
Occurrence of a functionally stable photoharvesting single peptide allophycocyanin α-subunit (16.4 kDa) in the cyanobacterium Nostoc sp. R76DM†
Abstract
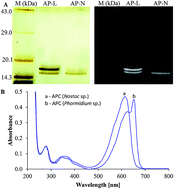
Allophycocyanin (APC) is a primary photoreceptor, usually composed of α- and β-polypeptide subunits. Herein, we report the occurrence of a functionally stable single peptide APC α-subunit in cyanobacterium Nostoc sp. R76DM. APC was purified successfully by ammonium sulfate fractionation. A series of biochemical characterizations like SDS-PAGE, native-PAGE, UV-visible spectroscopy, fluorescence spectroscopy and circular dichroism were performed to ensure the purity, integrity and functionality of the purified APC. Matrix-assisted laser-desorption ionization time-of-flight mass spectrometry (MALDI-TOF-MS) analysis of intact PBP revealed a ∼16.4 kDa protein. The MS/MS spectrum of five major peptides 1051, 1431, 936, 2291 and 2163 Da of trypsin digested purified PBP, followed by amino acid sequences of these peptides, shows a high degree (100%) of sequence similarities with that of the APC α-subunit (accession no. P16570, UniProtKB). The absorption as well as the fluorescence spectra of a single peptide APC α-subunit was shifted from normal absorption at 652 nm to 613 nm, and fluorescence at 663 nm to 645 nm. Urea-induced denaturation based Gibbs-free energy (ΔG°D) calculations suggested that the folding and structural stability of the APC α-subunit is almost similar to that of the standard APC (αβ) heterodimer from Lyngbya sp. Moreover, due to its conserved structural and functional integrity, the APC α-subunit may be widely used as a relatively low molecular weight fluorescent tag for fluorescence detection techniques.
 Please wait while we load your content...
Please wait while we load your content...

