
Carob pods (Ceratonia siliqua L.) inhibit human neutrophils myeloperoxidase and in vitro ROS-scavenging activity
Abstract
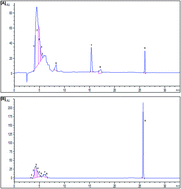
Natural antioxidants such as phenolic compounds protect cells against the damaging effects of reactive oxygen species (ROS). In this study, we investigated the effect of carob pods aqueous extract (CPAE) on the reactive oxygen species (ROS) production by human neutrophils, myeloperoxidase (MPO) activity and expression as well as lactoferrin and NADPH oxidase phosphorylation. Neutrophils were isolated from whole human blood using the ficoll–dextran method and ROS generation was measured by luminol-amplified chemiluminescence. Superoxide anion generation was detected by chemiluminescence using the lucigenin method. H2O2 was detected by the chemiluminescence assay. MPO activity was measured by the tetramethylbenzidine oxidation method. Western blotting analysis was used to determine the MPO and lactoferrin as well as P47phox–Ser-328 phosphorylation. The use of HPLC technique revealed the identification of many phenolic compounds in carob pods with pyrogallol as the main compound in the pulp and tannic acid in the seeds. We also found that CPAE inhibits luminol-amplified chemiluminescence in human neutrophils stimulated by PMA and that it is able to scavenge superoxide anion and hydrogen peroxide. Carob extract significantly reduces MPO activity and expression. More importantly, CPAE inhibits PMA-induced p47phox phosphorylation on Ser328 as well as lactoferrin release by neutrophils in a concentration-dependent manner. The effects are generally more marked for the seeds compared to the pulp. In conclusion, we suggest in the present study that Carob pods (Ceratonia siliqua L.) inhibit human neutrophils myeloperoxidase and in vitro ROS production.
 Please wait while we load your content...
Please wait while we load your content...

