financially supported by the National Natural Science Foundation of China (No. 21075102, 21177102, 21577110), and the Specialized Research Fund for the Doctoral Program of Higher Education (SRFDP) (20130121130005).
References
- Y. Teng, L. Y. Zou, M. Huang and W. S. Zong, J. Photochem. Photobiol., B, 2014, 141, 241–246 CrossRef CAS PubMed.
- Y. Q. Wang and H. M. Zhang, Mater. Res. Bull., 2014, 60, 51–56 CrossRef CAS PubMed.
- A. Gönenç, A. Hacışevki, S. Aslan, M. Torun and B. Şimşek, Eur. J. Intern. Med., 2012, 23, 350–354 CrossRef PubMed.
- C. Soto, Nat. Rev. Neurosci., 2003, 4, 49–60 CrossRef CAS PubMed.
- B. J. Yang, F. Hao, J. R. Li, D. L. Chen and R. T. Liu, J. Photochem. Photobiol., B, 2013, 128, 35–42 CrossRef CAS PubMed.
- P. F. Qin and R. T. Liu, J. Hazard. Mater., 2013, 252, 321–329 CrossRef PubMed.
- F. Hao, M. Y. Jing, X. C. Zhao and R. T. Liu, J. Photochem. Photobiol., B, 2015, 143, 100–106 CrossRef CAS PubMed.
- Y. Mao, M. M. Hong, A. F. Liu and D. M. Xu, J. Fluoresc., 2015, 25, 755–761 CrossRef CAS PubMed.
- C. M. Dai, C. W. Ji, H. X. Lan, Y. Z. Song, W. Yang and D. Zheng, Environ. Toxicol. Pharmacol., 2014, 37, 870–877 CrossRef CAS PubMed.
- L. Magos, S. Halbach and T. W. Clarkson, Biochem. Pharmacol., 1978, 27, 1373–1377 CrossRef CAS.
- N. J. Langford and R. E. Ferner, J. Hum. Hypertens., 1999, 13, 651–656 CAS.
- T. W. Clarkson and L. Magos, CRC Crit. Rev. Toxicol., 2006, 36, 609–662 CrossRef CAS PubMed.
- M. K. Sharma, A. Sharma, A. Kumar and M. Kumar, Food Chem. Toxicol., 2007, 45, 2412–2419 CrossRef CAS PubMed.
- M. Farina, F. A. A. Soares, G. Zeni, D. O. Souza and J. B. T. Rocha, Toxicol. Lett., 2004, 146, 227–235 CrossRef CAS PubMed.
- D. Durak, S. Kalender, F. G. Uzun, F. Demýr and Y. Kalender, Afr. J. Biotechnol., 2010, 9, 488–495 CAS.
- Q. Xu, Y. N. Lu, L. Y. Jing, L. J. Cai, X. F. Zhu, J. Xie and X. Y. Hu, Spectrochim. Acta, Part A, 2014, 123, 327–335 CrossRef CAS PubMed.
- W. L. DeLano, DeLano Scientific, San Carlos, CA, 2002, p. 452 Search PubMed.
- A. S. Sharma, S. Anandakumar and M. Ilanchelian, RSC Adv., 2014, 4, 36267–36281 RSC.
- X. F. Zhang, G. Yang, Y. Dong, Y. Q. Zhao, X. R. Sun, L. Chen and H. B. Chen, Spectrochim. Acta, Part A, 2015, 137, 1280–1285 CrossRef CAS PubMed.
- A. Das and G. S. Kumar, RSC Adv., 2014, 4, 33082–33090 RSC.
- S. Huang, H. N. Qiu, S. Y. Lu, F. W. Zhu and Q. Xiao, J. Hazard. Mater., 2015, 285, 18–26 CrossRef CAS PubMed.
- J. R. Lakowicz, Principles of fluorescence spectroscopy, Springer Science & Business Media, 2007 Search PubMed.
- W. R. Ware, J. Phys. Chem., 1962, 66, 455–458 CrossRef CAS.
- Z. X. Chi, R. T. Liu and H. Zhang, Sci. Total Environ., 2010, 408, 5399–5404 CrossRef CAS PubMed.
- J. E. N. Dolatabadi, V. Panahi-Azar, A. Barzegar, A. A. Jamali, F. Kheirdoosh, S. Kashanian and Y. Omidi, RSC Adv., 2014, 4, 64559–64564 RSC.
- M. Shahlaei, B. Rahimi, M. R. Ashrafi-Kooshk, K. Sadrjavadi and R. Khodarahmi, J. Lumin., 2015, 158, 91–98 CrossRef CAS PubMed.
- A. Belatik, S. Hotchandani, R. Carpentier and H. A. Tajmir-Riahi, PLoS One, 2012, 7, 1–9 Search PubMed.
- J. Zhang, S. L. Zhuang, C. L. Tong and W. P. Liu, J. Agric. Food Chem., 2013, 61, 7203–7211 CrossRef CAS PubMed.
- P. D. Ross and S. Subramanian, Biochemistry, 1981, 20, 3096–3102 CrossRef CAS.
- S. Tunc, O. Duman, İ. Soylu and B. K. Bozoğlan, J. Hazard. Mater., 2014, 273, 36–43 CrossRef CAS PubMed.
- Y. Shu, W. W. Xue, X. Y. Xu, Z. M. Jia, X. J. Yao, S. W. Liu and L. H. Liu, Food Chem., 2015, 173, 31–37 CrossRef CAS PubMed.
- O. F. Sarioglu, A. Ozdemir, K. Karaboduk and T. Tekinay, J. Trace Elem. Med. Biol., 2015, 29, 111–115 CAS.
- T. Samejima, M. Kamata and K. Shibata, J. Biochem., 1962, 51, 181–187 CAS.
- Y. Teng, R. T. Liu, C. Li, Q. Xia and P. J. Zhang, J. Hazard. Mater., 2011, 190, 574–581 CrossRef CAS PubMed.
- Z. X. Chi, R. T. Liu, B. J. Yang and H. Zhang, J. Hazard. Mater., 2010, 180, 741–747 CrossRef CAS PubMed.
- X. F. Zhu, Y. N. Lu, J. D. Wang and Q. Xu, Anal. Methods, 2013, 5, 6037–6044 RSC.
- X. R. Li, G. K. Wang, D. J. Chen and Y. Lu, RSC Adv., 2014, 4, 7301–7312 RSC.
- B. Zhang, W. X. Zhai, R. T. Liu, Z. H. Yu, H. M. Shen and X. X. Hu, J. Nanosci. Nanotechnol., 2015, 15, 1473–1479 CrossRef CAS PubMed.
- C. Jash and G. S. Kumar, RSC Adv., 2014, 4, 12514–12525 RSC.
- J. Zhang, W. X. Chen, W. Zhang, Y. Duan, Y. X. Zhu, Y. X. Zhu and Y. Zhang, Chem. J. Chin. Univ., 2015, 36, 1511–1516 CAS.
- S. Huang, F. W. Zhu, Q. Xiao, Q. Zhou, W. Su, H. N. Qiu, B. Q. Hu, J. R. Sheng and C. S. Huang, RSC Adv., 2014, 4, 36286–36300 RSC.
- X. H. Zhang, R. Q. Gao, D. P. Li, H. Y. Yin, J. L. Zhang, H. Y. Cao and X. F. Zheng, Spectrochim. Acta, Part A, 2015, 136, 1775–1781 CrossRef CAS PubMed.
- X. L. Wei and Z. Q. Ge, Carbon, 2013, 60, 401–409 CrossRef CAS PubMed.
- A. Saha and V. V. Yakovlev, J. Biophotonics, 2010, 3, 670–677 CrossRef CAS PubMed.
- X. R. Li, D. J. Chen, G. K. Wang and Y. Lu, J. Lumin., 2014, 156, 255–261 CrossRef CAS PubMed.
- Y. D. Hu and L. J. Da, Spectrochim. Acta, Part A, 2014, 121, 230–237 CrossRef CAS PubMed.
- R. Lavery and S. Sacquin-Mora, J. Biosci., 2007, 32, 891–898 CrossRef CAS.
- T. R. Griffiths and R. A. Anderson, Can. J. Chem., 1991, 69, 451–457 CrossRef CAS.
- F. Zhang, J. Zhang, C. L. Tong, Y. D. Chen, S. L. Zhuang and W. P. Liu, J. Hazard. Mater., 2013, 263, 618–626 CrossRef CAS PubMed.
- M. S. Elgawish, N. Kishikawa, M. A. Helal, K. Ohyama and N. Kuroda, Toxicol. Res., 2015, 4, 843–847 RSC.
- R. Thakur, A. Das, V. Sharma, C. Adhikari, K. S. Ghosh and A. Chakraborty, Phys. Chem. Chem. Phys., 2015, 7, 16937–16946 RSC.
- D. Wu, J. Yan, P. X. Tang, S. S. Li, K. L. Xu and H. Li, Food Chem., 2015, 188, 370–376 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c5ra15301h |
|
| This journal is © The Royal Society of Chemistry 2015 |
Click here to see how this site uses Cookies. View our privacy policy here.