
Host–guest interactions in an inverted cucurbit[7]uril with α,ω-alkyldiammonium guests†
Qing Li,
Sheng-Chao Qiu,
Ji-Hong Lu,
Sai-Feng Xue,
Xin Xiao*,
Zhu Tao and
Qian-Jiang Zhu*
Key Laboratory of Macrocyclic and Supramolecular Chemistry of Guizhou Province, Guizhou University, Guiyang 550025, P. R. China. E-mail: gyhxxiaoxin@163.com; gzuzhuqj@163.net
First published on 4th August 2015
Abstract
Host–guest interactions in an aqueous solution of an inverted cucurbit[7]uril (iQ[7]) with α,ω-alkyldiammonium guests (H3N(CH2)xNH3Cl2, x = 2, 4, 6, 8, 10, and 12 named 1–6) were investigated by nuclear magnetic resonance (NMR) and isothermal titration calorimetry (ITC). 1H NMR spectroscopy revealed that all alkyldiammonium cations are involved in interactions with iQ[7] that fall into three groups. Averaged signals of free and bound guests with shorter alkyl chains including 1–3 indicate a higher binding and unbinding exchange ratio, within the time scale of the NMR experiments. Averaged signals for the guest 1 compound exhibited a downfield shift indicative of a dominant portal interaction between 1 and iQ[7], in contrast to guest 2 and 3 compounds that exhibited an upfield shift. Averaged signals of guests with longer alkyl chains such as 4–6 displayed a lower binding and unbinding exchange ratio on the NMR time scale. The occurrence of upfield shifts for all guests except 1 indicate a dominant inclusion interaction between the guests and iQ[7]. ITC experiments showed that the thermodynamics of complexation of the alkyldiammonium guests with iQ[7] were both enthalpically and entropically driven. Interestingly, complexation was predominantly entropically driven for the first guest group, with the exception of guest 3, but was enthalpically driven for the third guest group. Host–guest interactions of the more conventional cucurbit[7]uril (Q[7]) with these α,ω-alkyldiammonium guests were also investigated under the same conditions for comparison.
Ten years ago, Isaacs and Kim first discovered two inverted cucurbit[n]urils (iQ[n]s, n = 6 and 7) that exhibited the characteristic movement of the two methine hydrogen atoms of a glycoluril unit into the cavity of the iQ[n]s, and patched host–guest interactions were also reported in 2005.1 In 2007 they demonstrated that iQ[n]s can convert into Q[n]s following heating in concentrated HCl.2 It is conceivable that iQ[7] has a slightly smaller cavity than that of Q[7], and displays some different host–guest interaction properties. However, the difficulties associated with separating iQ[n]s appears to have hindered investigation of iQ[n] chemistry since no further reports in this area have been made. To date, only four studies and one patent involving iQ[n]s have been reported.2–5 Recently, our group found that iQ[6] or iQ[7] can be easily isolated from mixtures of Q[n]s by column chromatography using a Dowex resin6,7 due to subtle differences in the surface electrostatic properties of Q[n] and iQ[n] that result in significant differences in chromatographic behavior.8 Both classes of compound have been shown to form close interactions with alkaline earth metal ions6,7 and lanthanide cations9,10 in the presence of [ZnCl4]2− that acts as a structure-directing agent10–12 to form a variety of supramolecular assemblies. However, our knowledge of the host–guest chemistry of iQ[n]s remains far from complete, although Isaacs and Kim have claimed that iQ[n]s can bridge and exceed the recognition properties of known Q[n]s by displaying unprecedented size and shape selectivity.
In order to investigate the host–guest chemistry of iQ[n]s, in the present work iQ[7] was used as a host (Fig. 1) and a series of α,ω-alkyldiammonium guests (H3N(CH2)xNH3, x = 2, 4, 6, 8, 10, and 12 named 1–6) were tested as guests. Host–guest interactions were investigated in aqueous solution by 1H NMR spectroscopy, which revealed that all alkyldiammonium cations formed interactions with iQ[7], and these fell into three groups. Averaged signals of guests with shorter alkyl chains such as 1–3 displayed a faster binding and unbinding exchange ratio on the NMR time scale, whereas averaged signals of guests with longer alkyl chains such as 4–6 displayed a lower binding and unbinding exchange ratio on the NMR time scale. Averaged signals for guest 1 compounds exhibited a downfield shift indicative of a dominant portal interaction between 1 and iQ[7], in contrast to guest 2–6 compounds that exhibited an upfield shift. The occurrence of upfield shifts for all guests except 1 indicate a dominant inclusion interaction between guests and iQ[7]. ITC experiments showed that the thermodynamics of complexation of the alkyldiammonium guests with iQ[7] were both enthalpically and entropically driven and complexation were predominantly entropically or enthalpically driven dependently on the substituted alkyl chains. Host–guest interactions of the more conventional cucurbit[7]uril (Q[7]) with these α,ω-alkyldiammonium guests were also investigated under the same condition for comparison.
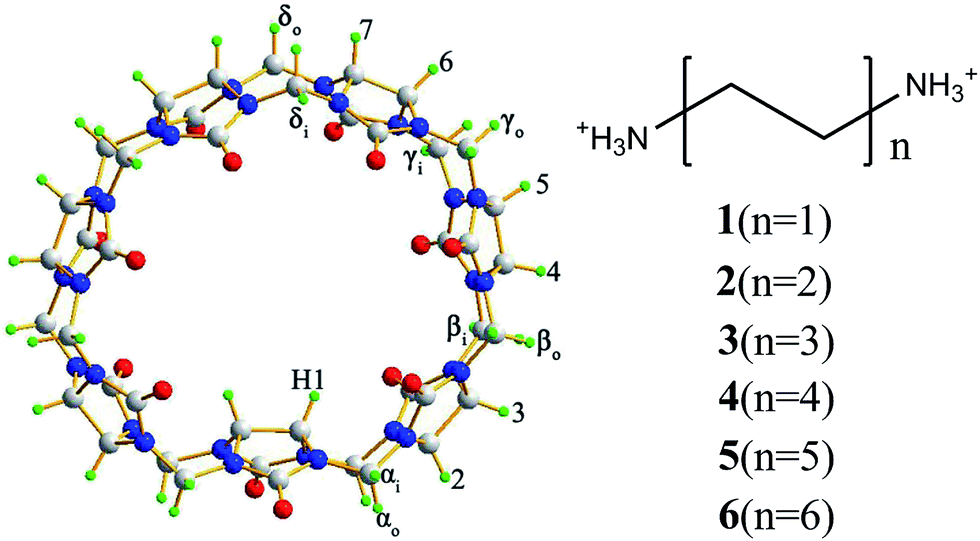 | ||
| Fig. 1 Structures of the inverted cucurbit[7]uril (iQ[7]) and α,ω-alkyldiammonium guests +H3N(CH2)xNH3+, (x = 2, 4, 6, 8, 10, and 12 named 1–6). | ||
We first investigated the interaction of guest 1 with the iQ[7] host in neutral aqueous solution. Fig. 2 (left) shows the 1H NMR spectra obtained when a fixed molar equivalent of iQ[7] was titrated with various equivalents of 1. One proton signal (Ha) from guest 1 was shifted slightly downfield with increasing amounts of 1 (Fig. 2b to e, left), suggesting a portal interaction between 1 with iQ[7] dominates. In contrast, the two methine protons (H1 on Fig. 1) of the inverted glycoluril unit in the cavity of iQ[7] were shifted upfield with increasing amounts of 1, suggesting that 1 also interacts with these methine protons (H1) in the cavity of iQ[7], but this interaction is not dominant. Thus, the averaged signals of free, portal and cavity-bound guest molecules were shifted slightly downfield. However, 1H NMR spectra obtained from a fixed equivalent of Q[7] titrated with various equivalents of 1 showed that the averaged signals of free, portal and cavity-bound guest molecules were shifted slightly upfield, suggesting that the cavity interaction of Q[7] with 1 was dominant (Fig. 2, right). In a previous study we demonstrated that the interaction of 1 with a Q[6] (TMeQ[6])13 was similar to that of iQ[7] with 1. Thus, the size of iQ[7] is estimated to be smaller than Q[7], and similar to Q[6].
 | ||
| Fig. 2 Titration 1H NMR spectra (400 MHz, D2O) for (top, a) neat iQ[7], (b–e) iQ[7] obtained in the presence of various equivalents of 1 and (f) neat 1; (below, a) neat Q[7], (b–d) Q[7] obtained in the presence of various equivalents of 1 or (e) neat 1. | ||
1H NMR spectra were obtained with a fixed equivalent of iQ[7] and various equivalents of 2 (Fig. 3, left). Only one set of proton signals of guest 2 displayed a gradual downfield shift with an increasing amount of 2 (Fig. 3b–e, left), suggesting that the guest is predominantly located in a shielded environment. These NMR signals are averages of free and bound guest molecules, which have a high binding and unbinding exchange ratio on the NMR time scale. The inverted methine proton signals of the iQ[7] host displayed a gradual upfield shift with increasing amounts of 2 compared with 1H NMR spectra of neat hosts (Fig. 3a). This indicates the existence of a cavity interaction between iQ[7] and 2. No other significant changes for other protons of the iQ[7] host in the presence of 2 were apparent, presumably because they are too far away from guest 2, unlike the inverted methine protons. Interactions between Q[6] and Q[7] with the same series of guests used in the present work were recently reported,14 and no interaction between Q[7] an guest 2 was apparent in 50% formic acid in ITC experiments or in formic acid/D2O (30/70 v/v) in NMR measurements. Competition between guest molecules and protonated water or formic acid molecules may greatly affect Q[7]–guest 2 interactions, whereas in neutral water, the iQ[7] host has a smaller cavity and Q[7] can more easily include guest 2. 1H NMR spectra obtained using a fixed equivalent of Q[7] and titrating various equivalents of 2 (Fig. 3, right) revealed a significant upfield shift with decreasing amounts of 2. This suggests that Q[7] has a stronger affinity for guest 2 than does iQ[7] and indicates the steric hindrance of the inverted methine group in the cavity of the iQ[7] molecule.
 | ||
| Fig. 3 Titration 1H NMR spectra (400 MHz, D2O) for (top, a) neat iQ[7], (b–e) iQ[7] obtained in the presence of various equivalents of 2 and (f) neat 2; (below, a) neat Q[7], (b–d) Q[7] obtained in the presence of various equivalents of 2 or (e) neat 2. | ||
The remarkable differences between the 1H NMR titration spectra of the iQ[7]–3 and Q[7]–3 interaction systems reflect the binding and unbinding exchange ratio of guest 3 on the NMR time scale. For iQ[7]–3, the exchange ratio was faster than the NMR time scale (Fig. 4, left), but this was clearly slower than the NMR time scale for Q[7]–3 (Fig. 4, right). A closer inspection revealed that although the exchange ratios of the two interaction systems differed, no significant difference in the shielding effect of cavities was apparent. For example, the resonance of proton Hc of guest 3 experienced a ∼0.8 ppm shift in Q[7]–3 but a ∼0.7 ppm shift in the iQ[7]–3 interaction system. Comparison of free iQ[7] vs. bound iQ[7] indicated that only βi, γi, δi (Fig. 1) experienced a slight upfield shift, while the rest of the proton resonances of the iQ[7] host showed no obvious changes upon an increasing number of equivalents of 3. In contrast, in the Q[7]–3 interaction system, the methylene proton resonances (at 4.2 ppm) and the waist proton resonances at (5.4 ppm) of the free Q[7] experienced a downfield shift of 0.1 ppm (Fig. 4, right).
 | ||
| Fig. 4 Titration 1H NMR spectra (400 MHz, D2O) for (top, a) neat iQ[7], (b–e) iQ[7] obtained in the presence of various equivalents of 3 or (f) neat 3; (below, a) neat Q[7], (b–d) Q[7] obtained in the presence of various equivalents of 3 or (e) neat 3. | ||
For the guests with longer alkyl chains, such as 4–6 show a slower exchange ratio on the NMR time scale, two sets of proton resonances represented the binding and unbinding guests can be observed in the titration 1H NMR spectra for the iQ[7]–guest (4–6) respectively. For example, Fig. 5 shows 1H NMR spectra obtained by using a fixed equivalent of iQ[7] and various equivalents of 4. One can see four proton resonances of the binding 4 when the host![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :guest mole ratio is 1:0.74 (Fig. 5b), and both of four proton resonances of the binding 4 and three proton resonances of the unbinding 4 when the host:guest mole ratio is 1:2.82 (Fig. 5c), the proton resonances of the binding 4 experience upfield shifts by 0.156 ppm (Ha), 0.52 ppm (Hb), 0.576 ppm (Hc) and 0.638 ppm (Hd), respectively.
:guest mole ratio is 1:0.74 (Fig. 5b), and both of four proton resonances of the binding 4 and three proton resonances of the unbinding 4 when the host:guest mole ratio is 1:2.82 (Fig. 5c), the proton resonances of the binding 4 experience upfield shifts by 0.156 ppm (Ha), 0.52 ppm (Hb), 0.576 ppm (Hc) and 0.638 ppm (Hd), respectively.
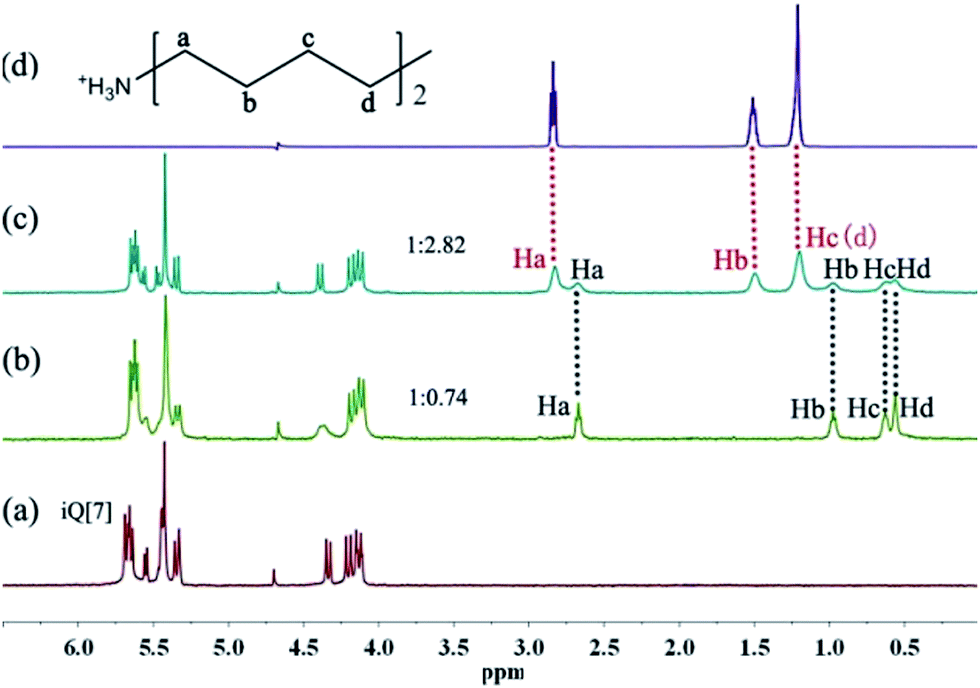 | ||
| Fig. 5 Titration 1H NMR spectra (400 MHz, D2O) (a) neat iQ[7], (b–c) iQ[7] obtained in the presence of various equivalents of 4 or (d) neat 4. | ||
The titration 1H NMR spectra for the iQ[7]–5 and iQ[7]–6 interaction systems present similar results (referring to Fig. SI-1I and K in the ESI†), and the comparison of the Q[7]–4, Q[7]–5 and Q[7]–6 interaction systems also show the similar titration 1H NMR spectra (referring to Fig. SI-1H, J and L in the ESI†). One can see both proton resonances of the binding and unbinding guests. A close inspection reveals that the shielding effect of cavity of Q[7] slightly stronger than that of iQ[7] for the guest, because proton resonances of the guest in the Q[7]–guest interaction systems generally present an extra upfield shift by 0.1–0.2 ppm than those in the iQ[7]–guest interaction systems (referring to the data in Fig. SI-3 in the ESI,† round brackets for iQ[7] systems and square brackets for Q[7] systems).
It should be noted that proton resonances of the inverted methine (H1) present interesting behaviors: in the iQ[7]–1 and iQ[7]–2 interaction systems, H1 experiences upfield shift with increasing number of equivalents of 1 or 2; in the iQ[7]–3 and iQ[7]–4 interaction systems, H1 does not show in the spectra; in the iQ[7]–5 and iQ[7]–6 interaction systems, H1 does not show in the spectra until in the presence of extra guest. When a guest, such as 2 to 6, is included in the cavity of an iQ[7] molecule, it must interact with the inverted methine protons (H1), resulting in the chemical shift change of them. The H1 could overlap in the HDO resonance, which is delimited in the magnetic saturation experiments; another reason could be broaden when they interact with guest. The further detailed explanation should get more information from further iQ[7]–guest interaction investigations.
Above, we described the interaction between iQ[7] and a series of α,ω-alkyldiammonium guests in neutral water solutions. Isothermal titration calorimetry (ITC) is one of limited suitable quantitative technique for detecting the interaction between iQ[7] and α,ω-alkyldiammonium guests in liquid states, because the iQ[7] host and the α,ω-alkyldiammonium guests are lack of proper spectroscopic properties. Table 1 and Fig. SI-2† show the ITC results for both of iQ[7]–α,ω-alkyldiammonium and Q[7]–α,ω-alkyldiammonium interaction systems. The experimental results reveal that the Ka of iQ[7]–α,ω-alkyldiammonium interaction systems are gradually increasing with the increase of the length of the alkyl chains of the α,ω-alkyldiammonium guests, whereas the Ka of Q[7]–α,ω-alkyldiammonium interaction systems are gradually increasing and then decreased with the increase of the length of the alkyl chains of the α,ω-alkyldiammonium guests. Generally, besides the cavity interaction of the Q[n] host with the α,ω-alkyldiammonium guest, the interaction of the protonated amino moieties of a α,ω-alkyldiammonium guest with the portal carbonyl oxygens of a Q[n] host could present the extra contribution to the interaction of the Q[n] host with the α,ω-alkyldiammonium guest. When the alkyl chain of the α,ω-alkyldiammonium guest is longer enough, the two amino moieties of a α,ω-alkyldiammonium guest can interact with both portals of the Q[7] or iQ[7] molecule, combination of the cavity interaction, the formed host–guest inclusion complex could be the most stable and present the highest Ka value. Whereas the alkyl chain of the α,ω-alkyldiammonium guest is too long to effectively interact with both portals of the Q[7] molecule, the formed host–guest inclusion complex could show less stable as shown in Q[7]–α,ω-alkyldiammonium interaction systems in Table 1. For the iQ[7]–α,ω-alkyldiammonium interaction systems, an extra interaction between the alkyl chain of the α,ω-alkyldiammonium guest with the inverted methine could lead to the increase trend of the Ka. On the other hand, the existence of the inverted methine in the cavity seem to frustrate the formation of helical conformations and interaction of the α,ω-alkyldiammonium guest with cavity of iQ[7] molecule, resulting in the decrease of Ka for the corresponding systems in comparison with that of Q[7]–α,ω-alkyldiammonium interaction systems (referring to the data in Table 1).
| Experiment | Model | Ka (1/M) | ΔH (kJ mol−1) | TΔS(kJ mol−1) |
|---|---|---|---|---|
| iQ[7]–1 | Independent | (6.002 ± 0.27) × 104 | −1.087 ± 0.14 | 26.19 |
| iQ[7]–2 | (6.286 ± 0.90) × 104 | −2.235 ± 0.19 | 25.16 | |
| iQ[7]–3 | (4.18 ± 0.84) × 105 | −16.63 ± 0.49 | 15.45 | |
| iQ[7]–4 | (1.79 ± 0.54) × 106 | −24.69 ± 0.65 | 11.01 | |
| iQ[7]–5 | (2.15 ± 0.64) × 106 | −25.13 ± 0.49 | 11.02 | |
| iQ[7]–6 | (2.47 ± 0.42) × 106 | −27.70 ± 0.48 | 8.78 | |
| Q[7]–1 | (1.56 ± 0.31) × 105 | −17.36 ± 0.75 | 12.28 | |
| Q[7]–2 | (1.74 ± 0.45) × 105 | −19.02 ± 0.39 | 16.60 | |
| Q[7]–3 | (4.596 ± 0.8) × 106 | −31.37 ± 0.29 | 6.65 | |
| Q[7]–4 | (7.706 ± 0.73) × 106 | −40.929 ± 0.25 | 1.62 | |
| Q[7]–5 | (4.356 ± 0.59) × 107 | −39.521 ± 0.79 | 4.08 | |
| Q[7]–6 | (1.33 ± 0.44) × 107 | −31.87 ± 0.26 | 8.79 |
According to the data in Table 1, both iQ[7]–α,ω-alkyldiammonium interaction systems and Q[7]–α,ω-alkyldiammonium interaction systems experience exothermal reactions, for the former, enthalpies gradually increase with the increase of the length of the alkyl chains of the α,ω-alkyldiammonium guests, whereas for the later, enthalpies gradually increase and then decrease with the increase of the length of the alkyl chains of the α,ω-alkyldiammonium guests (Table 1). The positive entropies for both host–guest interaction systems could be attributed to interactions between Q[7]s (iQ[7] or Q[7]) and solvent water molecules, including cavity inclusion and portal interaction through hydrogen bonding, also the surfaces of the extended alkyl chain of the guests are covered with solvent water molecules, and the release of solvent from these surfaces and cavity is favorable to the entropic and enthalpic changes as the inclusion complex forms. For the iQ[7]–guest systems, entropies gradually decrease with the increase of the length of the alkyl chains of the α,ω-alkyldiammonium guests, whereas for the Q[7]–guest system, entropies gradually decrease and then increase with the increase of the length of the alkyl chains of the α,ω-alkyldiammonium guests (Table 1). Thus, the host–guest interactions of both of the iQ[7]–guest and Q[7]–guest systems are basically driven by both enthalpy and entropy. However, a close inspection reveals that for the iQ[7]–1 and iQ[7]–2 systems, the intrinsic characteristics of the host–guest interaction are dominated by entropic-driven thermodynamics. Whereas for the iQ[7]–4 or 5 or 6 systems, the intrinsic characteristics of the host–guest interaction are dominated by enthalpic-driven thermodynamics. Whereas for the Q[7]–guest systems, the intrinsic characteristics of the host–guest interaction are dominated by enthalpic-driven thermodynamics.
Conclusions
To our knowledge, this is the first systematic investigation on the interactions of iQ[7] hosts with α,ω-alkyldiammonium guests (1–6) in aqueous solution. Our results revealed that interactions between iQ[7] and guests with shorter alkyl chains (1–3) exhibited a higher binding and unbinding exchange ratio that was visible on the time scale of the NMR experiments, whereas interactions with guests with longer alkyl chains (4–6) displayed lower binding and unbinding exchange ratios. The Q[7]–guest systems with both shorter (1–2) and longer (3–6) alkyl chains showed a higher exchange ratio than did the iQ[7] systems involving guests with longer alkyl chains (4–6). The interaction between iQ[7] and guest 1 was dominated by a portal interaction, whereas with the other α,ω-alkyldiammonium guests (2–6) an inclusion interaction model dominated in both Q[7] and iQ[7] systems. Guest proton resonances of Q[7]–guest systems were generally shifted upfield by 0.1–0.2 ppm compared with the corresponding iQ[7]–guest systems, indicating that Q[7]–guest interactions are more stable. Ka values from ITC measurements also supported this conclusion. Moreover, the ITC experiments revealed that host–guest interactions in both iQ[7]–1 and iQ[7]–2 systems were dominated by entropically-driven thermodynamics, whereas for iQ[7]–4, 5 and 6 systems, the thermodynamics of complexation was enthalpically driven.Notes and references
- L. Isaacs, S. K. Park, S. M. Liu, Y. H. Ko, N. Selvapalam, Y. Kim, H. Kim, P. Y. Zavalij, G. H. Kim, H. S. Lee and K. Kim, J. Am. Chem. Soc., 2005, 127, 18000 CrossRef CAS PubMed.
- S. M. Liu, K. Kim and L. Isaacs, J. Org. Chem., 2007, 72, 6840 CrossRef CAS PubMed.
- L. D. Isaacs, S. M. Liu, K. Kim, S. K. Park, Y. H. Ko, H. Kim, Y. Kim and N. Selvapalam, Introverted cucurbituril compounds and their preparation, crystal structure and binding properties., PCT Int. Appl., 2007, 48 Search PubMed.
- R. V. Pinjari and S. P. Gejji, J. Phys. Chem. A, 2009, 113, 1368 CrossRef CAS.
- I. A. Raja, V. V. Gobre, R. V. Pinjari and S. P. Gejji, J. Mol. Model., 2014, 20, 1 CrossRef CAS PubMed.
- D. Q. Zhang, T. Sun, Y. Q. Zhang, S. F. Xue, Q. J. Zhu, J. X. Zhang and Z. Tao, Eur. J. Inorg. Chem., 2015, 318, 318 CrossRef PubMed.
- Q. Li, Y. Q. Zhang, Q. J. Zhu, S. F. Xue, Z. Tao and X. Xiao, Chem.–Asian J., 2015, 10, 1159 CrossRef CAS PubMed.
- X. L. Ni, X. Xiao, H. Cong, Q. J. Zhu, S. F. Xue and Z. Tao, Acc. Chem. Res., 2014, 47, 1386 CrossRef CAS PubMed.
- L. L. Liang, X. L. Ni, Y. Zhao, K. Chen, X. Xiao, Y. Q. Zhang, C. Redshaw, Q. J. Zhu, S. F. Xue and Z. Tao, Inorg. Chem., 2013, 52, 1909 CrossRef CAS PubMed.
- Y. Zhao, L. L. Liang, K. Chen, T. Zhang, X. Xiao, Y. Q. Zhang, Z. Tao, S. F. Xue and Q. J. Zhu, CrystEngComm, 2013, 15, 7987 RSC.
- L. L. Liang, Y. Zhao, Y. Q. Zhang, Z. Tao, S. F. Xue, Q. J. Zhu and J. X. Liu, CrystEngComm, 2013, 15, 3943 RSC.
- L. L. Liang, X. L. Ni, Y. Zhao, K. Chen, X. Xiao, Y. Q. Zhang, C. Redshaw, Q. J. Zhu, S. F. Xue and Z. Tao, Inorg. Chem., 2013, 52, 1909 CrossRef CAS PubMed.
- B. Yang, L. M. Zheng, Z. Z. Gao, X. Xiao, Q. J. Zhu, S. F. Xue, Z. Tao, J. X. Liu and G. Wei, J. Org. Chem., 2014, 79, 11194 CrossRef CAS PubMed.
- S. J. C. Lee, J. W. Lee, H. H. Lee, J. Seo, D. H. Noh, Y. H. Ko, K. Kim and H. I. Kim, J. Phys. Chem. B, 2013, 117, 8855 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Titration 1H NMR spectra and ITC for iQ[7]–guest (1–6) and Q[7]–guest (1–6). See DOI: 10.1039/c5ra13339d |
| This journal is © The Royal Society of Chemistry 2015 |